Страница:
В интронах содержатся особые сигналы, которые узнаются специальными ферментами, осуществляющими сплайсинг. Так, в 98% случаев интроны начинаются с динуклеотида ГТ, а заканчиваются динуклеотидом АГ. Они–то и служат главными сигналами для правильного осуществления сплайсинга.
Во всем этом и состоит основное ноу–хау в организации генов человека и других эукариотических генов. «Лоскутное» устройство большинства генов – чрезвычайно важное эволюционное приобретение высших организмов. За счет сплайсинга в РНК может происходить соединение не только между соседними экзонными нуклео–тидными последовательностями, но и между другими, отстоящими порой в гене на значительном расстоянии. Это называют альтернативным сплайсингом (рис. 20). В геноме человека альтернативный сплайсинг характерен для более трети генов. Этому можно привести следующую аналогию: из слова «администрация» за счет удаления отдельных букв и слогов можно образовать множество совершенно разных по смыслу слов: ад, министр, амнистия, астра, нация. Существование множества альтернативных вариантов сплайсинга в конечном итоге обеспечивает появление более чем одного белка при экспрессии одного единственного гена. В частности, на гене белка тро–понина человека, содержащем 18 экзонов, за счет альтернативного сплайсинга может образовываться 64 различных продукта! Сейчас подсчитано, что в среднем один ген у человека способен кодировать около 3–х разных белков. Альтернативный сплайсинг зависит от множества внутриклеточных и внешних факторов, в том числе, как показано недавно, даже от вирусного воздействия на клетку.

Рис. 19. Образование молекул мРНК на РНК–предшественнике в результате сплайсинга. Из РНК–предшественника вырезаются фрагменты, синтезированные с интронных последовательностей гена, а участки, синтезированные на экзонах, соединяются друг с другом, что в конечном итоге приводит к формированию зрелой функционально активной мРНК
Экзон–интронное строение генов дало принципиально новую возможность для эволюции генов – комбинировать отдельные элементы при формировании генов. В результате этого в эволюции возникали новые гены, кодирующие разные белки за счет разного сочетания экзонов одного и того же гена. То есть из уже готовых «слов» составляются совершенно разные «предложения». И такой подход, используемый природой, оказался весьма продуктивным.
Следует отметить, что и в зрелой мРНК, которая образуется уже после удаления из первичного транскрипта интронных нуклеотидных последовательностей, также не все участки кодируют белок. Однако, в отличие от интронов, такие последовательности нуклео–тидов расположены в начале и в конце молекулы (см. рис. 18). Их размеры также существенно различаются. Так, в разных мРНК чело

Рис. 20. Схематическое изображение альтернативного сплайсинга. Прямоугольниками разного оттенка обозначены экзоны, черными линиями – интроны. Показаны лишь две из множества возможных мРНК, образующихся на одном гене
века некодирующая область, расположенная в начале молекулы, имеет длину от 2800 до 18 нуклеотидов (в среднем 200 нуклеотидов), а та, которая расположена в конце, составляет от 8500 до 20 нуклеотидов (в среднем 1000 нуклеотидов). Эти некодирующие белок области гена очень важны для нормального функционирования мРНК, ее трансляции, стабильности и локализации в клетке. Как уже говорилось, на одном гене за счет альтернативного сплайсинга может образовываться несколько мРНК. Следует подчеркнуть, что и на одной уже сформировавшейся мРНК могут образовываться разные белки. Это происходит за счет наличия в 50% мРНК человека не одного, а двух АУГ–кодонов, являющихся старт–сигналами для синтеза белка на мРНК. За счет этого, в разных ситуациях клетка может начать синтез белка с разных мест мРНК, в результате чего образуются разные по длине, а порой и по своим свойствам, белки. Опять же пример из лингвистики. Если читать текст не с первой, а со второй буквы, смысл зачастую совершенно меняется (сравните: удочка и дочка).
В связи со всем этим понятно, что классическая формула генетиков «один ген – один белок», предложенная лауреатами Нобелевской премии Джоржем Бидлом и Эдуардом Татумом, оказывается в реальности справедливой лишь для небольшой группы генов, а многие гены (возможно, большинство) кодируют семейства родственных, но существенно отличающихся белков, то есть нередко действует принцип «один ген – много белков». За счет этого в геноме осуществляется принцип экономии генетического материала (компактная запись) и достигается большой набор генных продуктов – белков – на относительно небольшом числе генов.
В таблице 3 приведена общая характеристика белок–кодирующих генов, выявленных в результате секвенирования генома человека.
Таблица 3. Усредненные данные о структуре белок–кодирую–щих генов генома человека

В реальности размеры разных генов, а также их экзонов и интро–нов сильно отличаются. И основной вклад в эту вариабельность вносят интроны. Уже говорилось, что максимальный по размерам ген дистрофина занимает в геноме человека 2,4 млн. п. н. Подавляющая часть занята именно интронами (их средний размер составляет несколько десятков тысяч п. н.). Наибольшую кодирующую последовательность в геноме человека имеет ген по имени титин. Его размер равен около 81000 п. н. Этот ген – чемпион и по числу интронов (178 штук!), и по длине единичного экзона (17106 п. н.).
Важно отметить, что разбивка генов на отдельные экзоны неслучайна. Индивидуальный экзон часто кодирует не просто какую–то часть белковой молекулы, а определенный фрагмент белка, выполняющий в целом белке отдельную функцию. Такой отдельный структурно–функциональный фрагмент, входящий в состав целого белка, называют доменом.
