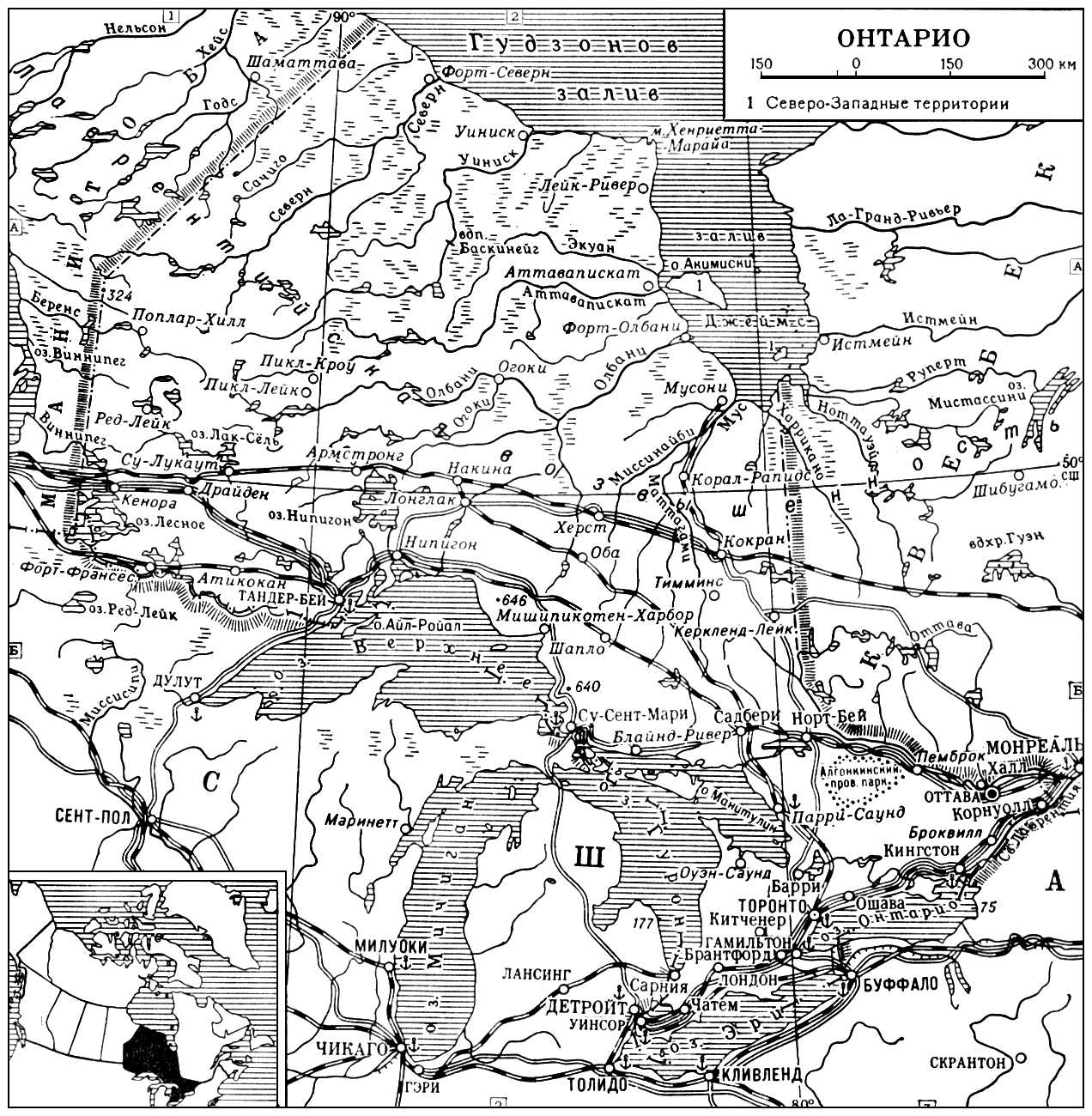
антропонимику
,
топонимику
, зоонимию (собственные имена животных), астронимию, космонимию (название зон и частей Вселенной), теонимию (имена богов) и др. Ономастические исследования помогают выявлять пути миграций и места былого расселения различных народов, языковые и культурные контакты, более древнее состояние языков и соотношение их диалектов. Топонимия (особенно гидронимия) зачастую является единственным источником информации об исчезнувших языках и народах.
