Страница:
Одно из существенных отличий экосистем древних (палеозойских и мезозойских) озер от современных – это полное отсутствие погруженных макрофитов, за исключением харовых водорослей (которые депонируют биогены почти на порядок хуже, чем покрытосеменные). Основными продуцентами там скорее всего были плавающие маты, причем в мезозое получили распространение водорослевые маты на каркасе из высших растений. В триасе это были плевромейи – крупные плавающие плауновидные (что-то вроде сильно увеличенного полушника), по виду напоминающие морской буй, где в роли верхней сигнальной лампочки выступает пучок листьев и спорангии; листья, скорее всего, плавали по поверхности воды, постепенно отгнивая с вершины.[49] На каркасе из этих плауновидных возникали плавающие острова из мелких водяных мхов и печеночников (типа любимой аквариумистами риччии), а снаружи все это обрастало зелеными водорослями и цианобактериями. На этих матах существовала богатая фауна беспозвоночных: в палеозое – сходные с многоножками растительноядные эвтикарциноиды, позже – ракообразные и насекомые.
Поскольку все фотосинтезирующие структуры были сосредоточены на поверхности озера (а дно было к тому же еще и затенено), гиполимнион почти всегда был асфиксным и безжизненным, а озера легко эвтрофицировались. Однако в непосредственной близости от плавающего мата концентрация кислорода в воде могла быть очень высока. Возможно, именно этим объясняется присутствие в составе фауны эвтрофных (судя по составу отложений) мезозойских озер целого ряда групп насекомых, являющихся ныне реофилами (обитателями быстрых, богатых кислородом рек и ручьев). Кроме того, в составе этих фаун комбинируются как пресноводные формы, имеющие адаптации к водному образу жизни, так и наземные (которые могли «ходить пешком» по поверхности плавающего мата). Эти имевшие огромную биомассу и богатые животным белком «плавучие острова» могли составлять основу питания для многих позвоночных, включая и динозавров. Окончательно экосистемы такого типа исчезли лишь в кайнозое, когда на дне водоемов возникли заросли погруженных макрофитов из покрытосеменных (типа элодеи), резко увеличившие разнообразие и устойчивость пресноводных экосистем. Это и дало А. Г. Пономаренко основание разделить всю историю континентальных водоемов на два этапа: первый – от докембрия до середины палеогена (когда основными продуцентами были маты, сперва прикрепленные, затем плавающие), а второй – от середины палеогена до настоящего времени (когда эту роль стали играть погруженные макрофиты).
Глава 10
Поскольку все фотосинтезирующие структуры были сосредоточены на поверхности озера (а дно было к тому же еще и затенено), гиполимнион почти всегда был асфиксным и безжизненным, а озера легко эвтрофицировались. Однако в непосредственной близости от плавающего мата концентрация кислорода в воде могла быть очень высока. Возможно, именно этим объясняется присутствие в составе фауны эвтрофных (судя по составу отложений) мезозойских озер целого ряда групп насекомых, являющихся ныне реофилами (обитателями быстрых, богатых кислородом рек и ручьев). Кроме того, в составе этих фаун комбинируются как пресноводные формы, имеющие адаптации к водному образу жизни, так и наземные (которые могли «ходить пешком» по поверхности плавающего мата). Эти имевшие огромную биомассу и богатые животным белком «плавучие острова» могли составлять основу питания для многих позвоночных, включая и динозавров. Окончательно экосистемы такого типа исчезли лишь в кайнозое, когда на дне водоемов возникли заросли погруженных макрофитов из покрытосеменных (типа элодеи), резко увеличившие разнообразие и устойчивость пресноводных экосистем. Это и дало А. Г. Пономаренко основание разделить всю историю континентальных водоемов на два этапа: первый – от докембрия до середины палеогена (когда основными продуцентами были маты, сперва прикрепленные, затем плавающие), а второй – от середины палеогена до настоящего времени (когда эту роль стали играть погруженные макрофиты).
Глава 10
Поздний палеозой – ранний мезозой: эволюция наземных позвоночных (1). Анамнии и амниоты. Две линии амниот – тероморфная и завроморфная.
Мы расстались с самыми первыми тетраподами (девонской ихтиостегой и родственными ей формами), когда те... Чуть было не сказал: «...делали первые шаги по суше». Но мы ведь теперь знаем, что те существа были чисто водными. Поэтому выражусь более осторожно: «...получили принципиальную возможность периодически покидать водную среду». Да и что они, собственно говоря, могли бы найти на той суше, кроме перегрузок из-за многократного увеличения собственного веса, теплового удара да смертоносного обезвоживания организма?.. Пищу? Какую пищу – в девонском-то периоде? В общем, в девоне целенаправленный «выход на сушу» был бы даже не авантюрой, а чем-то по разряду мазохизма. Есть пятипалая конечность, позволяющая при необходимости переползти из совсем уже пересохшей лужи в не совсем пересохшую, и ладно...
Ситуация принципиально изменилась с начала карбона, когда в уже знакомых нам «лесах-водоемах» возникла богатая фауна из амфибиотических и наземных членистоногих (паукообразные, многоножки, а потом и насекомые); за этот ресурс уже стоило побороться. Среди карбоновых амфибий повляются небольшие (менее 1 м), похожие на современных саламандр антракозавры, имеющие несомненные приспособления к наземной жизни и явно ориентированные на питание беспозвоночными (а не рыбой, как их более крупные родственники). Именно антракозавров считают предками рептилий[50] ; первые рептилии – карбоновые капториниды – напоминали по внешнему виду крупных ящериц, причем, судя по строению их челюстного аппарата, они специализировались именно на питании насекомыми. Впрочем, не будем забегать вперед.