Перекрывание генных текстов
Не все гены кодируют белки
Ген в гене (генная матрешка)
Генные семейства
Новые предложения из фрагментов старых текстов
Сколько же всего генов у человека?
Во всем этом и состоит основное ноу–хау в организации генов человека и других эукариотических генов. «Лоскутное» устройство большинства генов – чрезвычайно важное эволюционное приобретение высших организмов. За счет сплайсинга в РНК может происходить соединение не только между соседними экзонными нуклео–тидными последовательностями, но и между другими, отстоящими порой в гене на значительном расстоянии. Это называют альтернативным сплайсингом (рис. 20). В геноме человека альтернативный сплайсинг характерен для более трети генов. Этому можно привести следующую аналогию: из слова «администрация» за счет удаления отдельных букв и слогов можно образовать множество совершенно разных по смыслу слов: ад, министр, амнистия, астра, нация. Существование множества альтернативных вариантов сплайсинга в конечном итоге обеспечивает появление более чем одного белка при экспрессии одного единственного гена. В частности, на гене белка тро–понина человека, содержащем 18 экзонов, за счет альтернативного сплайсинга может образовываться 64 различных продукта! Сейчас подсчитано, что в среднем один ген у человека способен кодировать около 3–х разных белков. Альтернативный сплайсинг зависит от множества внутриклеточных и внешних факторов, в том числе, как показано недавно, даже от вирусного воздействия на клетку.
Рис. 19. Образование молекул мРНК на РНК–предшественнике в результате сплайсинга. Из РНК–предшественника вырезаются фрагменты, синтезированные с интронных последовательностей гена, а участки, синтезированные на экзонах, соединяются друг с другом, что в конечном итоге приводит к формированию зрелой функционально активной мРНК
Экзон–интронное строение генов дало принципиально новую возможность для эволюции генов – комбинировать отдельные элементы при формировании генов. В результате этого в эволюции возникали новые гены, кодирующие разные белки за счет разного сочетания экзонов одного и того же гена. То есть из уже готовых «слов» составляются совершенно разные «предложения». И такой подход, используемый природой, оказался весьма продуктивным.
Следует отметить, что и в зрелой мРНК, которая образуется уже после удаления из первичного транскрипта интронных нуклеотидных последовательностей, также не все участки кодируют белок. Однако, в отличие от интронов, такие последовательности нуклео–тидов расположены в начале и в конце молекулы (см. рис. 18). Их размеры также существенно различаются. Так, в разных мРНК чело
Рис. 20. Схематическое изображение альтернативного сплайсинга. Прямоугольниками разного оттенка обозначены экзоны, черными линиями – интроны. Показаны лишь две из множества возможных мРНК, образующихся на одном гене
века некодирующая область, расположенная в начале молекулы, имеет длину от 2800 до 18 нуклеотидов (в среднем 200 нуклеотидов), а та, которая расположена в конце, составляет от 8500 до 20 нуклеотидов (в среднем 1000 нуклеотидов). Эти некодирующие белок области гена очень важны для нормального функционирования мРНК, ее трансляции, стабильности и локализации в клетке. Как уже говорилось, на одном гене за счет альтернативного сплайсинга может образовываться несколько мРНК. Следует подчеркнуть, что и на одной уже сформировавшейся мРНК могут образовываться разные белки. Это происходит за счет наличия в 50% мРНК человека не одного, а двух АУГ–кодонов, являющихся старт–сигналами для синтеза белка на мРНК. За счет этого, в разных ситуациях клетка может начать синтез белка с разных мест мРНК, в результате чего образуются разные по длине, а порой и по своим свойствам, белки. Опять же пример из лингвистики. Если читать текст не с первой, а со второй буквы, смысл зачастую совершенно меняется (сравните: удочка и дочка).
В связи со всем этим понятно, что классическая формула генетиков «один ген – один белок», предложенная лауреатами Нобелевской премии Джоржем Бидлом и Эдуардом Татумом, оказывается в реальности справедливой лишь для небольшой группы генов, а многие гены (возможно, большинство) кодируют семейства родственных, но существенно отличающихся белков, то есть нередко действует принцип «один ген – много белков». За счет этого в геноме осуществляется принцип экономии генетического материала (компактная запись) и достигается большой набор генных продуктов – белков – на относительно небольшом числе генов.
В таблице 3 приведена общая характеристика белок–кодирующих генов, выявленных в результате секвенирования генома человека.
Таблица 3. Усредненные данные о структуре белок–кодирую–щих генов генома человека
В реальности размеры разных генов, а также их экзонов и интро–нов сильно отличаются. И основной вклад в эту вариабельность вносят интроны. Уже говорилось, что максимальный по размерам ген дистрофина занимает в геноме человека 2,4 млн. п. н. Подавляющая часть занята именно интронами (их средний размер составляет несколько десятков тысяч п. н.). Наибольшую кодирующую последовательность в геноме человека имеет ген по имени титин. Его размер равен около 81000 п. н. Этот ген – чемпион и по числу интронов (178 штук!), и по длине единичного экзона (17106 п. н.).
Важно отметить, что разбивка генов на отдельные экзоны неслучайна. Индивидуальный экзон часто кодирует не просто какую–то часть белковой молекулы, а определенный фрагмент белка, выполняющий в целом белке отдельную функцию. Такой отдельный структурно–функциональный фрагмент, входящий в состав целого белка, называют доменом.
