Температура
в астрофизике).">цветовая температура,т. е. температура в градусах Кельвина абсолютно чёрного тела, при которой оно имеет данную цветность.
Лит.:Гуревич М. М., Цвет и его измерение, М. — Л., 1950; Кривошеев М. И., Кустарев А. К., Световые измерения в телевидении, М., 1973; Нюберг Н. Д., Измерение цвета и цветовые стандарты, М., 1933; Wright W. D., The measurement of colour, 3 ed., L., 1964; Wyszecky G., Stiles W. S., Color science, N. Y., 1967.
А. К. Кустарёв.
 Рис. 3. График цветностей для x, y системы XYZ и цветовой треугольник системы RGB (R — красный, G — зелёный, B — синий).
Рис. 3. График цветностей для x, y системы XYZ и цветовой треугольник системы RGB (R — красный, G — зелёный, B — синий).
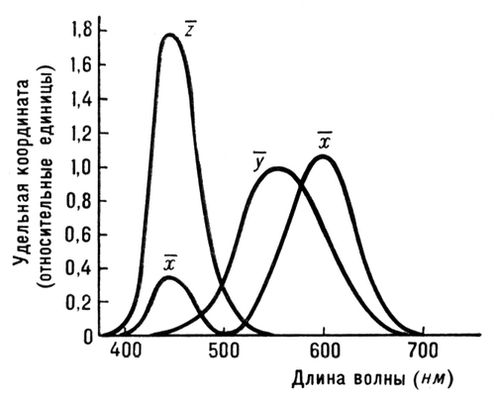 Рис. 2. Кривые сложения для ЦКС МКО XYZ.
Рис. 2. Кривые сложения для ЦКС МКО XYZ.
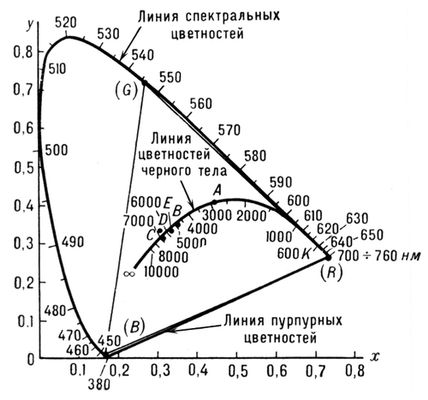 Рис. 1. Кривые сложения для ЦКС МКО RGB.
Рис. 1. Кривые сложения для ЦКС МКО RGB.
спектральной чувствительности
элементарных слоев многослойного
цветофотографического материала.Число диапазонов равно числу элементарных слоев (как правило, их три). Ц. — первая стадия цветного фотографического процесса, обеспечивающая получение комплекта одноцветных изображений на многослойном материале. В
цветной печати
эти изображения получают на отдельных светочувствительных слоях и называются цветоделёнными негативами. При идеальном Ц. вся спектральная область излучения объекта разделяется на граничащие друг с другом, но не перекрывающиеся диапазоны, которые по своим спектральным характеристикам (СХ) точно соответствуют СХ элементарных слоев. Однако практически Ц., осуществляемое всегда с помощью каких-либо селективно поглощающих сред (красителей в элементарных слоях фотоматериала,
светофильтров
в виде цветных стекол или окрашенных плёнок и т.д.), неизбежно несколько отличается от идеального, сопровождаясь, например, перекрыванием зон пропускания, неполным совпадением СХ. Поэтому в окончательном изображении возникают искажения цветовоспроизведения, причем эти искажения в принципе не могут быть устранены на последующих стадиях цветофотографического процесса. См. также
Цветная фотография.
Лит.:Гуревич М. М., Цвет и его измерение, М. — Л., 1950; Кривошеев М. И., Кустарев А. К., Световые измерения в телевидении, М., 1973; Нюберг Н. Д., Измерение цвета и цветовые стандарты, М., 1933; Wright W. D., The measurement of colour, 3 ed., L., 1964; Wyszecky G., Stiles W. S., Color science, N. Y., 1967.
А. К. Кустарёв.