Страница:
Еще более убедительные доказательства способности животных представлять упорядоченность (ординальность) в ряду чисел были получены даже не на шимпанзе, а на низших обезьянах (Brannon, Terrace 1998).
Способность к символизации у птиц (на примере врановых)
Операции логического вывода
Транзитивное заключение
Выявление аналогий
Сравнительная характеристика мышления человекообразных обезьян и перспективы поиска биологических истоков речи человека
Эволюция взглядов на интеллект антропоидов
Опыт проводили следующим образом: на чувствительном к прикосновениям мониторе макакам предъявляли по четыре множества, содержащих от 1 до 4 элементов (рис. 8, см. вклейку). Обезьяны должны были по очереди прикоснуться к каждому из этих множеств в возрастающем порядке. По завершении обучения, когда обезьяны усвоили порядок выбора данных четырех множеств, им предъявляли 150 новых наборов множеств в том же диапазоне от 1 до 4, причем каждый показывали лишь один раз. После того как обезьяны успешно справились с этой задачей, им предъявили множества, содержащие от 1 до 9 элементов. Обезьяны успешно ранжировали новые множества именно по числу элементов в них, используя для этого правило выбора по возрастанию, которому они ранее обучились на множествах, содержащих от 1 до 4 элементов (Brannon, Terrace 1998).Таким образом, приматы способны распознавать и обобщать признак «число элементов», устанавливать соответствие между этим отвлеченным признаком и ранее нейтральными для них стимулами – арабскими цифрами. Оперируя цифрами как символами, они способны ранжировать множества и упорядочивать их по признаку «число», а также совершать число действий, соответствующее цифре. Наконец, они способны к выполнению операций, изоморфных сложению и вычитанию, но этот вопрос, несомненно, требует более точных исследований.
Способность к символизации у птиц (на примере врановых)
Оценка способности к символизации у животных, не относящихся к приматам, представляет самостоятельный интерес, позволяя судить о том, на каких этапах филогенетического развития появляется эта когнитивная способность. Мы исследовали ее у врановых птиц, и выбор этого объекта был не случаен.
Изучение высших представителей класса птиц (попугаев и врановых) показало, что даже самые высокие степени обобщения – образование довербальных понятий – нельзя считать прерогативой приматов (Зорина и др. 2001; Koehler 1956; Mackintosh 1988; Pepperberg 1999/2002). Развитая функция обобщения и абстрагирования позволяет им оперировать рядом отвлеченных понятий, включая довербальное понятие о «числе» (Зорина, Смирнова 1995; Смирнова и др. 1998, 2002; Smirnova et al. 2000; Pepperberg 1991, 1996; Pepperberg, Gordon 2005). Поэтому появилось основание проверить, способны ли вороны к символизации, т. е. могут ли они установить эквивалентность цифр от 1 до 4 и соответствующих им множеств. Для этого был разработан особый методический подход (Зорина и др. 2001). В отличие от предыдущих исследований (Matsuzawa et al. 1986; Matsuzawa 1985; Murofushi 1997), у ворон не вырабатывали ассоциативных связей «цифра и соответствующее ей множество», но создавали условия для того, чтобы птицы смогли самостоятельно выявить эту связь на основе информации, полученной в специальных «демонстрационных» сериях.
Основываясь на результатах целого ряда поведенческих тестов, Л. В. Крушинский (1986) высказал предположение о существовании параллелизма в эволюции высших когнитивных функций птиц и млекопитающих – позвоночных с разными типами структурно-функциональной организации мозга. Мы привели эти данные, чтобы показать, что обобщение, абстрагирование, формирование довербальных понятий и способность к символизации – достаточно универсальные когнитивные операции, в сходной степени присущие высшим представителям разных классов позвоночных.
С тех пор всё новые и новые данные подтверждают, что, несмотря на принадлежность к различным ветвям эволюции и кардинальные различия в структуре мозга, способность к разным видам элементарного мышления, в том числе к обобщению и абстрагированию, у представителей этих классов характеризуется сходными градациями. Это свидетельствует о том, что предыстория человеческого мышления восходит к достаточно древним этапам филогенеза, общим для предков этих классов.
