фигуры 1 и 8
). При делении Я. (см.
Митоз
) весь хроматин конденсируется в хромосомы; по окончании митоза большая часть участков хромосом опять разрыхляется; эти участки (так называемый эухроматин) содержат в основном уникальные (неповторяющиеся) гены. Другие участки хромосом остаются плотными (так называемый гетерохроматин); в них располагаются главным образом повторяющиеся последовательности ДНК. В неделящемся Я. большая часть эухроматина представлена рыхлой сетью фибрилл ДНП толщиной 10 — 30
нм, гетерохроматин — плотными глыбками (хромоцентрами), в которых те же фибриллы плотно упакованы. Часть эухроматина также может переходить в компактное состояние; такой эухроматин считается неактивным в отношении синтеза РНК. Хромоцентры обычно граничат с ЯО или ядрышком. Есть данные о том, что фибриллы ДНП закреплены на внутренней мембране ЯО.
В неделящемся Я. происходит синтез ( репликация ) ДНК, изучаемый путём регистрации включенных в Я. меченных радиоактивными изотопами предшественников ДНК (обычно тимидина). Показано, что по длине хроматиновых фибрилл имеется множество участков (так называемых репликонов), каждый со своей точкой начала синтеза ДНК, от которой репликация распространяется в обе стороны. Вследствие репликации ДНК удваиваются и сами хромосомы.
В хроматине Я. происходит считывание закодированной в ДНК генетической информации путём синтеза на ДНК молекул матричной, или информационной, РНК (см. Транскрипция), а также молекул других типов РНК, участвующих в белковом синтезе. Специальные участки хромосом (и соответственно хроматина) содержат повторяющиеся гены, которые кодируют молекулы рибосомной РНК; в этих местах Я. формируются богатые рибонуклеопротеидами (РНП) ядрышки, основная функция которых — синтез РНК, входящей в состав рибосом. Наряду с компонентами ядрышка в Я. есть и другие виды частиц РНК. К ним относятся перихроматиновые фибриллы толщиной 3—5 нми перихроматиновые гранулы (ПГ) диаметром 40—50 нм, расположенные на границах зон рыхлого и компактного хроматина. И те и другие, вероятно, содержат матричную РНК в соединении с белками, а ПГ отвечают её неактивной форме; наблюдался выход ПГ из Я. в цитоплазму через поры ЯО. Имеются также интерхроматиновые гранулы (20—25 нм), а иногда и толстые (40—60 нм) нити РНП, скрученные в клубки. В ядрах амёб имеются нити РНП, скрученные в спирали (30—35 нмх 300 нм); спирали могут выходить в цитоплазму и, вероятно, содержат матричную РНК. Наряду с ДНК- и РНК-содержащими структурами некоторые Я. содержат чисто белковые включения в виде сфер (например, в Я. растущих яйцеклеток многих животных, в Я. ряда простейших), пучков фибрилл или кристаллоидов (например, в ядрах многих тканевых клеток животных и растений, макронуклеусах ряда инфузорий). В Я. обнаружены также фосфолипиды, липопротепды, ферменты (ДНК-полимераза, РНК-полимераза, комплекс ферментов оболочки Я., в том числе аденозинтрифосфатаза, и др.).
В природе встречаются различные специальные типы Я.: гигантские Я. растущих. яйцеклеток, особенно рыб и земноводных; Я., содержащие гигантские политенные хромосомы (см. Политения ), например в клетках слюнных желёз двукрылых насекомых; компактные, лишённые ядрышек Я. сперматозоидов и микронуклеусы инфузорий, сплошь заполненные хроматином и не синтезирующие РНК; Я., в которых хромосомы постоянно конденсированы, хотя ядрышки образуются (у некоторых простейших, в ряде клеток насекомых); Я., в которых произошло дву- или многократное увеличение числа наборов хромосом ( полиплоидия ; фигуры 7, 9 ).
Основной способ деления Я. — митоз, характеризующийся удвоением и конденсацией хромосом, разрушением ЯО (исключение — многие простейшие и грибы) и правильным расхождением сестринских хромосом в дочерние клетки. Однако Я. некоторых специализированных клеток, особенно полиплоидные, могут делиться простой перешнуровкой (см. Амитоз ) .Высокополиплоидные Я. могут делиться не только на 2, но и на много частей, а также почковаться ( фигура 7 ). При этом может происходить разделение целых хромосомных наборов (т. н. сегрегация геномов).