Перекрывание генных текстов
Принцип компактности записи информации в ДНК проявляется еще в одном. Важный факт, обнаруженный первоначально при анализе последовательностей генов у вирусов, заключается в том, что в одном и том же участке ДНК иногда может быть записана информация о двух совершенно разных РНК (и, соответственно, о совершенно разных белках). Такое случается и в геноме высших организмов, включая человека. Как же это осуществляется? Вспомним, что записанный в ДНК текст явно не разбит на слова (буквы – нук–леотиды – следуют в молекуле ДНК одна за другой). Но слова в этом тексте есть, просто знаки пунктуации и разрывы между словами скрыты от глаза и узнаются клеткой после перекодировки информации из ДНК в РНК. Для большей наглядности приведем широко известный пример такого текста:
НАПОЛЕОНКОСИЛТРАВУПОЛЯКИПЕЛИСОЛОВЬЯМИ.
В зависимости от разбивки этого текста на слова получается два предложения с совершенно разным значением:
НА ПОЛЕ ОН КОСИЛ ТРАВУ ПОЛЯ КИПЕЛИ
СОЛОВЬЯМИ
НАПОЛЕОН КОСИЛ ТРАВУ ПОЛЯКИ ПЕЛИ
СОЛОВЬЯМИ.
Теоретически на двух цепях ДНК с учетом того, что код триплет–ный, можно записать 6 разных текстов: три на одной цепи и три на другой. Это может быть достигнуто за счет сдвига точки начала считывания информации, что называют сдвигом рамки считывания (начало считывания возможно с любого из 3–х разных рядом лежащих нуклеотидов) (см. рис. 21). Такое явление получило название перекрывания генов. Впервые оно было обнаружено у вирусов. И здесь было понятно, зачем это потребовалось. Вирусные геномы обычно очень маленькие. Перекрывание генов диктовалось необходимостью как можно компактнее разместить информацию на отно

Рис. 21. Схематическое изображение перекрывания двух ДНКовых текстов, записанных на одном участке ДНК. Белок А образуется на мРНК, которая транслируется с одних кодо–нов, а белок Б образуется на мРНК, читаемой (транслируемой) со сдвигом в один нуклеотид. В результате кодоны (они разделены пробелами) совершенно разные, и, соответственно, при трансляции мРНК образуются совершенно разные белки сительно коротких молекулах ДНК. Большой геном человека теоретически позволяет обойтись без такого перекрывания. Однако и в геноме человека, хотя и относительно редко, перекрывание генов также имеется.
Каков может быть смысл в сохранении этого феномена в геноме человека? Пока еще не все ясно, но кое–какие моменты уже проясняются. Например, в ряде случаев обнаружили считывание РНК с разных цепей одного участка ДНК. Это приводит к образованию таких РНК, которые в силу комплементарности могут взаимодействовать друг с другом в клетке, образуя в результате двунитевые РНК. А в дву–нитевом виде мРНК не способна к такому важному процессу, как трансляция. Таков реальный механизм специфической регуляции (инактивации) экспрессии генов человека, число которых, по последним оценкам, составляет ~ 1600.
НАПОЛЕОНКОСИЛТРАВУПОЛЯКИПЕЛИСОЛОВЬЯМИ.
В зависимости от разбивки этого текста на слова получается два предложения с совершенно разным значением:
НА ПОЛЕ ОН КОСИЛ ТРАВУ ПОЛЯ КИПЕЛИ
СОЛОВЬЯМИ
НАПОЛЕОН КОСИЛ ТРАВУ ПОЛЯКИ ПЕЛИ
СОЛОВЬЯМИ.
Теоретически на двух цепях ДНК с учетом того, что код триплет–ный, можно записать 6 разных текстов: три на одной цепи и три на другой. Это может быть достигнуто за счет сдвига точки начала считывания информации, что называют сдвигом рамки считывания (начало считывания возможно с любого из 3–х разных рядом лежащих нуклеотидов) (см. рис. 21). Такое явление получило название перекрывания генов. Впервые оно было обнаружено у вирусов. И здесь было понятно, зачем это потребовалось. Вирусные геномы обычно очень маленькие. Перекрывание генов диктовалось необходимостью как можно компактнее разместить информацию на отно
Рис. 21. Схематическое изображение перекрывания двух ДНКовых текстов, записанных на одном участке ДНК. Белок А образуется на мРНК, которая транслируется с одних кодо–нов, а белок Б образуется на мРНК, читаемой (транслируемой) со сдвигом в один нуклеотид. В результате кодоны (они разделены пробелами) совершенно разные, и, соответственно, при трансляции мРНК образуются совершенно разные белки сительно коротких молекулах ДНК. Большой геном человека теоретически позволяет обойтись без такого перекрывания. Однако и в геноме человека, хотя и относительно редко, перекрывание генов также имеется.
Каков может быть смысл в сохранении этого феномена в геноме человека? Пока еще не все ясно, но кое–какие моменты уже проясняются. Например, в ряде случаев обнаружили считывание РНК с разных цепей одного участка ДНК. Это приводит к образованию таких РНК, которые в силу комплементарности могут взаимодействовать друг с другом в клетке, образуя в результате двунитевые РНК. А в дву–нитевом виде мРНК не способна к такому важному процессу, как трансляция. Таков реальный механизм специфической регуляции (инактивации) экспрессии генов человека, число которых, по последним оценкам, составляет ~ 1600.
Не все гены кодируют белки
Прежде всего, следует отметить, что кроме генов, кодирующих белки, в геноме имеются еще гены, на которых синтезируется РНК, которые не являются мРНК (то есть, не кодируют белок), но выполняют ряд самостоятельных важных функций в клетках. В результате получается ситуация, что традиционное привычное определение гена надо расширять, включив в него гены, кодирующие белки, и гены, не кодирующие белки, но кодирующие функционально значимые РНК.