Изучение высших представителей класса птиц (попугаев и врановых) показало, что даже самые высокие степени обобщения – образование довербальных понятий – нельзя считать прерогативой приматов (Зорина и др. 2001; Koehler 1956; Mackintosh 1988; Pepperberg 1999/2002). Развитая функция обобщения и абстрагирования позволяет им оперировать рядом отвлеченных понятий, включая довербальное понятие о «числе» (Зорина, Смирнова 1995; Смирнова и др. 1998, 2002; Smirnova et al. 2000; Pepperberg 1991, 1996; Pepperberg, Gordon 2005). Поэтому появилось основание проверить, способны ли вороны к символизации, т. е. могут ли они установить эквивалентность цифр от 1 до 4 и соответствующих им множеств. Для этого был разработан особый методический подход (Зорина и др. 2001). В отличие от предыдущих исследований (Matsuzawa et al. 1986; Matsuzawa 1985; Murofushi 1997), у ворон не вырабатывали ассоциативных связей «цифра и соответствующее ей множество», но создавали условия для того, чтобы птицы смогли самостоятельно выявить эту связь на основе информации, полученной в специальных «демонстрационных» сериях.
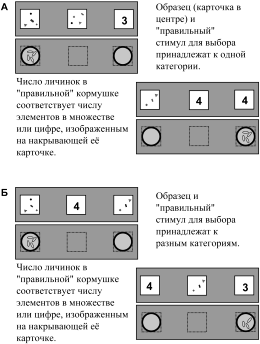
В «демонстрационной» серии впервые за весь период обучения птицы одновременно видели и цифры и множества – например, если образцом была цифра 4, то для выбора предъявляли какое-нибудь множество и цифру 4, а если образцом было множество из четырех элементов, то на карточках для выбора изображали какую-нибудь цифру и множество из четырех элементов (рис. 9А). Кроме того, вороны получали информацию о «цене» каждого стимула. В случае правильного выбора они получали дифференцированное подкрепление: находили то число личинок, которое соответствовало цифре или множеству на выбранной ими карточке. Например, и под карточкой с множеством из четырех элементов, и под цифрой 4 ворона находила 4 личинки. Для успешного решения задачи в демонстрационных сериях воронам достаточно было использовать ранее усвоенное правило выбора по соответствию с образцом – «выбирай изображение, похожее на образец».
В тесте мы выясняли, могут ли птицы установить эквивалентность цифр и соответствующих им множеств без направленного формирования такой связи. В этом опыте образец впервые принадлежал к одной категории, а обе карточки для выбора – к другой (рис. 9Б). Например, если образцом была цифра 4, то для выбора предъявляли два множества, но только одно из них состояло из четырех элементов. Если образцом было множество из четырех элементов, то для выбора предъявляли две цифры, одна из которых была четверкой. Таким образом, между образцом и карточками для выбора не было внешнего соответствия. Для успешного решения такой задачи воронам нужно было не только использовать ранее усвоенное правило выбора по образцу, но и произвести дополнительные операции, мысленно сопоставить еще какую-то ранее полученную информацию об образце и стимулах для выбора. Такой информацией было число единиц подкрепления, связанное с каждым из стимулов во время демонстрационных серий,
Птицы с первых же проб решали эту задачу правильно: в достоверном большинстве случаев они выбирали цифру, соответствующую изображенному на образце множеству и наоборот. Следовательно, вороны способны без специального обучения, за счет мысленного сопоставления ранее полученной информации, установить эквивалентность множеств и исходно индифферентных для них знаков (цифр от 1 до 4).
Рис. 9. Исследование способности к символизации у ворон (по Смирнова и др. 2002). А – демонстрационная серия; Б – тест (пояснения в тексте)
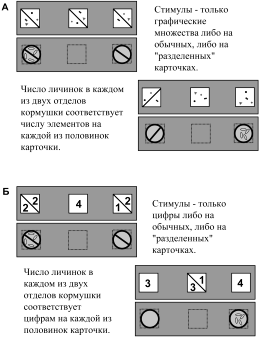
Мы предполагаем, что в данном случае птица принимала решение с помощью операции логического вывода, которую называют транзитивным заключением (см. ниже, раздел «Операции логического вывода»). Поскольку ранее каждому графическому множеству и каждой цифре соответствовало определенное число личинок, то выбирать нужно тот стимул, за который ранее давали столько же личинок, сколько и за образец (если А = В и В = С, то А = С). То есть, на основе двух посылок, полученных ассоциативным или условно-рефлекторным путем, животное может сделать вывод о наличии третьей связи.В следующем эксперименте мы выясняли, могут ли птицы оперировать усвоенной информацией – выполнять с цифрами комбинаторную операцию, аналогичную арифметическому сложению. «Слагаемые» (цифры или множества) были изображены на карточках, разделенных по диагонали чертой, так же как и соответствующие им кормушки были разделены вертикальной перегородкой на две равные части (рис. 10А). В «демонстрационной» серии использовали только множества либо на обычных, либо на «разделенных» карточках и демонстрировали соответствие числа элементов на обычных либо на «разделенных» карточках числу личинок в обычных или в «разделенных» кормушках.