Чем объяснить итоговый эволюционный неуспех амфибий? Ведь этот класс процветал лишь в карбоновом периоде (причем только на территории тогдашних тропиков, в Еврамерии; ни в Ангариде, ни в Гондване по сию пору не найдено ни единой косточки карбонового возраста[51] ), когда у него не было конкурентов. А во все остальные времена амфибии занимали сугубо подчиненное положение в экосистемах – как наземных, так и пресноводных. Судя по всему, амфибий сгубила даже не облигатная связь с водой (они не выработали яйца? и размножаются икрой), а неисправимое несовершенство их дыхательной системы, предопределившее целую цепь пренеприятнейших анатомо-физиологических следствий. Давайте по порядку.
Из курса зоологии вы должны помнить, что лягушка не имеет ни ребер, ни диафрагмы. Поэтому она (в отличие от нас) – неспособна делать вдох, увеличивая объем замкнутой грудной полости, и вынуждена нагнетать воздух в легкие крайне несовершенным способом: набирать его в ротовую полость, замыкать ее, а потом «проглатывать» воздух, поднимая дно ротовой полости и сокращая ее объем. Итак, легочное дыхание у амфибий развито слабо.
Слабость легочного дыхания не позволяет амфибиям освободить от дыхательных функций кожу. Об этом обычно упоминают в той связи, что земноводные должны всегда сохранять покровы влажными (именно в этой влаге и растворяется воздушный кислород, «всасываемый» затем кожей), а потому они навсегда прикованы к воде. Однако это еще полбеды. Хуже другое: не освободив от дыхательных функций кожу, невозможно изолировать друг от друга большой и малый круги кровообращения и отделить венозную кровь от артериальной. Появление легких у тетрапод привело к возникновению двух кругов кровообращения; при этом правая сторона сердца становится «венозной», а левая – «артериальной». У амфибий сердце трехкамерное – два предсердия и желудочек; у рептилий (формально!) тоже трехкамерное, но в желудочке возникает неполная продольная перегородка, делящая его на «венозную» и «артериальную» половины. В ходе дальнейшей эволюции перегородка становится полной, а сердце – четырехкамерным (хотя у крокодилов, птиц и млекопитающих это происходит по-разному). У амфибий же кислород приносят в сердце вены, идущие не только от легких (малый круг), но и от кожи (большой круг), поэтому бессмысленно создавать какую-либо перегородку между правой и левой половинами желудочка: кровь в сердце всегда будет смешанной – артериально-венозной. Пока кровь не разделена на артериальную и венозную, уровень энергетического обмена организма будет принципиально невысоким. К тому же при постоянно влажных покровах любое повышение температуры тела будет тут же сводиться на нет за счет поверхностного испарения (термическое охлаждение). Все это полностью закрывает амфибиям путь к достижению любых выриантов эндотермии («теплокровности»)[52] – с понятными эволюционными последствиями.
Поскольку единственно доступным для амфибий способом наполнения легких является «проглатывание» воздуха (гулярное дыхание), им необходимо сохранять «подчелюстной насос», поднимающий и опускающий дно ротовой полости. Его размещение требует специфической («лягушачьей») формы черепа – очень широкой и приплюснутой. При этом челюстная мускулатура оказывается размещенной крайне невыгодным образом: ее можно подсоединить лишь к самому челюстному суставу, с задней его стороны. Получающийся в результате рычаг будет крайне неэффективен – все равно как если бы мы затворяли дверь, упираясь не в ручку, а около петель. Для того чтобы развивать при помощи таких челюстей сколь-нибудь приличные усилия, приходится наращивать объем челюстной мускулатуры; в итоге эта мускулатура занимает практически все пространство черепа, не оставляя места для мозговой коробки. Следовательно, амфибии – существа принципиально «безмозглые», и это закрывает им путь к сложным формам поведения.
Понятно, что бороться (в эволюционном смысле) со всем этим комплексом неприятностей надо путем радикальной перестройки дыхательной системы. Анализируя переход от амфибий к рептилиям, Н. Н. Иорданский (1977) выдвинул свой принцип ключевого ароморфоза: изменение в одной системе органов ведет к целому каскаду последовательных прогрессивных перестроек всего организма.
Достаточно наладить нормальное легочное дыхание (изменив объем грудной полости), чтобы приведенные выше цепочки начали «раскручиваться» в обратную сторону. Убрав «подчелюстной насос», можно сделать череп высоким и узким, подвести жевательную мускулатуру к челюсти не сзади, а сверху (как у нас с вами), уменьшить – за счет улучшения рычага – ее объем и отдать освободившееся место под «мозги». Кроме того, такое расположение жевательных мышц позволит в дальнейшем не только удерживать схваченную добычу, но и пережевывать пищу (рис. 37). Если освободить кожу от дыхательных функций, то будет возможность разделить круги кровообращения и резко интенсифицировать обмен веществ. Эти инженерные решения вполне однозначны, равно как и необходимость одеть икринку особой оболочкой (амнионом), как бы создающей для зародыша маленький искусственный водоем и делающей его развитие независимым от водной среды. Отсюда фундаментальное разделение позвоночных на «прикованных к воде» анамний (рыб и амфибий) и «истинно сухопутных» амниот (рептилий, птиц и млекопитающих). А вот дальше начинается интереснейший эволюционный выбор: вариантов – два, в обоих есть свои плюсы и минусы.

Создав сухую кожу с роговым покрытием, мы сводим к минимуму потери влаги: организм становится практически независимым от внешних источников воды (как одетые в «пустынные скафандры» герои фантастического романа Ф. Херберта «Дюна»). Однако за удобство надо платить.