Лит.:. Чичагов В. К., Из истории русских имён, отчеств и фамилий, М., 1959; Ташицкий В., Место ономастики среди других гуманитарных наук, «Вопросы языкознания», 1961, № 2: Суперанская А. В., Общая теория имени собственного, М., 1973; Bach A., Deutsche Namenkunde, Bd 1—3, Hdlb., 1952—56; Gardiner A., The theory of proper names, 2 ed., L., 1957.
А. В. Суперанская.
изобразительных слов
. Нередко О. называют условную словесную имитацию звучаний живой и неживой природы и мира вещей («ку-ку», «бум-бум», «а девица — хи-хи-хи! да ха-ха-ха!»). В поэтике — применение (преимущественно в поэзии) обоих видов звуко-подражательных слов для создания звукового образа:
Буду а к а т ь, буду о к а т ь,
Каплю-степь возьму под локоть,
Конь пойдёт подковой ц о к а т ь,
Ё к а т ь селезёнкою.
(А. Тарковский).
кабуки
. В Японии существует традиция передачи сценического имени по наследству актёру-сыну или усыновлённому ученику. Династии О. Байко и О. Кикугоро насчитывают по семь поколений каждая. Наиболее известны из них: О. Байко 7-й (р. 31.8.1915, Токио) — один из лучших исполнителей женских ролей, а также ролей молодых героев. Особенно успешно выступает в любовных сценах и танцевальных пьесах. О. Кикугоро 5-й (4.6.1844, Эдо, — 18.2.1903, Токио), один из выдающихся актёров своего времени, обладал редкой по выразительности сценической внешностью, был искусным танцовщиком. Выступал в исторических и бытовых пьесах классического репертуара кабуки, а также в новых реалистических пьесах (кидзэвамоно). Оказал большое влияние на актёров театра кабуки следующего поколения. О. Кикугоро 6-й (6.6.1885, Токио, — 10.8.1949, там же), сын Кикугоро 5-го. Актёр разнообразного репертуара, выдающийся исполнитель мужских и женских ролей в классических и новых исторических, бытовых и танцевальных пьесах кабуки. В 1931—37 руководил основанной им школой актёров театра кабуки. С 1948 был член японской Академии искусств.
Лит.:Каватакэ Сигэтоси, Нихон энгэки дзэнси, (Всеобщая история японского театра), Токио, 1966; Гудзи М., Японский театр кабуки, пер. с япон., М., 1969.
Л. Д. Гришелёва.
Онсагера уравнение электропроводности
. Открыл (1931) принцип симметрии кинетических коэффициентов (см.
Онсагера теорема
),
послуживший основой феноменологической термодинамики неравновесных процессов. Получил (1942, опубликован в 1944) точное решение двумерной задачи Изинга, предсказывающее логарифмическую зависимость теплоёмкости от температуры вблизи критической точки. Предложил теорию квантовых вихрей в сверхтекучем гелии. Нобелевская премия (1968).
Соч.: Reciprocal relations in irreversible processes, «Physical Review», 1931, v. 38, № 12 p. 2265: Crystal statistics, там же, 1944, v. 65, № 3—4: Statistical hydrodynamics, «Nuovo Cimento», 1949, Supplemento al v. 6, ser. 9, № 2; The electrical properties of ice, в кн.: Electrolytes, Oxf., 1962 (совм. с М. Dupuis).
Д. Н. Зубарев.
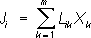 (
i= 1, 2, …,
m) ,(1)
(
i= 1, 2, …,
m) ,(1)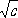 .
.