Давно уже известно, что кроме мРНК на ДНК синтезируются разнообразные вспомогательные РНК, которые сами не транслируются (т. е. они не кодируют белки), но участвуют в разных клеточных процессах. В первую очередь это РНК «домашнего хозяйства» – ри–босомные РНК (рРНК), транспортные РНК (тРНК) и др., которые участвуют непосредственно в синтезе белка на мРНК. Эти РНК кодируются участками, которые также называются генами, и составляют основную массу РНК в клетках. рРНК входят в состав рибосом, являясь важным компонентом их структурной организации. У человека присутствует два основных типа рРНК размером около 1900 нуклеотидов и более 5000 нуклеотидов в малой и большой субчастицах соответственно. Еще одни некодирующие белок РНК – тРНК, которые обеспечивают аппарат трансляции, подтаскивая к рибосомам различные аминокислоты, вступающие в реакцию друг с другом. В геноме человека гены, кодирующие рРНК и тРНК, представлены многочисленными копиями. Так, в секвенированной ДНК человека обнаружено около 500 генов, кодирующих тРНК. Генов для рРНК в геноме человека выявлено около 200 копий, которые расположены на 5–ти разных хромосомах (13, 14, 15, 21 и 22).
Изучение свойств РНК привело к тому, что представление об исключительности белков в катализе биохимических реакций пришел конец. Выяснилось, что в природе имеются виды РНК, которые, подобно белкам, обладают высокоспецифической каталитической активностью.
Очень важным стало обнаружение в геноме человека множества других генов, также производящих РНК, но не способных кодировать белок. Постепенно выяснилось, что некоторые из таких не кодирующих белки РНК принимают участие в важнейших процессах, происходящих в клетке: регуляции транскрипции ДНК, сплайсинга и трансляции мРНК, модулировании функций белков и их пространственного распределения в клетке. По этой причине их назвали риборегуляторами. И примеров таких риборегуляторов уже сейчас можно привести немало. Так, установлено, что не кодирующий белок участок гена H19 имеет отношение к ряду процессов, протекающих в клетках, и, в частности, к их злокачественному перерождению. Другой РНК–кодирующий ген контролирует работу белок–ко–дирующего гена HFE, вовлеченного в метаболизм железа и связанного с наследственным заболеванием хемохроматозом. В последнем случае РНК–продукт кодируется тем же самым геном, который он регулирует, но его образование осуществляется на другой нити ДНК. В результате так называемый антисмысловой (комплементарный) РНК–продукт способен взаимодействовать с мРНК, образуя гибриды, неспособные транслироваться в рибосомах с образованием белка. Еще один интересный ген – ген РНК–активатора стероидного рецептора. Он обеспечивает активность стероидных рецепторов за счет образования комплекса с этим белком.
В клетках человека, как и у других организмов, выявлены короткие двунитевые РНК (микроРНК), отдельные из которых, по–видимому, могут участвовать в процессе регуляции экспрессии генов через механизм, названный РНК–интерференцией. Этот механизм впервые был обнаружен в 1998 году у низших организмов. Различные микроРНК в клетках червя C. elegans оказались в состоянии «приглушать» работу строго определенных генов путем воздействия на процесс синтеза кодируемых ими белков. Вполне вероятно, и у человека микроРНК играют подобную роль.
Таким образом, мы видим, что продукты генов, кодирующих только РНК, вмешиваются в различные клеточные процессы, используя при этом совершенно разные механизмы. Гены риборегу–ляторов составляют, по–видимому, заметную часть генома человека. Примерные оценки говорят о величине на порядок больше, чем доля белок–кодирующих нуклеотидных последовательностей. Сам факт существования таких генов, неспособных кодировать белок, но реально проявляющих себя в производстве функционирующих в клетках РНК, ставит большой вопрос перед исследователями генома. И, в первую очередь, что следует после этого считать собственно геном?
Давно уже известно, что кроме мРНК на ДНК синтезируются разнообразные вспомогательные РНК, которые сами не транслируются (т. е. они не кодируют белки), но участвуют в разных клеточных процессах. В первую очередь это РНК «домашнего хозяйства» – ри–босомные РНК (рРНК), транспортные РНК (тРНК) и др., которые участвуют непосредственно в синтезе белка на мРНК. Эти РНК кодируются участками, которые также называются генами, и составляют основную массу РНК в клетках. рРНК входят в состав рибосом, являясь важным компонентом их структурной организации. У человека присутствует два основных типа рРНК размером около 1900 нуклеотидов и более 5000 нуклеотидов в малой и большой субчастицах соответственно. Еще одни некодирующие белок РНК – тРНК, которые обеспечивают аппарат трансляции, подтаскивая к рибосомам различные аминокислоты, вступающие в реакцию друг с другом. В геноме человека гены, кодирующие рРНК и тРНК, представлены многочисленными копиями. Так, в секвенированной ДНК человека обнаружено около 500 генов, кодирующих тРНК. Генов для рРНК в геноме человека выявлено около 200 копий, которые расположены на 5–ти разных хромосомах (13, 14, 15, 21 и 22).
Изучение свойств РНК привело к тому, что представление об исключительности белков в катализе биохимических реакций пришел конец. Выяснилось, что в природе имеются виды РНК, которые, подобно белкам, обладают высокоспецифической каталитической активностью.