Во-первых, необходимо перестроить выделительную систему. Почки амфибий функционально не отличаются от рыбьих и предназначены для выведения из организма избытка воды (которую тот постоянно «насасывает» за счет осмоса из гипотонической внешней среды). Проблема удаления конечного продукта белкового обмена – весьма токсичной мочевины CO(NH2)2 – решается в этом случае элементарно: ее просто растворяют в водяном потоке, который так и так постоянно «течет сквозь организм». Иное дело, когда мы начинаем экономить воду, потребляя ее по минимуму. (Легко создать канализацию в населенном пункте, стоящем на реке. А что делать, если в твоем распоряжении лишь артезианский колодец?) Тогда «почки выведения» необходимо заменить на «почки сбережения», призванные выводить во внешнюю среду как можно меньше воды. При этом приходится менять конечный продукт белкового обмена с мочевины на менее токсичную мочевую кислоту, а этот дополнительный «технологический цикл» весьма энергоемок.[53]
Во-вторых (и это даже более важно), сухая, лишенная желез кожа создает большие трудности с терморегуляцией; при жизни на суше, где обычны резкие температурные перепады, эта проблема – одна из важнейших. С голыми, ороговевшими покровами нелегко осуществлять и теплоизоляцию (в холод), и теплосброс (в жару). Вот почему для этой эволюционной линии путь к достижению гомойотермии (поддержанию постоянной, независимой от внешней среды температуры тела) если и не закрыт полностью, то очень сильно затруднен.
Отказавшись от экономии воды и оставив кожу железистой, не ороговевшей, мы получим много новых возможностей. Кожные железы можно превратить в волоски, выполняющие осязательные функции (а сделав эти волоски достаточно густыми, создать теплоизолирующий покров – шерсть), или в дополнительный орган выделения – потовые железы, которые являются еще и терморегулятором (пот, испаряясь, охлаждает поверхность тела), или в млечные (изменив состав их выделений),чтобы выкармливать с их помощью детенышей. В этой эволюционной линии с ее терморегуляторными возможностями появление гомойотермии просто напрашивается. Так оно и было на самом деле. Правда, по степени зависимости от источников воды эти существа не слишком превосходили амфибий, но ведь за все в этом мире приходится платить. А с другой стороны, раз уж внутренняя среда организма все равно будет существовать как бы в постоянном водяном токе, нет нужды радикально перестраивать почки на «водосберегающую технологию» и возиться со сменой конечного продукта азотного обмена!
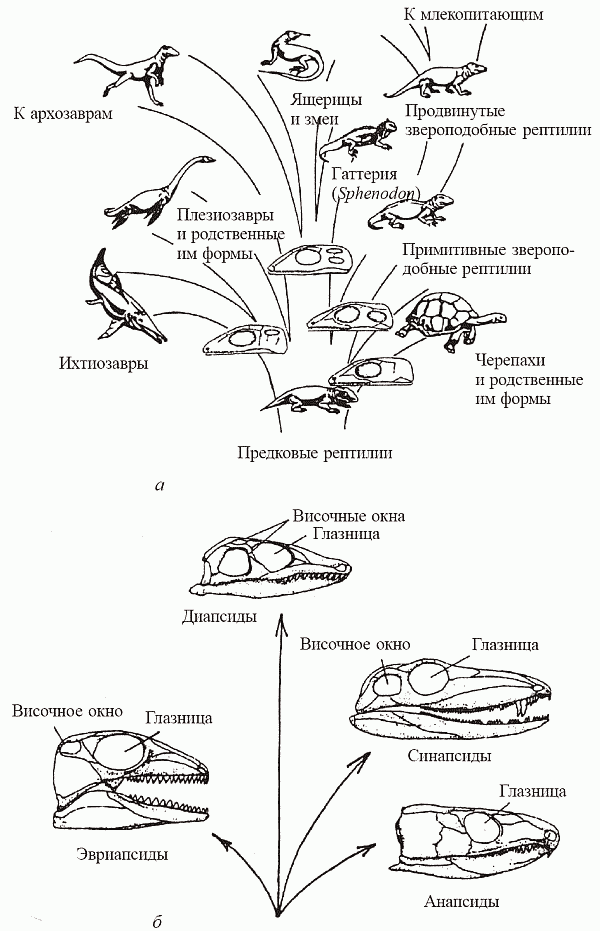
Что же это за «две эволюционные линии»? Традиционный взгляд на эволюцию рептилий – он и сегодня отражен во многих учебниках – предполагает, что от каких-то антракозавров (здесь часто называют сеймурию, имевшую уже рептилийное строение челюсти и грудной клетки, пригодной для реберного дыхания, но еще не расставшуюся с водой – личинки ее так и оставались «головастиками» с наружными жабрами) происходит некий гипотетический «общий предок пресмыкающихся», который обладает уже всем набором рептилийных признаков (водонепроницаемой кожей, почками сбережения и т.д.). От него-то и берут начало основные эволюционные ветви этого класса – анапсиды, синапсиды, диапсиды и эвриапсиды; этим группам зачастую придают статус подклассов. Они выделены на основе того, каким способом в каждом из них трансформировался исходный стегальный (крышевидный) череп лабиринтодонтов (рис. 38). У анапсид (сюда относятся черепахи и ряд вымерших групп, например, парейазавры) сохраняется исходный тип черепа – массивный, лишенный облегчающих конструкцию отверстий. У синапсид – «зверообразных ящеров», чьи зубы были дифференцированы по форме и функциям (пеликозавров и более продвинутых терапсид, которых традиционно считают предками млекопитающих), возникает одно височное окно, от краев которого отходят челюстные мышцы, и единственная скуловая дуга. У диапсид же возникают два височных окна и, соответственно, две скуловые дуги – верхняя и нижняя. К этой группе принадлежит большинство рептилий, как современных (ящерицы со змеями, гаттерии и крокодилы), так и вымерших (динозавры, птерозавры). К эвриапсидам относят несколько вымерших морских групп (ихтиозавры, плезиозавры и родственные им нотозавры, плакодонты); их черепа имеют единственное височное окно, но образуется оно иным, чем у синапсид, образом.[54] Необходимо заметить, что черепа млекопитающих и птиц могут быть отнесены соответственно к синапсидному и диапсидному типам – что отражает происхождение этих высших тетрапод.