Лит.:Руководство по цитологии, т. 1, М. —Л., 1965; Райков И. Б., Кариология простейших, Л., 1967; Робертис Э., Новинский В., Саэс Ф.,. Биология клетки, пер. с англ., М., 1973; Ченцов Ю. С., Поляков В. Ю., Ультраструктура клеточного ядра, М., 1974; The nucleus, ed. A. J. Dalton, F, Haguenau, N. Y. — L., 1968; The cell nucleus, ed. Н. Busch, v. 1—3, N. Y. — L., 1974.
И. Б. Райков.
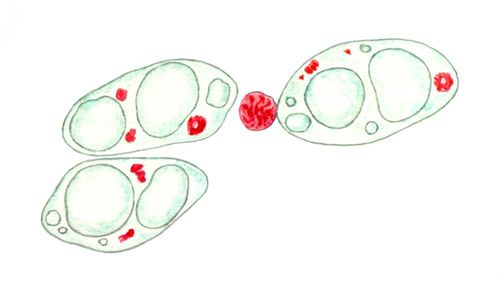
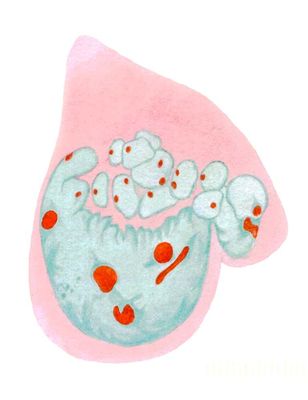 Клеточные ядра на окрашенных препаратах: фрагментация гигантского полиплоидного ядра трофобласта крысы, окраска метиловым зеленым — пиронином (хроматин зеленый, ядрышки красные, цитоплазма розовая).
Клеточные ядра на окрашенных препаратах: фрагментация гигантского полиплоидного ядра трофобласта крысы, окраска метиловым зеленым — пиронином (хроматин зеленый, ядрышки красные, цитоплазма розовая).
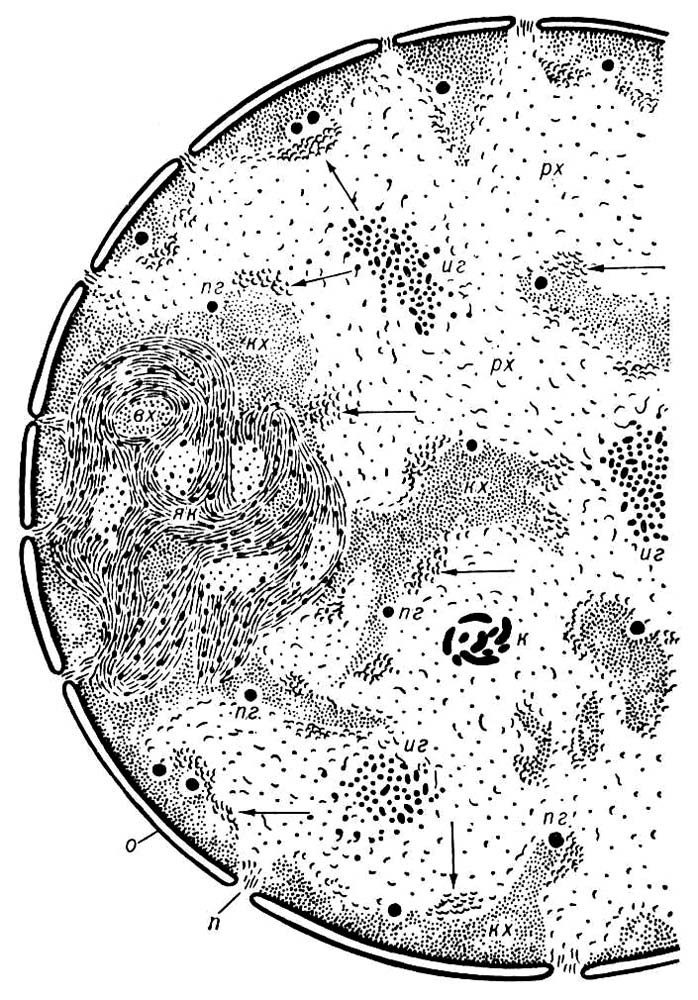 Схема ультраструктуры ядра клетки печени: зоны компактного (кх) и рыхлого (рх) хроматина; ядрышко (як) с внутри-ядрышковым хроматином (вх), перихро-матиновые фибриллы (стрелки), перихроматнновые (пг) и интерхроматиновые (иг) гранулы; рибонуклеопротеидная нить, свёрнутая в клубок (к); оболочка ядра (яо) с порами (п).
Схема ультраструктуры ядра клетки печени: зоны компактного (кх) и рыхлого (рх) хроматина; ядрышко (як) с внутри-ядрышковым хроматином (вх), перихро-матиновые фибриллы (стрелки), перихроматнновые (пг) и интерхроматиновые (иг) гранулы; рибонуклеопротеидная нить, свёрнутая в клубок (к); оболочка ядра (яо) с порами (п).