В отличие от «демонстрационной» серии, в тесте на «сложение» мы использовали только цифры. Если в качестве образца предъявляли отдельную цифру, то для выбора – две «разделенные» карточки с цифрой в каждой половине, сумма которых на одной из них соответствовала цифре на образце (рис. 10Б). Если в качестве образца использовали «разделенную» карточку с парой цифр, то для выбора предлагали отдельные цифры.
Птицы успешно справились с этой задачей. К началу серии они уже знали, что каждому конкретному графическому множеству и каждой цифре соответствует определенное число личинок, и на этом основании во время теста они (экстренно) определяли, что определенные цифры и графические множества соответствуют друг другу. Затем, в ходе демонстрационной серии вороны получали дополнительную информацию о том, что под карточкой с «разделенным» множеством находится соответствующим образом «разделенное» число личинок. Для правильного выполнения теста на «сложение» им нужно было сделать мысленное заключение об эквивалентности друг другу отдельных цифр и соответствующих комбинаций двух цифр.
Рис. 10. Исследование способности к оперированию символами («сложению» цифр) у ворон (по Смирнова и др. 2002). А – демонстрационная серия; Б – тест
Оказалось, что вороны способны сохранять информацию о числовых признаках стимулов не только в форме образных представлений, но и в некой отвлеченной и обобщенной форме, которую они смогли связать с ранее нейтральными для них знаками – цифрами. Следовательно, не только у высших приматов, но и у некоторых птиц довербальное мышление достигло в своем развитии того промежуточного этапа, который, по мнению Орбели (1949), обеспечивает возможность использования символов вместо реальных объектов и явлений и который в эволюции предшествовал формированию второй сигнальной системы.Основываясь на результатах целого ряда поведенческих тестов, Л. В. Крушинский (1986) высказал предположение о существовании параллелизма в эволюции высших когнитивных функций птиц и млекопитающих – позвоночных с разными типами структурно-функциональной организации мозга. Мы привели эти данные, чтобы показать, что обобщение, абстрагирование, формирование довербальных понятий и способность к символизации – достаточно универсальные когнитивные операции, в сходной степени присущие высшим представителям разных классов позвоночных.
С тех пор всё новые и новые данные подтверждают, что, несмотря на принадлежность к различным ветвям эволюции и кардинальные различия в структуре мозга, способность к разным видам элементарного мышления, в том числе к обобщению и абстрагированию, у представителей этих классов характеризуется сходными градациями. Это свидетельствует о том, что предыстория человеческого мышления восходит к достаточно древним этапам филогенеза, общим для предков этих классов.
Операции логического вывода
Говоря о мышлении животных в контексте обучения обезьян языкам-посредникам, нельзя не упомянуть о способности животных к совершению двух операций логического вывода (inferential reasoning), когда при получении новой информации ее усвоение происходит на основе уже имеющейся, путем мысленного сопоставления ранее полученных сведений. Об этих операциях необходимо упомянуть по двум причинам. Во-первых, само начало их изучения связано с именем Д. Примэка – одного из первопроходцев в области изучения зачатков речи у обезьян (Gillan et al. 1981; Premack 1983; Premack & Premack 1972, 2003). Во-вторых, одна из операций – транзитивное заключение – тесно связана с процессом символизации.
Транзитивное заключение
Упомянутое выше транзитивное заключение составляет одну из базовых операций, присущих дедуктивному мышлению человека. Определение транзитивного отношения пришло в психологию из формальной и математической логики. Отношение r называют транзитивным, если оно, попарно связывая стимулы B и C, C и D, с необходимостью связывает также стимулы B и D. Стимулы, связанные такими отношениями, образуют транзитивный ряд: B r C r D. Таким образом, если субъект способен из двух предпосылок B r C и C r D сделать вывод, что B r D, то говорят, что он способен к транзитивному заключению. Примерами транзитивных отношений могут служить такие, как «больше, чем» (например, если B > C и C > D, то B > D), «дальше», «ярче» и т. д. Наряду с отношением транзитивных неравенств существует и возможность транзитивного равенства: если A=B и B=C, то A=С. Именно эта операция, как мы увидим, вовлечена в процесс символизации у птиц – в превращение нейтральных для них стимулов (цифр) в символы-числительные, которыми они далее оперируют вместо соответствующих множеств.
Способность к транзитивному заключению, по мнению многих известных приматологов (Altmann 1962; de Waal 1977; Kummer 1982; Premack 1986 и др.), играет важную роль в организации социального поведения, в частности, в установлении иерархии в сообществах. Например, если обезьяна видит, что субъект B подчиняется субъекту A, а она сама подчиняется субъекту B, то без дополнительного выяснения отношений приходит к выводу, что ей надо уступать субъекту A.
