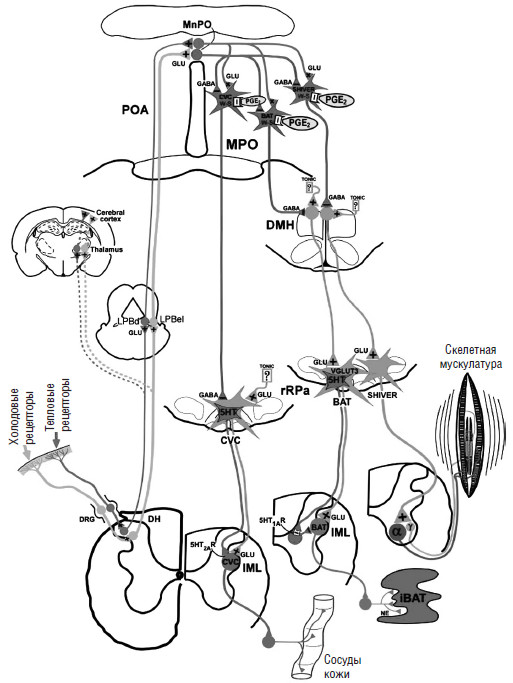
Рис. 13. Схема контроля терморегуляции (Morrison S. F., Nakamura К., 2011) «Тепловые» или «холодовые» сенсорные рецепторы передают информацию в соответствующие сенсорные нейроны дорсальных ганглиев (DRG). Далее информация «поступает» к тепловым сенсорным нейронам второго порядка, находящимся в дорсальных рогах спинного мозга (DH). Холодовые сенсорные глутаматчувствительные DH-нейроны активируют нейроны третьего порядка, находящиеся во внешнем боковом подъядре латерального парабрахиального ядра (LPBel). Нейроны третьего порядка «тепловых» сенсорных нейронов находятся в дорсальном подъядре латерального парабранхиального ядра (LPBd). Термосенсорные сигналы от нейронов спинного мозга (DH) передаются в таламус, а далее в кору больших полушарий для сознательного восприятия тепловой локализации. Термосенсорные сигналы, для возникновения терморегуляторного ответа, передаются от нейронов третьего порядка (LPB) в преоптическую область (РОА). В ней ГАМК-ергические (GABA) интернейроны, находящиеся в медиальном преоптическом подъядре (МпРО), активируются вводом глутамата из «холод»-активированных нейронов, локализующихся в парабранхиальном ядре (LPB), и ингибируют различные нейроны, чувствительные к «тепловым» стимулам (W-S) медиального преоптического подъядра (МРО), контролирующие вазоконстрикцию сосудов кожи (CVC), бурую жировую ткань (ВАТ), сократимость скелетных мышц (озноб). С другой стороны, глута-матергические интернейроны в медиальном преоптическом подъядре (МпРО), активируются глутаматом из «тепло»-активированных нейронов, находящихся в латеральном парабронхиальном ядре. Простагландин Е2 (PGE2) связывается с простагландиновыми рецепторами 3 (ЕРЗ г) на «тепло»-чувствительных нейронах в преоптической области (РОА), ингибируя их активность. Преоптические «тепло»-чувствительные нейроны обеспечивают контроль терморегуляции (вазоконстрикцию сосудов кожи (CVC)), симпатическими премоторными нейронами в продолговатом мозге, включающими ростральный бледный шов (rRPa) и симпатические преганглионарные нейроны интермедиолатерального ядра (IML). Премоторные нейроны могут увеличивать влияние симпатической НС выбросом глутамата и/или серотонина в пределах интермедиолатерального ядра (IML). Преоптические «тепло»-чувствительные нейроны, обеспечивающие терморегуляторный контроль термогенеза бурой жировой ткани, ингибируют нейроны гипоталамуса, активирующиеся во время охлаждения кожи. Некоторые премоторные нейроны, содержащие везикулярный глутаматный переносчик, могут выбрасывать глутамат, активирующий симпатические преганглионарные нейроны. Другие нейроны выбрасывают серотонин (5-НТ), взаимодействующий с 5-НТ1А рецепторами на нейронах, находящихся в интермедиолатеральном ядре (IML) и регулирующих термогенез. Преоптические «тепло»-чувствительные нейроны обеспечивают терморегуляторный контроль, обеспечивают контроль сократительного термогенеза, активируя альфа– и гамма-мотонейроны в скелетных мышцах.