Очень важным стало обнаружение в геноме человека множества других генов, также производящих РНК, но не способных кодировать белок. Постепенно выяснилось, что некоторые из таких не кодирующих белки РНК принимают участие в важнейших процессах, происходящих в клетке: регуляции транскрипции ДНК, сплайсинга и трансляции мРНК, модулировании функций белков и их пространственного распределения в клетке. По этой причине их назвали риборегуляторами. И примеров таких риборегуляторов уже сейчас можно привести немало. Так, установлено, что не кодирующий белок участок гена H19 имеет отношение к ряду процессов, протекающих в клетках, и, в частности, к их злокачественному перерождению. Другой РНК–кодирующий ген контролирует работу белок–ко–дирующего гена HFE, вовлеченного в метаболизм железа и связанного с наследственным заболеванием хемохроматозом. В последнем случае РНК–продукт кодируется тем же самым геном, который он регулирует, но его образование осуществляется на другой нити ДНК. В результате так называемый антисмысловой (комплементарный) РНК–продукт способен взаимодействовать с мРНК, образуя гибриды, неспособные транслироваться в рибосомах с образованием белка. Еще один интересный ген – ген РНК–активатора стероидного рецептора. Он обеспечивает активность стероидных рецепторов за счет образования комплекса с этим белком.
В клетках человека, как и у других организмов, выявлены короткие двунитевые РНК (микроРНК), отдельные из которых, по–видимому, могут участвовать в процессе регуляции экспрессии генов через механизм, названный РНК–интерференцией. Этот механизм впервые был обнаружен в 1998 году у низших организмов. Различные микроРНК в клетках червя C. elegans оказались в состоянии «приглушать» работу строго определенных генов путем воздействия на процесс синтеза кодируемых ими белков. Вполне вероятно, и у человека микроРНК играют подобную роль.
Таким образом, мы видим, что продукты генов, кодирующих только РНК, вмешиваются в различные клеточные процессы, используя при этом совершенно разные механизмы. Гены риборегу–ляторов составляют, по–видимому, заметную часть генома человека. Примерные оценки говорят о величине на порядок больше, чем доля белок–кодирующих нуклеотидных последовательностей. Сам факт существования таких генов, неспособных кодировать белок, но реально проявляющих себя в производстве функционирующих в клетках РНК, ставит большой вопрос перед исследователями генома. И, в первую очередь, что следует после этого считать собственно геном?
Ген в гене (генная матрешка)
Изредка обнаруживаются варианты, когда внутри одного гена целиком содержится другой, меньший по размерам ген. Этакая своеобразная «матрешка», построенная из генов. Такая организация генов весьма редка. Так, в хромосоме 22 имеется лишь 2 таких случая. Чаще всего белок–кодирующий ген располагается в интроне другого белок–кодирующего гена. Но встречаются и другие варианты. В качестве примера можно привести ситуацию, имеющую место для ми–тохондриального гена одной из рибосомных РНК. Ген, кодирующий эту рРНК, обеспечивает ею рибосомы митохондрий в качестве структурного компонента (т. е. не кодирует белок). Однако вместе с тем небольшой участок, расположенный внутри этого гена, кодирует короткий белок (полипептид), получивший название гуманин (от англ. human – человек), который принимает участие в процессе программированной клеточной гибели. То есть в РНК–кодирующем гене может содержатся белок–кодирующий ген. Другой вариант—уже упоминавшийся выше ген H19. Здесь, наоборот, ген, кодирующий белок, содержит внутри своей кодирующей части другой более короткий ген, кодирующий только РНК, которая принимает участие в регуляции работы этого гена.
Генные семейства
Как у людей имеются семьи, так и у генов нередко существуют семейства. Семейством генов называют набор из двух или более генов, чьи экзоны родственны между собой, т. е. похожи (хотя и не идентичны) по нуклеотидной последовательности. В геноме человека присутствует около полутора тысяч таких семейств генов. Причем только около сотни из них специфичны для человека и других позвоночных животных, тогда как основная масса генных семейств имеется и у человека, и у червя.
Разные члены одного семейства генов возникали в эволюции из одного гена–предшественника (условно – от одного прапрапрадеда, как и в человеческой популяции). Процесс, в результате которого появлялись «копии», получил название дупликации, т. е. удвоения. Такие дупликации в некоторых случаях происходят не одноразово, а многократно. Сейчас стало ясно, что дупликации играют очень важную роль в эволюции. Подсчитано, что в геноме человека в сумме дуплицировано около 3,6% нуклеотидных последовательностей размером в 1000 и более п. н.
Разные копии одного семейства генов могут располагаться в геноме рядом и следовать друг за другом (это называют тандемной дупликацией). Рассмотрим несколько примеров. Так, гены, кодирующие рибосомную РНК (рРНК) и белки гистоны, расположены в геноме человека в виде кластеров, построенных из одних и тех же последовательностей, следующих друг за другом (рис. 22).

Рис. 22. Строение участка генома человека, состоящего из множества генов, кодирующих рибосомные РНК. Утолщенные линии – транскрибируемые участки
Такие «гроздья» одинаковых генов обычно требуются организму для того, чтобы нарабатывать большие количества определенного продукта. Действительно, рРНК составляет по массе основную часть всей клеточной РНК, входя в состав таких важных «машин» клетки, как рибосомы. Огромная масса гистонов нужна клетке для упаковки гигантских молекул ДНК и плотной укладки их в хромосомы. Во всех таких случаях говорят о большой «дозе» гена в геноме, которая в конечном итоге и обеспечивает клетку большим количеством продукта.