Способность к решению теста на транзитивное заключение у шимпанзе описана в лаборатории Примэка в 1981 году (Gillan 1981). Впоследствии она была обнаружена у многих животных, в том числе и у птиц, включая голубей и ворон (Зорина и др. 1995; Siemann et al. 1996). Интересно, что механизмы осуществления этой операции до сих пор неясны. Ряд исследователей считают, что это форма когнитивной деятельности, которая осуществляется за счет мысленного сравнения положения стимулов в транзитивном ряду (Зорина и др. 1995; Davis 1993; Boysen et al. 1993; Rapp et al. 1996; Lazareva et al. 2004 и др.). Другие авторы склоняются к тому, что у голубей механизм решения имеет чисто ассоциативную природу и выбор в тесте осуществляется за счет различий в вероятности подкрепления стимулов во время предшествующего обучения (Wynne 1995; Siemann, Delius 1998).
По-видимому, у животных разных видов и в разных вариантах опытов могут быть задействованы разные механизмы. Во всяком случае, проведенные нами эксперименты (Lazareva et al. 2004) позволили показать, что у врановых птиц транзитивное заключение осуществляется именно на основе мысленного сопоставления независимо полученной информации об относительном положении стимулов в транзитивном ряду.
Способность к транзитивному заключению, по мнению многих известных приматологов (Altmann 1962; de Waal 1977; Kummer 1982; Premack 1986 и др.), играет важную роль в организации социального поведения, в частности, в установлении иерархии в сообществах. Например, если обезьяна видит, что субъект B подчиняется субъекту A, а она сама подчиняется субъекту B, то без дополнительного выяснения отношений приходит к выводу, что ей надо уступать субъекту A.
Способность к решению теста на транзитивное заключение у шимпанзе описана в лаборатории Примэка в 1981 году (Gillan 1981). Впоследствии она была обнаружена у многих животных, в том числе и у птиц, включая голубей и ворон (Зорина и др. 1995; Siemann et al. 1996). Интересно, что механизмы осуществления этой операции до сих пор неясны. Ряд исследователей считают, что это форма когнитивной деятельности, которая осуществляется за счет мысленного сравнения положения стимулов в транзитивном ряду (Зорина и др. 1995; Davis 1993; Boysen et al. 1993; Rapp et al. 1996; Lazareva et al. 2004 и др.). Другие авторы склоняются к тому, что у голубей механизм решения имеет чисто ассоциативную природу и выбор в тесте осуществляется за счет различий в вероятности подкрепления стимулов во время предшествующего обучения (Wynne 1995; Siemann, Delius 1998).
По-видимому, у животных разных видов и в разных вариантах опытов могут быть задействованы разные механизмы. Во всяком случае, проведенные нами эксперименты (Lazareva et al. 2004) позволили показать, что у врановых птиц транзитивное заключение осуществляется именно на основе мысленного сопоставления независимо полученной информации об относительном положении стимулов в транзитивном ряду.
Выявление аналогий
Транзитивное заключение – не единственная операция логического вывода, которую совершают животные. Некоторым видам, по крайней мере антропоидам, доступно выполнение и еще одной операции логического вывода – построения аналогий. Этот вид индуктивного мышления впервые исследовал также Д. Примэк (Premack 1983; Gillan et al. 1981). Как и тест на транзитивное заключение, он входил в разработанную Примэком программу изучения тех высших когнитивных функций антропоидов, которые, по его предположениям, могли быть связаны с усвоением языка-посредника.
Наряду с выявлением «функциональных» аналогий между предметами Сара продемонстрировала также понимание отношений пропорций разнородных предметов. Например, из нескольких возможных вариантов они выбирали 1/4 яблока, как стимул, аналогичный стакану, на 1/4 заполненному водой.
Первые эксперименты Д. Примэка позволяли предполагать, что подобная операция доступна только обезьянам, обученным языку-посреднику, однако более поздние эксперименты (Tanaka 1996; Thompson et al. 1997; Thompson, Oden 2000; Vonk 2003) этого не подтвердили. Например, Томпсон и его коллеги (Tompson et al. 1997) провели более формализованный эксперимент. Шимпанзе, которые были обучены выбору по образцу и у которых было сформировано обобщение по признаку «соответствие», оказались способны выбирать стимул, элементы которого были объединены по такому же принципу, как и элементы стимула-образца. Например, образец состоял из двух идентичных фигур (АА), а стимулы для сравнения – из других фигур, либо также одинаковых (ВВ), либо различных (CF). Единственным признаком соответствия между образцом (АА) и одним из стимулов (ВВ) была аналогия в соотношении компонентов стимула. В этом случае для правильного выбора животное должно было использовать более отвлеченный смысл понятия «соответствие», т. е. организованный по тому же принципу (relational matching).