Центральные механизмы регуляции теплообмена включают в себя центр терморегуляции, локализующийся в медиальной преоптической области переднего и заднем гипоталамусе. Именно этот центр устанавливает равновесие между теплопродукцией и теплоотдачей через эфферентные нейроны заднего гипоталамуса. В центре терморегуляции имеются неодинаковые по функциям нейроны, соответственно реагирующие на различные БАВ (ацетилхолин, серотонин, норадреналин и др.). Так, нейроны, расположенные в переднем гипоталамусе и «задающие» уровень поддерживаемой в организме температуры тела, реагируют на ацетилхолин, а также соотношение концентраций ионов натрия и кальция. Поэтому патологические процессы, приводящие к изменению концентрации указанных электролитов (например, генерализованный инфекционный процесс или тяжелая интранатальная асфиксия), будут сопровождаться нарушениями температурного баланса. По нашему мнению, этот факт необходимо учитывать в повседневной клинической практике, особенно у ургентных больных. Поскольку переливание гиперосмолярных растворов, а, к сожалению, это бывает очень часто, приводит к нарушению одного из основных законов организма (закона изоосмолярности) и, как следствие, к гипертермии, что иногда трактуется врачами, как «присоединение инфекции» с последующими не вполне однозначными действиями (назначение массивной антибактериальной терапии, введение иммуноглобулинов и т. д.).
Доказано (Elmquist J. К. et al., 1997; Matsumura К. et al., 1998; Yamaga-ta K. et al., 2001) что при физиологических условиях в центральных механизмах терморегуляции простагландины и цитокины существенного значения не имеют. Но при патологических процессах, особенно при лихорадке или гипотермии (последнее у новорожденных), они могут изменять уровень поддерживаемой температуры тела. Важную роль в изменении температуры играют простагландины Е (Stitt J. Т. et al., 1973).
По нашему мнению, эти наблюдения редко используются в неонатальной клинической практике при трактовке состояния больного. Вероятно, напрасно. Так, многочисленные наблюдения подтверждают склонность недоношенных детей к гипотермии, в том числе и при развитии сепсиса. Механизмов развития снижения температуры тела у них много, но продемонстрировано (Fairchild К. D. et al., 2003), что у новорожденных детей, особенно недоношенных, более низкая, чем у взрослых, продукция ИЛ-1β и ФНОа. Кроме того, рецепторы к указанным цитокинам у детей менее чувствительны, чем у взрослых, что является одним из механизмов, объясняющих склонность недоношенных к гипотермии. Другими словами, отмечается не существующий на самом деле парадокс: уровень указанных провоспалительных цитокинов может быть высок, наблюдается «цитокиновый шторм», а выраженной температурной реакции нет. Вообще это достаточно распространенный феномен в неонатальной медицине, патогенетически связанный с поддержанием баланса в различных системах. Например, если рассмотреть такое часто встречающееся заболевание, как ГБН, а именно желтушную форму этой болезни. Иногда, врачей удивляет тот факт, что при гемолитической болезни, когда идет интенсивный гемолиз – анемии нет. При этом они забывают, что одновременно с разрушением эритроцитов происходит их синтез. И если синтез и разрушение (гемолиз) уравновешены, то желтуха будет, а анемия – нет. Или почему при назначении эритропоэтина анемия не купируется? Дело в том, что мало наличия эритропоэтина, мало наличия клетки, еще на клетке должен быть чувствительный рецептор. А если рецептора нет, в силу генетической причины, или он не чувствителен, то и эритропоэтин не поможет: не на что действовать… То же самое относится ко всем эндогенно вводимым веществам, например сурфактанту.
Эффекторные механизмы теплообмена включаются, если не совпадают величины температур, «установленные» центром терморегуляции и периферической температурой тела. Это несоответствие приводит к изменению просвета сосудов (изменению кровотока), изменению частоты сердечных сокращений и/или корректировке потоотделения симпатической нервной системой. Указанные механизмы, плохо развитые у детей, включаются при необходимости увеличить теплоотдачу.
Когда же, несмотря на вазоконстрикцию и минимальное потоотделение, уровень температуры ниже, чем «установленный» центром терморегуляции, активизируются процессы теплопродукции (терморегуляционная мышечная активность и липолиз). В повышении теплопродукции быстрое, но непродолжительное влияние оказывают адреналин и норадреналин (что необходимо учитывать при назначении инотропной терапии и трактовке появившейся гипертермии). Более продолжительное усиление обменных процессов достигается под влиянием тироксина и трийодтиронина, синтез которых снижен у глубоконедоношенных детей (см. рисунок 14 и таблицы 4 и 5 (Петренко Ю. В., 1995)) и ряде патологических состояний у доношенных новорожденных. Например, описанном нами, совместно с Н. П. Шабаловым и Н. Н. Шабаловой в 2002 году, гипоэргическом варианте неонатального сепсиса.
Конец бесплатного ознакомительного фрагмента