Однако в большинстве других случаев возникающие в результате дупликации «копии» генов постепенно претерпевают изменения (мутации), что приводит к возникновению определенных различий между ними, хотя формальное родство при этом сохраняется. Такой процесс был назван дивергенцией, т. е. расхождением. Дивергенцию обычно выражает в процентах различий между двумя родственными последовательностями нуклеотидов в ДНК или последовательностями аминокислот в белках.
Однако даже после того, как разные «копии» генов одного семейства становятся не совсем похожими друг на друга, они чаще всего выполняют сходную функцию. Они остаются очень дружной семейкой! Но экспрессируются разные члены одного такого семейства обычно в разное время или в разных типах клеток. Так, в геноме человека обнаружено тридцать генов, кодирующих родственные факторы роста клеток кожи (фибробластов). А вот у низших организмов число таких генов существенно меньше (у дрозофилы и червя их всего 2). Другой пример – гены, кодирующие кератины – белки наружного слоя кожи и ее производных (волосы, ногти). Их у человека 111 штук! Но и это не предел. Геном человека, как ни удивительно, имеет около 1 000 копий генов–рецепторов обоняния.
Дивергировавшие в ходе эволюции человека гены семейства иногда оказываются разбросанными в разных местах на одной или даже разных хромосомах. При этом ген может быть удвоен целиком, а может происходить удвоение небольшого набора его экзонов или даже всего лишь одного из его экзонов.
Как и в семье не без урода, так и в семействах генов встречаются иногда такие члены, которые не утруждают себя никакой работой. Нарушение их способности функционировать в клетке обычно связано с накоплением опечаток в их ДНКовом тексте. Такие гены–тунеядцы получили название «псевдогенов». Перед их названием ставят греческую букву ψ. Не совсем ясно, зачем геному нужны такие гены, почему он сохранил их в эволюции, не избавился от них. Но факт остается фактом. По имеющимся оценкам, в геноме человека имеется около 20000 таких, подобных вымершим реликтам, псевдогенов. В частности, в огромной семействе генов–рецепторов обоняния, состоящем из примерно 1000 генов, около 60% являются псевдогенами. Считается, что массивная потеря функциональных генов произошла за последние 10 млрд. лет, и связано это со снижением роли обоняния у человека по сравнению с другими позвоночными организмами. Здесь, по–видимому, действует принцип отмирания из–за неупотребления. Установлено, что существует, по крайней мере, два механизма, приводящих к образованию псевдогенов. Один из них заключается в удвоении уже существующего гена (дупликация), а потом дополнительная копия накапливает мутации, которые препятствуют его работе. Другой механизм связан с процессом сплайсинга и обратной транскрипции. На сформировавшейся мРНК синтезируется ее ДНК–копия, которая в дальнейшем встраивается в геном. Такие псевдогены не содержат ни интроны, ни промоторные участки. Кроме того, в них со временем накапливаются многочисленные мутации, не позволяющие этим генам обеспечивать синтез нормального белка.
Все вышесказанное можно проиллюстрировать на примере семейства генов, кодирующих белки крови человека (рис. 23). У человека имеется два семейства глобиновых генов (альфа и бета). Эти семейства расположены на разных хромосомах. Каждое семейство состоит из нескольких членов, имеются здесь и псевдогены. Нормальные гены глобиновых семейств экспрессируются в клетках крови на разных стадиях развития человека. Одни у эмбрионов (ξ, ε), другие у плода ( Gγ, Аγ, a1), а третьи – у взрослого организма (α2, δ, β). Функция у всех глобиновых генов одинакова – синтез глобинов, участвующих в переносе кислорода клетками крови. И место их работы одно – клетки крови эритроциты. Отличаются они лишь тем, что включаются в разное время, в результате чего выполняют одну и ту же роль, но на разных стадиях развития организма.

Рис. 23. Строение двух локусов генома, состоящих из кластеров родственных глобиновых генов человека. Экспрессирую–щиеся гены указаны стрелками (направление транскрипции), молчащие гены (псевдогены) – квадратиками
Организация кластеров глобиновых генов у человека совершенно одинакова с организацией этих генов у гориллы и павиана. Это указывает на то, что такая организация полностью сформировалась еще от 20 до 40 млн. лет назад и с тех пор не изменялась. Анализ мутаций в межгенных областях позволил сделать вывод об эволюции не отдельных генов, собранных в такой кластер, а кластера как единого целого супергена.
По–иному ведут себя многочисленные гены семейства актинов, которые кодируют сократительные белки клетки. Эти гены экспрес–сируются почти все время, и их экспрессия происходит как в мышечных клетках, так и во многих других немышечных. При этом функция актинов везде одинакова.
Разные члены одного семейства генов возникали в эволюции из одного гена–предшественника (условно – от одного прапрапрадеда, как и в человеческой популяции). Процесс, в результате которого появлялись «копии», получил название дупликации, т. е. удвоения. Такие дупликации в некоторых случаях происходят не одноразово, а многократно. Сейчас стало ясно, что дупликации играют очень важную роль в эволюции. Подсчитано, что в геноме человека в сумме дуплицировано около 3,6% нуклеотидных последовательностей размером в 1000 и более п. н.
Разные копии одного семейства генов могут располагаться в геноме рядом и следовать друг за другом (это называют тандемной дупликацией). Рассмотрим несколько примеров. Так, гены, кодирующие рибосомную РНК (рРНК) и белки гистоны, расположены в геноме человека в виде кластеров, построенных из одних и тех же последовательностей, следующих друг за другом (рис. 22).