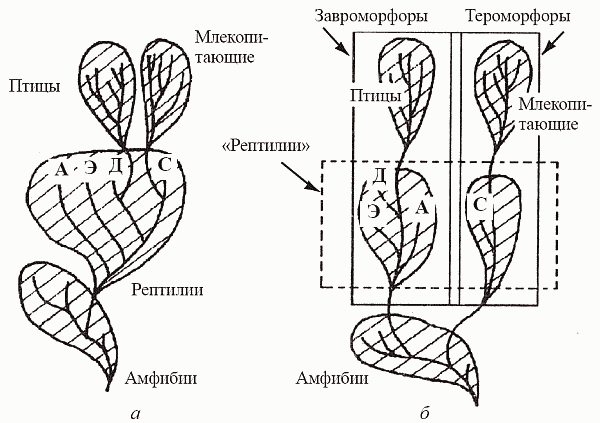
Так вот, по наиболее современным представлениям (они, как часто бывает в науке, могут считаться развитием на новом уровне взглядов Гексли), в нескольких (минимум в двух) эволюционных линиях амфибий независимо выработался синдром[55] рептилийных признаков и был достигнут рептилийный уровень организации. Выходит, «рептилии» – это категория не вертикальной, а горизонтальной классификации; это не таксон, члены которого связанны единством происхождения, а града – уровень организации, достигаемый тетраподами при утере ими облигатной связи с водой (рис. 39). Существование двух независимых эволюционных ветвей амниот – тероморфной (от греческого th?rion – зверь) и завроморфной (от греческого sauros – ящер), разошедшихся еще на уровне амфибий и венчаемых: первая – млекопитающими, а вторая – птицами и динозаврами, сейчас практически общепризнано[56] . Собственно говоря, все 300 млн лет существования наземных тетрапод – это история состязания теро-морфов с завроморфами, где эволюционный успех сопутствовал то одним, то другим.
Основными фитофагами на суше являлись насекомые, но вначале они, как мы помним из предыдущей главы, только «снимали сливки» – потребляли одни высококалорийные генеративные части растений (пыльцу и семезачатки). В перми насекомые освоили питание соками растений, высасывая их при помощи хоботка из проводящих пучков (цикады и родственные им формы из отряда полужесткокрылых), а также древесиной, уже подвергшейся первичному грибному разложению (жуки). Однако питаться низкокалорийными живыми вегетативными тканями растений они стали лишь в мезозое. В карбоне (и даже еще раньше, в девоне) существовали другие потребители этого (прямо скажем, не особо привлекательного) ресурса: крупные двупарноногие многоножки; именно ими, как считают, оставлены погрызы на ископаемых листьях из палеозойских отложений. Пик разнообразия и обилия этих членистоногих пришелся на поздний карбон, а затем все они бесследно исчезли (возможно, разделив судьбу гигантских насекомых – см. предыдущую главу). Эту-то «вакансию» и заполнили первые позвоночные-фитофаги. Именно в раннепермских «пеликозавровых» сообществах впервые складывается нормальное для современности численное соотношение между растительноядными и хищниками (до того все тетраподы были либо рыбоядными, либо насекомоядными, т.е. хищниками, консументами второго порядка).
Мы уже говорили о том, что анатомия тероморфов просто-таки подталкивает их к различного рода эволюционным экспериментам с активной терморегуляцией. И хотя пеликозавры, без сомнения, были существами вполне «холоднокровными» (пойкилотермными) и метаболизм имели еще типично рептилийный, первые опыты по части температурного контроля начались уже на этом уровне. У многих из них возник «парус» – высокие остистые отростки позвоночника, заключенные в единую пластину из мягких тканей; в бороздах у основания отростков проходили крупные кровеносные сосуды. Парус, имевший огромную поверхность при малом объеме, служил, как сейчас считают, для быстрого поглощения или отдачи тепла телом. Расчеты показали, что особь пеликозавра массой 200 кг с парусом нагревается с 26 до 32°С за 80 мин (против 205 мин без паруса). По всей видимости, это позволяло пеликозаврам активизироваться по утрам значительно раньше добычи или других хищников сравнимого размера. С другой стороны, парус мог способствовать рассеиванию тепла в полуденное время, оберегая животное от перегрева. Характерно, что парус возникал как в хищных, так и в растительноядных группах пеликозавров, т.е. многократно и независимо. В дальнейшем сходные структуры изредка появлялись у динозавров (опять-таки как у хищных – спинозавр, так и у растительноядных – стегозавр), однако в целом такой способ терморегуляции оказался явно тупиковым.
По иному решили эту проблему более продвинутые тероморфы из отряда терапсид, появившиеся в поздней перми. Среди них были и специализированные растительноядные формы вроде дицинодонтов, достигавших размеров быка и имевших выступающие «моржовые» клыки в верхней челюсти (рис. 40, б), и крупные хищники – такие, как горгонопсиды, чем-то похожие на саблезубого тигра (рис. 40, в и г).