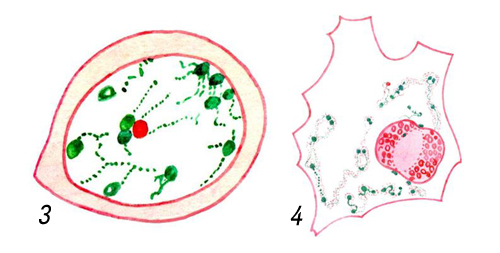 Клеточные ядра на окрашенных препаратах: яйцеклетки креветки рода Palaemon на ранней (3) и поздней (4, изображено только ядро) стадиях, в процессе развития ядро приобретает неправильную форму, хроматин (зеленый) рассеивается, ядрышко (красное) растет и вакуолизируется, окраска метиловым зеленым — пиронином.
Клеточные ядра на окрашенных препаратах: яйцеклетки креветки рода Palaemon на ранней (3) и поздней (4, изображено только ядро) стадиях, в процессе развития ядро приобретает неправильную форму, хроматин (зеленый) рассеивается, ядрышко (красное) растет и вакуолизируется, окраска метиловым зеленым — пиронином.
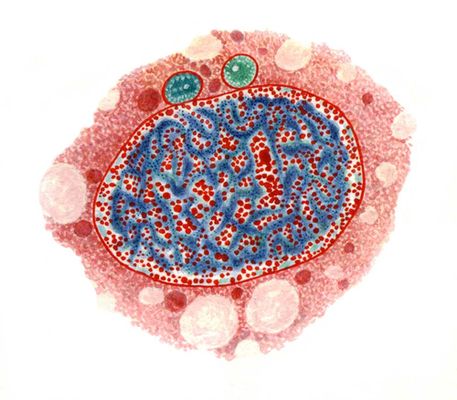 Клеточные ядра на окрашенных препаратах: полиплоидный макронуклеус и два микронуклеуса инфузории рода Nassula, окраска метиловым зеленым — пиронином: хроматин (фиолетовый) содержит и ДНК, и РНК, ядрышки (красные) — РНК, микронуклеусы зеленые.
Клеточные ядра на окрашенных препаратах: полиплоидный макронуклеус и два микронуклеуса инфузории рода Nassula, окраска метиловым зеленым — пиронином: хроматин (фиолетовый) содержит и ДНК, и РНК, ядрышки (красные) — РНК, микронуклеусы зеленые.
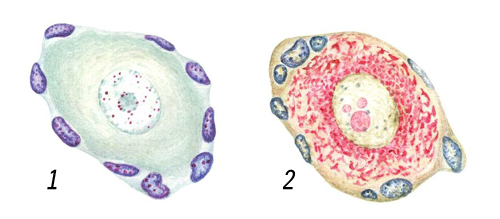 Клеточные ядра на окрашенных препаратах: нервные клетки с крупным ядром, окраска (красно-фиолетовая) по Фёльгену на ДНК (1), метиловым зеленым на ДНК и пиронином (красный цвет) на РНК (2); по краям видны ядра соединительнотканных клеток.
Клеточные ядра на окрашенных препаратах: нервные клетки с крупным ядром, окраска (красно-фиолетовая) по Фёльгену на ДНК (1), метиловым зеленым на ДНК и пиронином (красный цвет) на РНК (2); по краям видны ядра соединительнотканных клеток.
 Клеточные ядра на окрашенных препаратах: три диплоидных макронуклеуса и один микронуклеус инфузории рода Remanella, окраска по Фёльгену на ДНК, в макронуклеусах видны хромоцентры (красно-фиолетовые) и ядрышки (бледно-зеленые).
Клеточные ядра на окрашенных препаратах: три диплоидных макронуклеуса и один микронуклеус инфузории рода Remanella, окраска по Фёльгену на ДНК, в макронуклеусах видны хромоцентры (красно-фиолетовые) и ядрышки (бледно-зеленые).
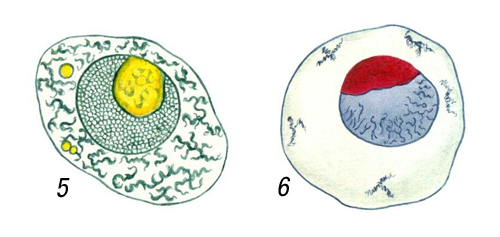 Клеточные ядра на окрашенных препаратах: ядра яйцеклеток моллюсков рода Littorina, видны две различно окрашивающиеся части ядрышка (амфинуклеолы), окраска метиловым синим — оранженвым Ж (5) и азаном (6).