Рис. 22. Строение участка генома человека, состоящего из множества генов, кодирующих рибосомные РНК. Утолщенные линии – транскрибируемые участки
Такие «гроздья» одинаковых генов обычно требуются организму для того, чтобы нарабатывать большие количества определенного продукта. Действительно, рРНК составляет по массе основную часть всей клеточной РНК, входя в состав таких важных «машин» клетки, как рибосомы. Огромная масса гистонов нужна клетке для упаковки гигантских молекул ДНК и плотной укладки их в хромосомы. Во всех таких случаях говорят о большой «дозе» гена в геноме, которая в конечном итоге и обеспечивает клетку большим количеством продукта.
Однако в большинстве других случаев возникающие в результате дупликации «копии» генов постепенно претерпевают изменения (мутации), что приводит к возникновению определенных различий между ними, хотя формальное родство при этом сохраняется. Такой процесс был назван дивергенцией, т. е. расхождением. Дивергенцию обычно выражает в процентах различий между двумя родственными последовательностями нуклеотидов в ДНК или последовательностями аминокислот в белках.
Однако даже после того, как разные «копии» генов одного семейства становятся не совсем похожими друг на друга, они чаще всего выполняют сходную функцию. Они остаются очень дружной семейкой! Но экспрессируются разные члены одного такого семейства обычно в разное время или в разных типах клеток. Так, в геноме человека обнаружено тридцать генов, кодирующих родственные факторы роста клеток кожи (фибробластов). А вот у низших организмов число таких генов существенно меньше (у дрозофилы и червя их всего 2). Другой пример – гены, кодирующие кератины – белки наружного слоя кожи и ее производных (волосы, ногти). Их у человека 111 штук! Но и это не предел. Геном человека, как ни удивительно, имеет около 1 000 копий генов–рецепторов обоняния.
Дивергировавшие в ходе эволюции человека гены семейства иногда оказываются разбросанными в разных местах на одной или даже разных хромосомах. При этом ген может быть удвоен целиком, а может происходить удвоение небольшого набора его экзонов или даже всего лишь одного из его экзонов.
Как и в семье не без урода, так и в семействах генов встречаются иногда такие члены, которые не утруждают себя никакой работой. Нарушение их способности функционировать в клетке обычно связано с накоплением опечаток в их ДНКовом тексте. Такие гены–тунеядцы получили название «псевдогенов». Перед их названием ставят греческую букву ψ. Не совсем ясно, зачем геному нужны такие гены, почему он сохранил их в эволюции, не избавился от них. Но факт остается фактом. По имеющимся оценкам, в геноме человека имеется около 20000 таких, подобных вымершим реликтам, псевдогенов. В частности, в огромной семействе генов–рецепторов обоняния, состоящем из примерно 1000 генов, около 60% являются псевдогенами. Считается, что массивная потеря функциональных генов произошла за последние 10 млрд. лет, и связано это со снижением роли обоняния у человека по сравнению с другими позвоночными организмами. Здесь, по–видимому, действует принцип отмирания из–за неупотребления. Установлено, что существует, по крайней мере, два механизма, приводящих к образованию псевдогенов. Один из них заключается в удвоении уже существующего гена (дупликация), а потом дополнительная копия накапливает мутации, которые препятствуют его работе. Другой механизм связан с процессом сплайсинга и обратной транскрипции. На сформировавшейся мРНК синтезируется ее ДНК–копия, которая в дальнейшем встраивается в геном. Такие псевдогены не содержат ни интроны, ни промоторные участки. Кроме того, в них со временем накапливаются многочисленные мутации, не позволяющие этим генам обеспечивать синтез нормального белка.
Все вышесказанное можно проиллюстрировать на примере семейства генов, кодирующих белки крови человека (рис. 23). У человека имеется два семейства глобиновых генов (альфа и бета). Эти семейства расположены на разных хромосомах. Каждое семейство состоит из нескольких членов, имеются здесь и псевдогены. Нормальные гены глобиновых семейств экспрессируются в клетках крови на разных стадиях развития человека. Одни у эмбрионов (ξ, ε), другие у плода ( Gγ, Аγ, a1), а третьи – у взрослого организма (α2, δ, β). Функция у всех глобиновых генов одинакова – синтез глобинов, участвующих в переносе кислорода клетками крови. И место их работы одно – клетки крови эритроциты. Отличаются они лишь тем, что включаются в разное время, в результате чего выполняют одну и ту же роль, но на разных стадиях развития организма.
Рис. 23. Строение двух локусов генома, состоящих из кластеров родственных глобиновых генов человека. Экспрессирую–щиеся гены указаны стрелками (направление транскрипции), молчащие гены (псевдогены) – квадратиками
Организация кластеров глобиновых генов у человека совершенно одинакова с организацией этих генов у гориллы и павиана. Это указывает на то, что такая организация полностью сформировалась еще от 20 до 40 млн. лет назад и с тех пор не изменялась. Анализ мутаций в межгенных областях позволил сделать вывод об эволюции не отдельных генов, собранных в такой кластер, а кластера как единого целого супергена.
По–иному ведут себя многочисленные гены семейства актинов, которые кодируют сократительные белки клетки. Эти гены экспрес–сируются почти все время, и их экспрессия происходит как в мышечных клетках, так и во многих других немышечных. При этом функция актинов везде одинакова.
Новые предложения из фрагментов старых текстов
Экзон–интронная организация генов способствует еще одному механизму возникновения новых генов – созданию новых ДНКовых текстов из фрагментов старых. Это осуществляется в результате «тасования» уже предсуществующих экзонов, что может приводить к появлению нового их сочетания и новых белков с новыми функциями. Чаще всего это происходит за счет процесса присоединения новых экзонов к старым. Примером этому может служить случай, представленный на рис. 24.