Однако сейчас полагают, что продвинутые терапсиды гораздо более походили на млекопитающих: доказано, что они, в отличие от прочих рептилий, имели развитое обоняние, слюнные железы и жесткие «усы»-вибри?ссы – осязательные волоски на морде (это можно установить исходя из того, как проходили поверхностные кровеносные сосуды черепа), а также «маммальное» строение поясов конечностей (об этом чуть ниже). Это были довольно «башковитые» создания – их коэффициент цефализации (отношение веса мозга к весу тела) заметно выше, чем у современных им завроморфов. Предполагают, что по уровню метаболизма эти животные не уступали наиболее примитивным млекопитающим – однопроходным (утконос и ехидна). Судя по всему, высшие терапсиды откладывали яйца, а детенышей выкармливали выделениями специальных желез – в точности, как однопроходные. Они были, скорее всего, покрыты шерстью (шерсть – это не что иное, как «расползшиеся» на всю поверхность тела и ставшие достаточно густыми осязательные вибрисы), а значит, являлись теплокровными: иначе зачем нужен теплоизолирующий покров?[57] Именно в одной из групп высших терапсид – цинодонтов (рис. 40, д) – в следующем, триасовом, периоде начался процесс, названный «параллельной маммализацией» (см. главу 11). Он, этот процесс, привел в конечном счете к возникновению настоящих млекопитающих.
Ситуация принципиально изменилась с начала карбона, когда в уже знакомых нам «лесах-водоемах» возникла богатая фауна из амфибиотических и наземных членистоногих (паукообразные, многоножки, а потом и насекомые); за этот ресурс уже стоило побороться. Среди карбоновых амфибий повляются небольшие (менее 1 м), похожие на современных саламандр антракозавры, имеющие несомненные приспособления к наземной жизни и явно ориентированные на питание беспозвоночными (а не рыбой, как их более крупные родственники). Именно антракозавров считают предками рептилий[50] ; первые рептилии – карбоновые капториниды – напоминали по внешнему виду крупных ящериц, причем, судя по строению их челюстного аппарата, они специализировались именно на питании насекомыми. Впрочем, не будем забегать вперед.
Чем объяснить итоговый эволюционный неуспех амфибий? Ведь этот класс процветал лишь в карбоновом периоде (причем только на территории тогдашних тропиков, в Еврамерии; ни в Ангариде, ни в Гондване по сию пору не найдено ни единой косточки карбонового возраста[51] ), когда у него не было конкурентов. А во все остальные времена амфибии занимали сугубо подчиненное положение в экосистемах – как наземных, так и пресноводных. Судя по всему, амфибий сгубила даже не облигатная связь с водой (они не выработали яйца? и размножаются икрой), а неисправимое несовершенство их дыхательной системы, предопределившее целую цепь пренеприятнейших анатомо-физиологических следствий. Давайте по порядку.
Из курса зоологии вы должны помнить, что лягушка не имеет ни ребер, ни диафрагмы. Поэтому она (в отличие от нас) – неспособна делать вдох, увеличивая объем замкнутой грудной полости, и вынуждена нагнетать воздух в легкие крайне несовершенным способом: набирать его в ротовую полость, замыкать ее, а потом «проглатывать» воздух, поднимая дно ротовой полости и сокращая ее объем. Итак, легочное дыхание у амфибий развито слабо.
Слабость легочного дыхания не позволяет амфибиям освободить от дыхательных функций кожу. Об этом обычно упоминают в той связи, что земноводные должны всегда сохранять покровы влажными (именно в этой влаге и растворяется воздушный кислород, «всасываемый» затем кожей), а потому они навсегда прикованы к воде. Однако это еще полбеды. Хуже другое: не освободив от дыхательных функций кожу, невозможно изолировать друг от друга большой и малый круги кровообращения и отделить венозную кровь от артериальной. Появление легких у тетрапод привело к возникновению двух кругов кровообращения; при этом правая сторона сердца становится «венозной», а левая – «артериальной». У амфибий сердце трехкамерное – два предсердия и желудочек; у рептилий (формально!) тоже трехкамерное, но в желудочке возникает неполная продольная перегородка, делящая его на «венозную» и «артериальную» половины. В ходе дальнейшей эволюции перегородка становится полной, а сердце – четырехкамерным (хотя у крокодилов, птиц и млекопитающих это происходит по-разному). У амфибий же кислород приносят в сердце вены, идущие не только от легких (малый круг), но и от кожи (большой круг), поэтому бессмысленно создавать какую-либо перегородку между правой и левой половинами желудочка: кровь в сердце всегда будет смешанной – артериально-венозной. Пока кровь не разделена на артериальную и венозную, уровень энергетического обмена организма будет принципиально невысоким. К тому же при постоянно влажных покровах любое повышение температуры тела будет тут же сводиться на нет за счет поверхностного испарения (термическое охлаждение). Все это полностью закрывает амфибиям путь к достижению любых выриантов эндотермии («теплокровности»)[52] – с понятными эволюционными последствиями.