Более того, недавно появились данные о том, что не только антропоиды, но и павианы способны решать экспериментальные задачи, основанные на сопоставлении соотношений в организации стимулов (Bovet, Vauclair 2001; Fagot et al. 2001). Однако в отличие от антропоидов, спонтанно воспринимающих этот признак, павианам для выявления сходства по аналогии соотношений требовалось длительное обучение – порядка 15 тысяч предъявлений.
Из других видов животных эта когнитивная функция пока найдена только у ворон (рис. 12, см. вклейку), которые оказались способны выявлять аналогию в организации двухкомпонентного образца и одного из стимулов в условиях, когда между последними не было никакого физического сходства, но один из них соответствовал образцу по соотношению размера, формы или цвета составляющих их элементов (Zorina, Smirnova 2005).
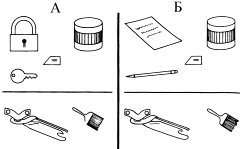
Эксперимент, где впервые была продемонстрирована способность шимпанзе к выявлению аналогий, давно стал классическим. Его проводили с шимпанзе Сарой, которая была второй после Уошо обезьяной, овладевшей небольшим запасом знаков. В частности, в ее лексикон входили «слова» одинаковый, тождественный и разный. В одном из опытов (рис. 11) ей показывали замóк и ключ, рядом (симметрично замку) ставили банку с гуашью, а между ними помещали знак тождества, оставив свободное место рядом с ключом. Для выбора ей предлагали консервный нож и кисть – предметы, назначение которых она хорошо знала. В этом случае Сара уверенно выбирала нож, который выполнял ту же функцию, что и ключ, – тоже открывал банку. В следующем опыте ей продемонстрировали лист бумаги и карандаш и предложили выбрать из тех же двух предметов то, что составляет аналогичную пару с банкой гуаши; она уверенно выбрала кисть, которая по своим функциям в данном сочетании была аналогична карандашу.
Рис. 11. Тест на выявление функциональной аналогии – пояснение в тексте (по Gillan et al. 1981, с изменениями)
Сара успешно выполнила целый ряд таких тестов на «функциональную аналогию», однако ее способности не следует преувеличивать: как подчеркивает Примэк, когда ей предлагали для выбора не 2, а 5 или 6 предметов, ее достижения уже не были столь впечатляющими.Наряду с выявлением «функциональных» аналогий между предметами Сара продемонстрировала также понимание отношений пропорций разнородных предметов. Например, из нескольких возможных вариантов они выбирали 1/4 яблока, как стимул, аналогичный стакану, на 1/4 заполненному водой.
Первые эксперименты Д. Примэка позволяли предполагать, что подобная операция доступна только обезьянам, обученным языку-посреднику, однако более поздние эксперименты (Tanaka 1996; Thompson et al. 1997; Thompson, Oden 2000; Vonk 2003) этого не подтвердили. Например, Томпсон и его коллеги (Tompson et al. 1997) провели более формализованный эксперимент. Шимпанзе, которые были обучены выбору по образцу и у которых было сформировано обобщение по признаку «соответствие», оказались способны выбирать стимул, элементы которого были объединены по такому же принципу, как и элементы стимула-образца. Например, образец состоял из двух идентичных фигур (АА), а стимулы для сравнения – из других фигур, либо также одинаковых (ВВ), либо различных (CF). Единственным признаком соответствия между образцом (АА) и одним из стимулов (ВВ) была аналогия в соотношении компонентов стимула. В этом случае для правильного выбора животное должно было использовать более отвлеченный смысл понятия «соответствие», т. е. организованный по тому же принципу (relational matching).
Более того, недавно появились данные о том, что не только антропоиды, но и павианы способны решать экспериментальные задачи, основанные на сопоставлении соотношений в организации стимулов (Bovet, Vauclair 2001; Fagot et al. 2001). Однако в отличие от антропоидов, спонтанно воспринимающих этот признак, павианам для выявления сходства по аналогии соотношений требовалось длительное обучение – порядка 15 тысяч предъявлений.
Из других видов животных эта когнитивная функция пока найдена только у ворон (рис. 12, см. вклейку), которые оказались способны выявлять аналогию в организации двухкомпонентного образца и одного из стимулов в условиях, когда между последними не было никакого физического сходства, но один из них соответствовал образцу по соотношению размера, формы или цвета составляющих их элементов (Zorina, Smirnova 2005).