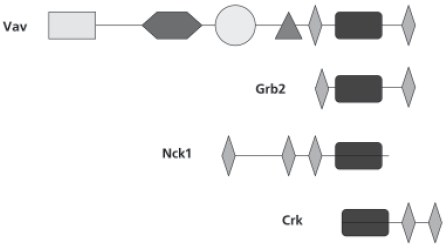
Рис. 24. Пример формирования новых генов за счет различного сочетания уже имеющихся экзонов. Изображены четыре разных белка, состоящих из сходных блоков–доменов (обозначены разными символами), которые кодируются разными экзонами
В процессе эволюции из общего предшественника образовались гены дрозофилы trx и ген человека alr. Ген человека hrx возник в результате присоединения к гену trx четырех новых экзонов. Это приводит к появлению в белке, кодируемом этим геном, четырех новых функциональных доменов, что существенно меняет его свойства по сравнению с предшественником. Такая ситуация вообще–то характерна для всех высших организмов, но у человека она встречается в 2—5 раз чаще, чем, например, у дрозофилы или червя.
После получения всех этих новых многочисленных сведений об особенностях организации и эволюции генов у человека (сходным образом обстоит дело и у других высших организмов) стало ясно, что дать строгое определение того, что же такое собой представляет ген, довольно сложно. Но даже если бы такое определение сегодня имелось, его, как и всякое другое, не следовало бы считать незыблемым. Не зря говорил Спиноза: «Всякое определение есть ограничение». Однако в целом базовое представление о гене осталось в силе. Ген – это физическая (определенный участок ДНК) и функциональная (кодирует белок или РНК) единица наследственности.
Рис. 24. Пример формирования новых генов за счет различного сочетания уже имеющихся экзонов. Изображены четыре разных белка, состоящих из сходных блоков–доменов (обозначены разными символами), которые кодируются разными экзонами
В процессе эволюции из общего предшественника образовались гены дрозофилы trx и ген человека alr. Ген человека hrx возник в результате присоединения к гену trx четырех новых экзонов. Это приводит к появлению в белке, кодируемом этим геном, четырех новых функциональных доменов, что существенно меняет его свойства по сравнению с предшественником. Такая ситуация вообще–то характерна для всех высших организмов, но у человека она встречается в 2—5 раз чаще, чем, например, у дрозофилы или червя.
После получения всех этих новых многочисленных сведений об особенностях организации и эволюции генов у человека (сходным образом обстоит дело и у других высших организмов) стало ясно, что дать строгое определение того, что же такое собой представляет ген, довольно сложно. Но даже если бы такое определение сегодня имелось, его, как и всякое другое, не следовало бы считать незыблемым. Не зря говорил Спиноза: «Всякое определение есть ограничение». Однако в целом базовое представление о гене осталось в силе. Ген – это физическая (определенный участок ДНК) и функциональная (кодирует белок или РНК) единица наследственности.
Сколько же всего генов у человека?
Это наиболее интересный вопрос, ради которого собственно и затевалось полное секвенирование генома человека. После получения основной информации о структуре генома человека в первую очередь были произведены различные анализы по поиску генов и определению их числа. Однако задача оказалась не простой. Это может показаться странным для читателя, но однозначного ответа на поставленный вопрос до сих пор нет.
Сколько же всего генов в ДНК человека? Еще несколько лет назад полагали, что их около 100 тыс., затем решили, что не более 80 тыс. В конце 1998 г. пришли к выводу, что в геноме человека не более 50—60 тыс. генов и на их долю приходится около 3% общей длины ДНК.
Последние подсчеты общего числа генов в геноме человека проводили несколько международных команд ученых. Уже упоминавшаяся компания «Celera» провела собственные исследования, результаты которого изложены в журнале «Science» в 2001 году. По ее оценкам общее число генов в геноме человека составляет от 26383 до 39114. Средний размер гена оценивается равным примерно 3000 п. н. Если принять, что число генов у человека порядка 30 тысяч генов и на каждый ген приходится примерно 3 тыс. п. н., то нетрудно подсчитать, что в кодировании белков принимает участие менее 1,5% хромосомной ДНК. Таким образом, генетические инструкции по формированию человеческой личности занимают меньше 3 сантиметров на двухметровой молекуле ДНК. Удивляет и малое количество генов, несущих эти инструкции, – их всего в пять раз больше, чем, например, у такого на наш взгляд совершенно примитивного организма, как муха дрозофила.
Сколько же всего генов в ДНК человека? Еще несколько лет назад полагали, что их около 100 тыс., затем решили, что не более 80 тыс. В конце 1998 г. пришли к выводу, что в геноме человека не более 50—60 тыс. генов и на их долю приходится около 3% общей длины ДНК.
Последние подсчеты общего числа генов в геноме человека проводили несколько международных команд ученых. Уже упоминавшаяся компания «Celera» провела собственные исследования, результаты которого изложены в журнале «Science» в 2001 году. По ее оценкам общее число генов в геноме человека составляет от 26383 до 39114. Средний размер гена оценивается равным примерно 3000 п. н. Если принять, что число генов у человека порядка 30 тысяч генов и на каждый ген приходится примерно 3 тыс. п. н., то нетрудно подсчитать, что в кодировании белков принимает участие менее 1,5% хромосомной ДНК. Таким образом, генетические инструкции по формированию человеческой личности занимают меньше 3 сантиметров на двухметровой молекуле ДНК. Удивляет и малое количество генов, несущих эти инструкции, – их всего в пять раз больше, чем, например, у такого на наш взгляд совершенно примитивного организма, как муха дрозофила.