Поскольку единственно доступным для амфибий способом наполнения легких является «проглатывание» воздуха (гулярное дыхание), им необходимо сохранять «подчелюстной насос», поднимающий и опускающий дно ротовой полости. Его размещение требует специфической («лягушачьей») формы черепа – очень широкой и приплюснутой. При этом челюстная мускулатура оказывается размещенной крайне невыгодным образом: ее можно подсоединить лишь к самому челюстному суставу, с задней его стороны. Получающийся в результате рычаг будет крайне неэффективен – все равно как если бы мы затворяли дверь, упираясь не в ручку, а около петель. Для того чтобы развивать при помощи таких челюстей сколь-нибудь приличные усилия, приходится наращивать объем челюстной мускулатуры; в итоге эта мускулатура занимает практически все пространство черепа, не оставляя места для мозговой коробки. Следовательно, амфибии – существа принципиально «безмозглые», и это закрывает им путь к сложным формам поведения.
Понятно, что бороться (в эволюционном смысле) со всем этим комплексом неприятностей надо путем радикальной перестройки дыхательной системы. Анализируя переход от амфибий к рептилиям, Н. Н. Иорданский (1977) выдвинул свой принцип ключевого ароморфоза: изменение в одной системе органов ведет к целому каскаду последовательных прогрессивных перестроек всего организма.
Достаточно наладить нормальное легочное дыхание (изменив объем грудной полости), чтобы приведенные выше цепочки начали «раскручиваться» в обратную сторону. Убрав «подчелюстной насос», можно сделать череп высоким и узким, подвести жевательную мускулатуру к челюсти не сзади, а сверху (как у нас с вами), уменьшить – за счет улучшения рычага – ее объем и отдать освободившееся место под «мозги». Кроме того, такое расположение жевательных мышц позволит в дальнейшем не только удерживать схваченную добычу, но и пережевывать пищу (рис. 37). Если освободить кожу от дыхательных функций, то будет возможность разделить круги кровообращения и резко интенсифицировать обмен веществ. Эти инженерные решения вполне однозначны, равно как и необходимость одеть икринку особой оболочкой (амнионом), как бы создающей для зародыша маленький искусственный водоем и делающей его развитие независимым от водной среды. Отсюда фундаментальное разделение позвоночных на «прикованных к воде» анамний (рыб и амфибий) и «истинно сухопутных» амниот (рептилий, птиц и млекопитающих). А вот дальше начинается интереснейший эволюционный выбор: вариантов – два, в обоих есть свои плюсы и минусы.
Рис. 37. Расположение жевательной мускулатуры: а – лабиринтодонт Ichthyostega; б —примитивная капториноморфная рептилия Paleothyrus (по Norman, 1994)
Главная проблема, которую нам предстоит решать теперь, когда осуществлен настоящий выход на сушу, – это экономия воды. Возникает вопрос: как нам быть с кожей, доставшейся в наследство от амфибий – мягкой и влажной, богатой железами? Можно ее полностью заизолировать, создав на поверхности водонепроницаемый роговой слой, а можно лишь модифицировать, сохранив ее основные характеристики. Оба решения вполне реализуемы, и каждое из них влечет за собой целую цепь физиологических следствий.Создав сухую кожу с роговым покрытием, мы сводим к минимуму потери влаги: организм становится практически независимым от внешних источников воды (как одетые в «пустынные скафандры» герои фантастического романа Ф. Херберта «Дюна»). Однако за удобство надо платить.
Во-первых, необходимо перестроить выделительную систему. Почки амфибий функционально не отличаются от рыбьих и предназначены для выведения из организма избытка воды (которую тот постоянно «насасывает» за счет осмоса из гипотонической внешней среды). Проблема удаления конечного продукта белкового обмена – весьма токсичной мочевины CO(NH2)2 – решается в этом случае элементарно: ее просто растворяют в водяном потоке, который так и так постоянно «течет сквозь организм». Иное дело, когда мы начинаем экономить воду, потребляя ее по минимуму. (Легко создать канализацию в населенном пункте, стоящем на реке. А что делать, если в твоем распоряжении лишь артезианский колодец?) Тогда «почки выведения» необходимо заменить на «почки сбережения», призванные выводить во внешнюю среду как можно меньше воды. При этом приходится менять конечный продукт белкового обмена с мочевины на менее токсичную мочевую кислоту, а этот дополнительный «технологический цикл» весьма энергоемок.[53]
Во-вторых (и это даже более важно), сухая, лишенная желез кожа создает большие трудности с терморегуляцией; при жизни на суше, где обычны резкие температурные перепады, эта проблема – одна из важнейших. С голыми, ороговевшими покровами нелегко осуществлять и теплоизоляцию (в холод), и теплосброс (в жару). Вот почему для этой эволюционной линии путь к достижению гомойотермии (поддержанию постоянной, независимой от внешней среды температуры тела) если и не закрыт полностью, то очень сильно затруднен.
Отказавшись от экономии воды и оставив кожу железистой, не ороговевшей, мы получим много новых возможностей. Кожные железы можно превратить в волоски, выполняющие осязательные функции (а сделав эти волоски достаточно густыми, создать теплоизолирующий покров – шерсть), или в дополнительный орган выделения – потовые железы, которые являются еще и терморегулятором (пот, испаряясь, охлаждает поверхность тела), или в млечные (изменив состав их выделений),чтобы выкармливать с их помощью детенышей. В этой эволюционной линии с ее терморегуляторными возможностями появление гомойотермии просто напрашивается. Так оно и было на самом деле. Правда, по степени зависимости от источников воды эти существа не слишком превосходили амфибий, но ведь за все в этом мире приходится платить. А с другой стороны, раз уж внутренняя среда организма все равно будет существовать как бы в постоянном водяном токе, нет нужды радикально перестраивать почки на «водосберегающую технологию» и возиться со сменой конечного продукта азотного обмена!