Сравнительная характеристика мышления человекообразных обезьян и перспективы поиска биологических истоков речи человека
После краткого описания основных представлений об элементарном мышлении животных, сложившихся к началу экспериментов по обучению шимпанзе языкам-посредникам, еще раз обратимся к некоторым ключевым вопросам подобных исследований, а также к описанию некоторых тенденций их дальнейшего развития. На протяжении XX века представления о существовании у животных зачатков разума постоянно обсуждались и пересматривались, постепенно эволюционируя от полного (или почти полного отрицания) до той или иной степени признания. К настоящему времени голоса скептиков звучат, но все слабее.
В арсенале современной науки о поведении имеется целый набор методик, которые используются для сравнительного исследования когнитивных способностей животных. Именно количественное и качественное сопоставление всего комплекса современных сведений позволило констатировать, что зачатки мышления имеются у многих животных. Они проявляются в разнообразных формах и могут достигать разного уровня, который зависит от сложности структурно-функциональной организации мозга у данного вида. Простейшие формы (способность к экстраполяции и комбинированию элементов приобретенного опыта, о которых мы почти не упоминали за недостатком места) имеются у большинства изученных видов, включая рептилий (Крушинский 1986), а высшее их проявление – способность к усвоению и использованию символов – выявлено не только у человекообразных обезьян, но также и у дельфинов и у высокоорганизованных птиц: попугаев и врановых. Последний факт говорит о том, что появление когнитивных функций, лежащих в основе возникновения речи человека, восходит к достаточно ранним этапам филогенеза. Это обстоятельство делает еще более оправданным предположение о том, что какие-то зачатки способностей к коммуникации с помощью символов могут иметься у современных ближайших родственников человека.
Упомянутые данные весомо подтверждали пророческое высказывание Ч. Дарвина о наличии преемственности в развитии даже наиболее сложных психических функций человека и высших животных. Совокупность полученных данных ставила под сомнение и весьма распространенное представление о пропасти, отделяющей человека от остального животного мира. В связи с этим на повестку дня снова и уже в более конкретной и доступной прямому анализу форме вставал вопрос о том, имеются ли у животных какие-то способности, которые можно было бы рассматривать как эволюционную предпосылку для возникновения человеческой речи. Этот вопрос поднимался еще в самом начале ХХ в., когда не было реальных возможностей его исследовать, но к 60-м годам ХХ в. конкретные подходы к его анализу появились. Основным объектом исследования были, разумеется, человекообразные обезьяны, а среди них шимпанзе. Общепринято, что это наиболее близкие к человеку представители семейства Pongidae, которые к тому же считались более удобными для лабораторных исследований, чем остальные виды, и получили даже название «лабораторный двойник человека» (Фридман 1972).
В арсенале современной науки о поведении имеется целый набор методик, которые используются для сравнительного исследования когнитивных способностей животных. Именно количественное и качественное сопоставление всего комплекса современных сведений позволило констатировать, что зачатки мышления имеются у многих животных. Они проявляются в разнообразных формах и могут достигать разного уровня, который зависит от сложности структурно-функциональной организации мозга у данного вида. Простейшие формы (способность к экстраполяции и комбинированию элементов приобретенного опыта, о которых мы почти не упоминали за недостатком места) имеются у большинства изученных видов, включая рептилий (Крушинский 1986), а высшее их проявление – способность к усвоению и использованию символов – выявлено не только у человекообразных обезьян, но также и у дельфинов и у высокоорганизованных птиц: попугаев и врановых. Последний факт говорит о том, что появление когнитивных функций, лежащих в основе возникновения речи человека, восходит к достаточно ранним этапам филогенеза. Это обстоятельство делает еще более оправданным предположение о том, что какие-то зачатки способностей к коммуникации с помощью символов могут иметься у современных ближайших родственников человека.
Упомянутые данные весомо подтверждали пророческое высказывание Ч. Дарвина о наличии преемственности в развитии даже наиболее сложных психических функций человека и высших животных. Совокупность полученных данных ставила под сомнение и весьма распространенное представление о пропасти, отделяющей человека от остального животного мира. В связи с этим на повестку дня снова и уже в более конкретной и доступной прямому анализу форме вставал вопрос о том, имеются ли у животных какие-то способности, которые можно было бы рассматривать как эволюционную предпосылку для возникновения человеческой речи. Этот вопрос поднимался еще в самом начале ХХ в., когда не было реальных возможностей его исследовать, но к 60-м годам ХХ в. конкретные подходы к его анализу появились. Основным объектом исследования были, разумеется, человекообразные обезьяны, а среди них шимпанзе. Общепринято, что это наиболее близкие к человеку представители семейства Pongidae, которые к тому же считались более удобными для лабораторных исследований, чем остальные виды, и получили даже название «лабораторный двойник человека» (Фридман 1972).
