Страница:
зубчатой передачи.Профиль зуба колеса
1очерчен по эвольвенте
Э
1основной окружности с радиусом r
01
, а профиль зуба колеса 2
—по эвольвенте
Э
2основной окружности с радиусом
r
02.Эвольвенты
Э
1и
Э
2касаются в точке
К.Нормаль к эвольвенте
Э
1в точке
Кно условию построения эвольвенты должна быть касательной к основной окружности с радиусом
r
01, нормаль к эвольвенте
Э
2
—касательной к основной окружности с радиусом
r
02. Отсюда следует, что в Э. з. общая нормаль к соприкасающимся профилям зубьев всегда совпадает с общей касательной к основным окружностям. Эта касательная называется линией зацепления, т.к. по ней перемещается точка касания зубьев при движении колёс. Точка
Рпересечения линии зацепления с межосевой линией
O
1
O
2называется полюсом зацепления, а угол, который составляет линия зацепления с перпендикуляром к межосевой линии, — углом зацепления a
w.
Передаточное отношение
i
12, т. е. отношение угловых скоростей колёс
1и
2, определяется при Э. з. по условию
i
12
= O
2
P/O
1
P =r
02/r
01.Отсюда видно, что при Э. з. изменение межосевого расстояния не влияет на значение передаточного отношения. С изменением этого расстояния изменяется лишь угол зацепления. Э. з. — наиболее распространённый вид зубчатого зацепления.
Н. И. Левитский.
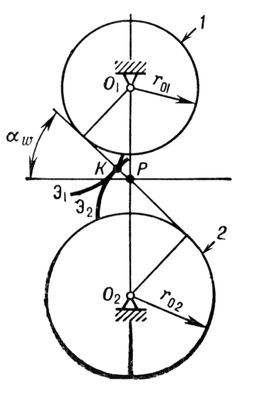 Эвольвентное зацепление: 1 и 2 — зубчатые колёса.
Эвольвентное зацепление: 1 и 2 — зубчатые колёса.
зубоизмерительный прибор
, предназначенный для измерения профиля зуба эвольвентных
зубчатых колёс
в сечении, перпендикулярном оси колёс. Принцип работы Э. заключается в сопоставлении теоретической эвольвенты (см.
Эволюта и эвольвента
),
воспроизводимой прибором, с реальной. По принципиальной схеме механизма, воссоздающего эвольвенту, Э. разделяются на индивидуально-дисковые и универсальные. В индивидуально-дисковых Э. для воспроизведения проверяемой кривой применяются сменные диски, диаметры которых равны диаметрам основной окружности проверяемых колёс. В универсальных Э. для воспроизведения этой кривой используется специальное устройство, настраиваемое на различные радиусы основной окружности. Среди них наибольшее распространение получили Э., в которых используется т. н. механизм обката, содержащий диск постоянного диаметра и рычажно-ленточную передачу, или механизмы с постоянным эвольвентным копиром и рычажной передачей. Более точными являются индивидуально-дисковые Э.
Н. И. Левитский.
зубоизмерительный прибор
, предназначенный для измерения профиля зуба эвольвентных
зубчатых колёс
в сечении, перпендикулярном оси колёс. Принцип работы Э. заключается в сопоставлении теоретической эвольвенты (см.
Эволюта и эвольвента
),
воспроизводимой прибором, с реальной. По принципиальной схеме механизма, воссоздающего эвольвенту, Э. разделяются на индивидуально-дисковые и универсальные. В индивидуально-дисковых Э. для воспроизведения проверяемой кривой применяются сменные диски, диаметры которых равны диаметрам основной окружности проверяемых колёс. В универсальных Э. для воспроизведения этой кривой используется специальное устройство, настраиваемое на различные радиусы основной окружности. Среди них наибольшее распространение получили Э., в которых используется т. н. механизм обката, содержащий диск постоянного диаметра и рычажно-ленточную передачу, или механизмы с постоянным эвольвентным копиром и рычажной передачей. Более точными являются индивидуально-дисковые Э.
Э. изготовляются нескольких типоразмеров для измерения профиля зуба колёс с диаметрами основных окружностей обычно от 40 до 1250
ммнаружного и внутреннего зацепления. Э., как правило, снабжается дополнит. устройством для измерения винтовой линии косозубых зубчатых колёс (см.
Ходомер
), которое обеспечивает перемещение измерит. наконечника регистрирующего устройства вдоль оси измеряемого колеса, согласованное с поворотом. Как правило, результаты измерения регистрируются самописцем. Перспективным направлением в развитии Э. является разработка систем, использующих импульсные преобразователи и мини-ЭВМ. В ряде приборов измеряемое колесо вращается и перемещает измерительный наконечник, а в ЭВМ одновременно поступают импульсы, пропорциональные углу поворота колеса и линейному перемещению наконечника. В ЭВМ эти данные сравниваются с теоретическими, предварительно введёнными в ЭВМ. В др. вариантах этих Э. теоретические угловые и линейные перемещения колесу и измерительному наконечнику задаются с помощью ЭВМ, а регистрирующее устройство воспринимает отклонение реального профиля от теоретического.
Н. Н. Марков.
огибающей
семейства нормалей эвольвенты.
Если линия
lзадана параметрическими уравнениями
х = x(
t),
y = y(
t), то параметрические уравнения её эволюты будут следующие:
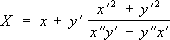 ,
,
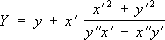
Эвольвенту пространственной кривой можно определить как ортогональную траекторию касательных этой кривой.
Лит.:Рашевский П. К., Курс дифференциальной геометрии, 4 изд., М., 1956.
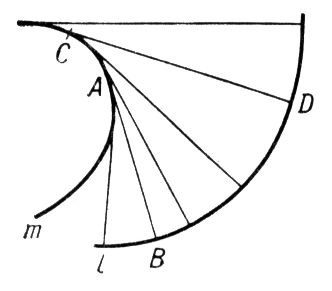 Рис. к ст. Эволюта и эвольвента.
Рис. к ст. Эволюта и эвольвента.