Что же это за «две эволюционные линии»? Традиционный взгляд на эволюцию рептилий – он и сегодня отражен во многих учебниках – предполагает, что от каких-то антракозавров (здесь часто называют сеймурию, имевшую уже рептилийное строение челюсти и грудной клетки, пригодной для реберного дыхания, но еще не расставшуюся с водой – личинки ее так и оставались «головастиками» с наружными жабрами) происходит некий гипотетический «общий предок пресмыкающихся», который обладает уже всем набором рептилийных признаков (водонепроницаемой кожей, почками сбережения и т.д.). От него-то и берут начало основные эволюционные ветви этого класса – анапсиды, синапсиды, диапсиды и эвриапсиды; этим группам зачастую придают статус подклассов. Они выделены на основе того, каким способом в каждом из них трансформировался исходный стегальный (крышевидный) череп лабиринтодонтов (рис. 38). У анапсид (сюда относятся черепахи и ряд вымерших групп, например, парейазавры) сохраняется исходный тип черепа – массивный, лишенный облегчающих конструкцию отверстий. У синапсид – «зверообразных ящеров», чьи зубы были дифференцированы по форме и функциям (пеликозавров и более продвинутых терапсид, которых традиционно считают предками млекопитающих), возникает одно височное окно, от краев которого отходят челюстные мышцы, и единственная скуловая дуга. У диапсид же возникают два височных окна и, соответственно, две скуловые дуги – верхняя и нижняя. К этой группе принадлежит большинство рептилий, как современных (ящерицы со змеями, гаттерии и крокодилы), так и вымерших (динозавры, птерозавры). К эвриапсидам относят несколько вымерших морских групп (ихтиозавры, плезиозавры и родственные им нотозавры, плакодонты); их черепа имеют единственное височное окно, но образуется оно иным, чем у синапсид, образом.[54] Необходимо заметить, что черепа млекопитающих и птиц могут быть отнесены соответственно к синапсидному и диапсидному типам – что отражает происхождение этих высших тетрапод.
Рис. 38. Филогения рептилий: а – родословное древо (в основании соответствующих ветвей – схематически изображенные типы черепа); б – происхождение четырех типов черепа (по Romer, 1950, и Norman, 1994)
Однако еще в середине прошлого века Т. Гексли, друг и сподвижник Ч. Дарвина, провел анатомическое сравнение четырех классов тетрапод, чтобы установить, кто чьим предком является. Глубокое сходство птиц и рептилий уже в те времена не вызывало особых сомнений; когда говорят, что «птицы – всего лишь продвинутая и сильно специализированная группа рептилий», это, в общем и целом, соответствует истине. Гексли обратил внимание и на менее очевидное, но весьма существенное сходство млекопитающих (маммалий) с амфибиями. Это сходство он видел в строении кожи (мягкой и богатой железами) и почек, выделяющих мочевину (у рептилий с птицами почки выделяют мочевую кислоту), а также в наличии двух затылочных мыщелков, которыми череп сочленяется с позвоночником (у рептилий с птицами – один). Но самым серьезным оказались для Гексли различия в строении кровеносной системы: у амфибий имеются две дуги аорты – правая и левая, у рептилий и птиц сохраняется только правая дуга, а у млекопитающих – только левая. Значит, «вывести» маммальную кровеносную систему из рептилийной (где левая дуга уже редуцирована) принципиально невозможно. На этом основании он заключил, что предками млекопитающих не могут быть рептилии (по крайней мере, нынеживущие) и маммалии должны вести свое происхождение прямо от амфибий.Так вот, по наиболее современным представлениям (они, как часто бывает в науке, могут считаться развитием на новом уровне взглядов Гексли), в нескольких (минимум в двух) эволюционных линиях амфибий независимо выработался синдром[55] рептилийных признаков и был достигнут рептилийный уровень организации. Выходит, «рептилии» – это категория не вертикальной, а горизонтальной классификации; это не таксон, члены которого связанны единством происхождения, а града – уровень организации, достигаемый тетраподами при утере ими облигатной связи с водой (рис. 39). Существование двух независимых эволюционных ветвей амниот – тероморфной (от греческого th?rion – зверь) и завроморфной (от греческого sauros – ящер), разошедшихся еще на уровне амфибий и венчаемых: первая – млекопитающими, а вторая – птицами и динозаврами, сейчас практически общепризнано[56] . Собственно говоря, все 300 млн лет существования наземных тетрапод – это история состязания теро-морфов с завроморфами, где эволюционный успех сопутствовал то одним, то другим.
Рис. 39. Обобщенные схемы филогении тетрапод: а – традиционная; б – современная. А – анапсиды; Д – диапсиды; С – синапсиды; Э – эвриапсиды
Первыми такого успеха добились тероморфы. Появившиеся еще в позднем карбоне пеликозавры – удивительные создания, многие из которых имели «парус» из остистых отростков позвонков (рис. 40, а) – составляли в ранней перми 70% всех амниот. Мощные клыки и непропорционально большая голова свидетельствуют о том, что пеликозавры были изначально ориентированы на питание крупной добычей; они были первыми хищными (в смысле плотоядными, а не насекомоядными) наземными существами. Тем интереснее то обстоятельство, что многие пеликозавры переходят к растительноядности – первый опыт фитофагии среди наземных позвоночных! Это чрезвычайно важно, ибо дефицит потребителей растительной массы в континентальных экосистемах должен был стать к тому времени просто угрожающим.Основными фитофагами на суше являлись насекомые, но вначале они, как мы помним из предыдущей главы, только «снимали сливки» – потребляли одни высококалорийные генеративные части растений (пыльцу и семезачатки). В перми насекомые освоили питание соками растений, высасывая их при помощи хоботка из проводящих пучков (цикады и родственные им формы из отряда полужесткокрылых), а также древесиной, уже подвергшейся первичному грибному разложению (жуки). Однако питаться низкокалорийными живыми вегетативными тканями растений они стали лишь в мезозое. В карбоне (и даже еще раньше, в девоне) существовали другие потребители этого (прямо скажем, не особо привлекательного) ресурса: крупные двупарноногие многоножки; именно ими, как считают, оставлены погрызы на ископаемых листьях из палеозойских отложений. Пик разнообразия и обилия этих членистоногих пришелся на поздний карбон, а затем все они бесследно исчезли (возможно, разделив судьбу гигантских насекомых – см. предыдущую главу). Эту-то «вакансию» и заполнили первые позвоночные-фитофаги. Именно в раннепермских «пеликозавровых» сообществах впервые складывается нормальное для современности численное соотношение между растительноядными и хищниками (до того все тетраподы были либо рыбоядными, либо насекомоядными, т.е. хищниками, консументами второго порядка).