Эволюция взглядов на интеллект антропоидов
Какие же представления об интеллекте антропоидов сложились к середине 1960-х годов? Взрыв работ, посвященных этой проблеме после открытий В. Келера, обеспечил разностороннюю характеристику их когнитивных способностей, отчасти упомянутую в предыдущей главе. При этом оформились две точки зрения относительно степени близости познавательных способностей антропоидов и человека.
Исходя из имевшихся в тот период данных, А. Н. Леонтьев (а вслед за ним К. Э. Фабри и другие психологи, как, впрочем, и физиологи высшей нервной деятельности) подчеркивал ограниченность интеллекта приматов. К. Э. Фабри сформулировал это общее мнение в своем учебнике «Основы зоопсихологии» (1976/ 2001), и оно с тех пор так и продолжает широко тиражироваться. По его мнению, «интеллектуальные способности обезьян, включая антропоидов, ограничены тем, что вся их психическая деятельность имеет биологическую обусловленность, поэтому они не способны к установлению мысленной связи между одними лишь представлениями и их комбинированием в образы» (с. 260; курсив наш. – З. З., А. С.).
Такого же мнения придерживались многие психологи и физиологи. Так, В. П. Протопопов (1950) на основании исследований способности к обобщению и формирования сложных двигательных навыков у капуцинов, макак и человекообразных обезьян одним из первых заявил, что животным доступно абстрагирование. Однако при этом он подчеркивал, что у обезьян «относительный признак не отвлекается полностью, как это имеет место благодаря слову у человека, а только выделяется в наглядно представленных конкретных объектах. Это абстракция in concreto <…> истинная же полная абстракция (vera) выражается в полном отвлечении признака от реального объекта… и возможна, конечно, лишь у человека в его речевом периоде» (Протопопов 1950, с. 163).
Такое представление об уровне когнитивных способностей антропоидов было достаточно характерно для того периода развития науки, оно прослеживается в большинстве работ того времени. Обобщая эти работы, Н. Н. Ладыгина-Котс (1963, с. 318) писала, что «обезьяны имеют элементарное конкретное образное мышление (интеллект), способны к элементарной абстракции и обобщению, и эти черты приближают их психику к человеческой». При этом автор подчеркивала, что «… их интеллект качественно, принципиально отличен от понятийного мышления человека, имеющего язык, оперирование словами как сигналами сигналов, системой кодов, в то время как звуки обезьян хотя и чрезвычайно многообразны, но выражают лишь их эмоциональное состояние и не имеют направленного характера. Они обладают, как и другие животные, лишь первой сигнальной системой».
Высказываясь столь осторожно, Надежда Николаевна в то же время от монографии к монографии последовательно подводила фундаментальную базу под представление о том, что в психике антропоидов наличествуют «предпосылки человеческого мышления» – и именно так она назвала свою последнюю монографию о когнитивной деятельности шимпанзе (Ладыгина-Котс 1965).
Более радикальную точку зрения высказывал ведущий американский приматолог Р. Йеркс (Yerkes 1943), который писал, что «… результаты экспериментальных исследований подтверждают рабочую гипотезу, согласно которой научение у шимпанзе связано с иными процессами, нежели подкрепление и торможение. <…> Можно предполагать, что в скором времени эти процессы будут рассматриваться как предшественники символического мышления человека».
В 1960-е годы начался пересмотр того представления, что «нет никаких оснований усматривать пропасть между низшими и высшими обезьянами и уподоблять поведение высших обезьян человеческому поведению» (Хильченко 1953, с. 121). Американский психолог Г. Харлоу был одним из первых на Западе, кто пробил брешь в глухой стене примитивных бихевиористских методик, которые ставили крыс и шимпанзе практически на один уровень. Его программа серийного обучения предусматривала выработку нескольких сотен (а у голубей и крыс – даже тысяч) дифференцировочных условных рефлексов одного типа. Это приводило к формированию так называемой установки на обучение (learning set formation), основу которой составлял процесс обобщения. Применение этой методики позволило впервые дать широкую сравнительную характеристику когнитивных способностей представителей разных таксономических групп, причем выявленные различия коррелировали с уровнем структурно-функциональной организации мозга. Один из наиболее интересных результатов этой работы состоял в том, что она доказала существенное превосходство антропоидов над всеми остальными видами приматов, не говоря уже о представителях других отрядов млекопитающих и о птицах. Более того, показатели антропоидов приближались к обнаруженным у детей (Harlow 1958; Warren 1965). Такой метод позволил провести и более тонкое сопоставление способностей разных представителей отряда приматов и даже выявить некую связь между способностью к этому типу когнитивной деятельности и экологическими особенностями вида (Rumbaugh et al. 2000). Ниже мы покажем, как развивались эти представления в следующие десятилетия ХХ века.