Клеточные ядра на окрашенных препаратах: ядра яйцеклеток моллюсков рода Littorina, видны две различно окрашивающиеся части ядрышка (амфинуклеолы), окраска метиловым синим — оранженвым Ж (5) и азаном (6).
заболони
(коричневая — у дуба, жёлтая — у барбариса, акации, красноватая — у тисса, оранжевая — у ольхи, почти чёрная — у эбенового дерева). Вследствие закупорки сосудов и трахеид тиллами, а также смолами, камедями, эфирными маслами и пр. Я. д. почти не проницаемо для воды и воздуха, устойчиво к гниению и поражению грибами.
В неделящемся Я. происходит синтез ( репликация ) ДНК, изучаемый путём регистрации включенных в Я. меченных радиоактивными изотопами предшественников ДНК (обычно тимидина). Показано, что по длине хроматиновых фибрилл имеется множество участков (так называемых репликонов), каждый со своей точкой начала синтеза ДНК, от которой репликация распространяется в обе стороны. Вследствие репликации ДНК удваиваются и сами хромосомы.
В хроматине Я. происходит считывание закодированной в ДНК генетической информации путём синтеза на ДНК молекул матричной, или информационной, РНК (см. Транскрипция), а также молекул других типов РНК, участвующих в белковом синтезе. Специальные участки хромосом (и соответственно хроматина) содержат повторяющиеся гены, которые кодируют молекулы рибосомной РНК; в этих местах Я. формируются богатые рибонуклеопротеидами (РНП) ядрышки, основная функция которых — синтез РНК, входящей в состав рибосом. Наряду с компонентами ядрышка в Я. есть и другие виды частиц РНК. К ним относятся перихроматиновые фибриллы толщиной 3—5 нми перихроматиновые гранулы (ПГ) диаметром 40—50 нм, расположенные на границах зон рыхлого и компактного хроматина. И те и другие, вероятно, содержат матричную РНК в соединении с белками, а ПГ отвечают её неактивной форме; наблюдался выход ПГ из Я. в цитоплазму через поры ЯО. Имеются также интерхроматиновые гранулы (20—25 нм), а иногда и толстые (40—60 нм) нити РНП, скрученные в клубки. В ядрах амёб имеются нити РНП, скрученные в спирали (30—35 нмх 300 нм); спирали могут выходить в цитоплазму и, вероятно, содержат матричную РНК. Наряду с ДНК- и РНК-содержащими структурами некоторые Я. содержат чисто белковые включения в виде сфер (например, в Я. растущих яйцеклеток многих животных, в Я. ряда простейших), пучков фибрилл или кристаллоидов (например, в ядрах многих тканевых клеток животных и растений, макронуклеусах ряда инфузорий). В Я. обнаружены также фосфолипиды, липопротепды, ферменты (ДНК-полимераза, РНК-полимераза, комплекс ферментов оболочки Я., в том числе аденозинтрифосфатаза, и др.).
В природе встречаются различные специальные типы Я.: гигантские Я. растущих. яйцеклеток, особенно рыб и земноводных; Я., содержащие гигантские политенные хромосомы (см. Политения ), например в клетках слюнных желёз двукрылых насекомых; компактные, лишённые ядрышек Я. сперматозоидов и микронуклеусы инфузорий, сплошь заполненные хроматином и не синтезирующие РНК; Я., в которых хромосомы постоянно конденсированы, хотя ядрышки образуются (у некоторых простейших, в ряде клеток насекомых); Я., в которых произошло дву- или многократное увеличение числа наборов хромосом ( полиплоидия ; фигуры 7, 9 ).
Основной способ деления Я. — митоз, характеризующийся удвоением и конденсацией хромосом, разрушением ЯО (исключение — многие простейшие и грибы) и правильным расхождением сестринских хромосом в дочерние клетки. Однако Я. некоторых специализированных клеток, особенно полиплоидные, могут делиться простой перешнуровкой (см. Амитоз ) .Высокополиплоидные Я. могут делиться не только на 2, но и на много частей, а также почковаться ( фигура 7 ). При этом может происходить разделение целых хромосомных наборов (т. н. сегрегация геномов).
Лит.:Руководство по цитологии, т. 1, М. —Л., 1965; Райков И. Б., Кариология простейших, Л., 1967; Робертис Э., Новинский В., Саэс Ф.,. Биология клетки, пер. с англ., М., 1973; Ченцов Ю. С., Поляков В. Ю., Ультраструктура клеточного ядра, М., 1974; The nucleus, ed. A. J. Dalton, F, Haguenau, N. Y. — L., 1968; The cell nucleus, ed. Н. Busch, v. 1—3, N. Y. — L., 1974.
И. Б. Райков.