Мы уже говорили о том, что анатомия тероморфов просто-таки подталкивает их к различного рода эволюционным экспериментам с активной терморегуляцией. И хотя пеликозавры, без сомнения, были существами вполне «холоднокровными» (пойкилотермными) и метаболизм имели еще типично рептилийный, первые опыты по части температурного контроля начались уже на этом уровне. У многих из них возник «парус» – высокие остистые отростки позвоночника, заключенные в единую пластину из мягких тканей; в бороздах у основания отростков проходили крупные кровеносные сосуды. Парус, имевший огромную поверхность при малом объеме, служил, как сейчас считают, для быстрого поглощения или отдачи тепла телом. Расчеты показали, что особь пеликозавра массой 200 кг с парусом нагревается с 26 до 32°С за 80 мин (против 205 мин без паруса). По всей видимости, это позволяло пеликозаврам активизироваться по утрам значительно раньше добычи или других хищников сравнимого размера. С другой стороны, парус мог способствовать рассеиванию тепла в полуденное время, оберегая животное от перегрева. Характерно, что парус возникал как в хищных, так и в растительноядных группах пеликозавров, т.е. многократно и независимо. В дальнейшем сходные структуры изредка появлялись у динозавров (опять-таки как у хищных – спинозавр, так и у растительноядных – стегозавр), однако в целом такой способ терморегуляции оказался явно тупиковым.
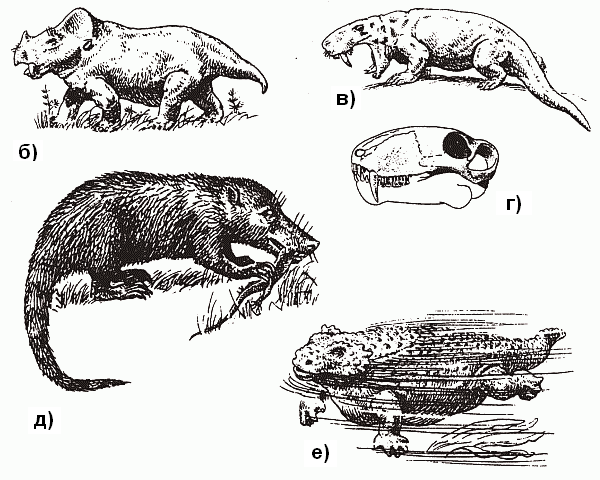
По иному решили эту проблему более продвинутые тероморфы из отряда терапсид, появившиеся в поздней перми. Среди них были и специализированные растительноядные формы вроде дицинодонтов, достигавших размеров быка и имевших выступающие «моржовые» клыки в верхней челюсти (рис. 40, б), и крупные хищники – такие, как горгонопсиды, чем-то похожие на саблезубого тигра (рис. 40, в и г).
а)
Рис. 40. Палеозойские рептилии: тероморфы (а–д) и анапсиды (е) (по Ивахненко и Корабельникову, 1986)
а – хищный пеликозавр Dimetrodon; б – растительноядный дицинодонт Dicinodontus; в – горгонопс Inostrancevia; г – его череп; д – продвинутый цинодонт Dvinia prima («почти млекопитающее»); е – парейазавр
Еще недавно всех их считали просто «ящерами» (ну, зубы там дифференцированы, экая важность!) и именно в таком виде они обычно изображены в книжках по палеонтологии.Однако сейчас полагают, что продвинутые терапсиды гораздо более походили на млекопитающих: доказано, что они, в отличие от прочих рептилий, имели развитое обоняние, слюнные железы и жесткие «усы»-вибри?ссы – осязательные волоски на морде (это можно установить исходя из того, как проходили поверхностные кровеносные сосуды черепа), а также «маммальное» строение поясов конечностей (об этом чуть ниже). Это были довольно «башковитые» создания – их коэффициент цефализации (отношение веса мозга к весу тела) заметно выше, чем у современных им завроморфов. Предполагают, что по уровню метаболизма эти животные не уступали наиболее примитивным млекопитающим – однопроходным (утконос и ехидна). Судя по всему, высшие терапсиды откладывали яйца, а детенышей выкармливали выделениями специальных желез – в точности, как однопроходные. Они были, скорее всего, покрыты шерстью (шерсть – это не что иное, как «расползшиеся» на всю поверхность тела и ставшие достаточно густыми осязательные вибрисы), а значит, являлись теплокровными: иначе зачем нужен теплоизолирующий покров?[57] Именно в одной из групп высших терапсид – цинодонтов (рис. 40, д) – в следующем, триасовом, периоде начался процесс, названный «параллельной маммализацией» (см. главу 11). Он, этот процесс, привел в конечном счете к возникновению настоящих млекопитающих.