Исходя из имевшихся в тот период данных, А. Н. Леонтьев (а вслед за ним К. Э. Фабри и другие психологи, как, впрочем, и физиологи высшей нервной деятельности) подчеркивал ограниченность интеллекта приматов. К. Э. Фабри сформулировал это общее мнение в своем учебнике «Основы зоопсихологии» (1976/ 2001), и оно с тех пор так и продолжает широко тиражироваться. По его мнению, «интеллектуальные способности обезьян, включая антропоидов, ограничены тем, что вся их психическая деятельность имеет биологическую обусловленность, поэтому они не способны к установлению мысленной связи между одними лишь представлениями и их комбинированием в образы» (с. 260; курсив наш. – З. З., А. С.).
Такого же мнения придерживались многие психологи и физиологи. Так, В. П. Протопопов (1950) на основании исследований способности к обобщению и формирования сложных двигательных навыков у капуцинов, макак и человекообразных обезьян одним из первых заявил, что животным доступно абстрагирование. Однако при этом он подчеркивал, что у обезьян «относительный признак не отвлекается полностью, как это имеет место благодаря слову у человека, а только выделяется в наглядно представленных конкретных объектах. Это абстракция in concreto <…> истинная же полная абстракция (vera) выражается в полном отвлечении признака от реального объекта… и возможна, конечно, лишь у человека в его речевом периоде» (Протопопов 1950, с. 163).
Такое представление об уровне когнитивных способностей антропоидов было достаточно характерно для того периода развития науки, оно прослеживается в большинстве работ того времени. Обобщая эти работы, Н. Н. Ладыгина-Котс (1963, с. 318) писала, что «обезьяны имеют элементарное конкретное образное мышление (интеллект), способны к элементарной абстракции и обобщению, и эти черты приближают их психику к человеческой». При этом автор подчеркивала, что «… их интеллект качественно, принципиально отличен от понятийного мышления человека, имеющего язык, оперирование словами как сигналами сигналов, системой кодов, в то время как звуки обезьян хотя и чрезвычайно многообразны, но выражают лишь их эмоциональное состояние и не имеют направленного характера. Они обладают, как и другие животные, лишь первой сигнальной системой».
Высказываясь столь осторожно, Надежда Николаевна в то же время от монографии к монографии последовательно подводила фундаментальную базу под представление о том, что в психике антропоидов наличествуют «предпосылки человеческого мышления» – и именно так она назвала свою последнюю монографию о когнитивной деятельности шимпанзе (Ладыгина-Котс 1965).
Более радикальную точку зрения высказывал ведущий американский приматолог Р. Йеркс (Yerkes 1943), который писал, что «… результаты экспериментальных исследований подтверждают рабочую гипотезу, согласно которой научение у шимпанзе связано с иными процессами, нежели подкрепление и торможение. <…> Можно предполагать, что в скором времени эти процессы будут рассматриваться как предшественники символического мышления человека».
В 1960-е годы начался пересмотр того представления, что «нет никаких оснований усматривать пропасть между низшими и высшими обезьянами и уподоблять поведение высших обезьян человеческому поведению» (Хильченко 1953, с. 121). Американский психолог Г. Харлоу был одним из первых на Западе, кто пробил брешь в глухой стене примитивных бихевиористских методик, которые ставили крыс и шимпанзе практически на один уровень. Его программа серийного обучения предусматривала выработку нескольких сотен (а у голубей и крыс – даже тысяч) дифференцировочных условных рефлексов одного типа. Это приводило к формированию так называемой установки на обучение (learning set formation), основу которой составлял процесс обобщения. Применение этой методики позволило впервые дать широкую сравнительную характеристику когнитивных способностей представителей разных таксономических групп, причем выявленные различия коррелировали с уровнем структурно-функциональной организации мозга. Один из наиболее интересных результатов этой работы состоял в том, что она доказала существенное превосходство антропоидов над всеми остальными видами приматов, не говоря уже о представителях других отрядов млекопитающих и о птицах. Более того, показатели антропоидов приближались к обнаруженным у детей (Harlow 1958; Warren 1965). Такой метод позволил провести и более тонкое сопоставление способностей разных представителей отряда приматов и даже выявить некую связь между способностью к этому типу когнитивной деятельности и экологическими особенностями вида (Rumbaugh et al. 2000). Ниже мы покажем, как развивались эти представления в следующие десятилетия ХХ века.