Страница:
Даже при физиологической беременности у 3 % женщин эритроциты плода проникают через формирующуюся плаценту уже в первом триместре беременности. Число подобных случаев увеличивается во втором триместре беременности, достигая 15 %. В третьем триместре фетальные эритроциты можно обнаружить в кровотоке матери уже у 45 % беременных. Таким образом, с увеличением срока физиологической беременности объем фетальной крови в кровотоке матери возрастает, достигая 30–40 мл в родах.
Объем плодово-материнских трансфузий может значительно увеличиваться на всех этапах беременности при наличии у беременной акушерских и экстрагенитальных заболеваний, которые сопровождаются как повышением проницаемости плаценты, так и ее отслойкой частичной или тотальной. Прежде всего, к такой патологии следует отнести угрозу прерывания беременности, особенно при наличии у беременной клинических признаков начавшегося выкидыша (кровомазания или кровотечения); чистый и сочетанный гестоз; экстрагенитальные заболевания, в генезе которых наблюдается поражение стенок сосудов (заболевания почек, гипертоническая болезнь и другие сердечно-сосудистые заболевания, сахарный диабет различного типа и др.). В этих случаях в кровоток беременной попадает больший объем фетальных эритроцитов, что, при их антигенной несовместимости с эритроцитами матери, может привести к раннему развитию заболевания у плода, в том числе его тяжелых форм, особенно у пациенток, уже имеющих отягощенный иммунологический анамнез.
При попадании антигена в кровоток матери развивается гуморальный иммунный ответ, зависящий от того, первично или повторно антиген попадает в организм матери.
При первичном иммунном ответе у матери образуются иммуноглобулины класса М (IgM), которые в силу своей высокой молекулярной массы не способны проникать к плоду через плацентарный барьер и вызывать аллоиммунную цитопению у плода. При первичном ответе антитела в крови матери не определяются или наблюдаются в невысоком титре.
Иммунный ответ начинается с неспецифического восприятия антигена антиген-презентирующими (антиген-представляющими) клетками, локализующимися в лимфоидных центрах организма матери, таких как селезенка и лимфатические узлы. В фаголизосомах этих клеток под влиянием протеолитических ферментов антигены подвергаются частичному распаду на пептидные фрагменты. Затем эти пептиды, связываются с молекулами II класса HLA системы и возвращаются на поверхность клетки. В результате образуются интердигитатные дендритные клетки, которые при первичном иммунном ответе являются единственными, способными эффективно презентировать антиген Т-хелперам.
Вторичный иммунный ответ возникает, как правило, при повторной беременности. Он характеризуется появлением антител в значительно более высоком титре. При этом образуются иммуноглобулины класса G (IgG), которые участвуют в реакциях агглютинации, преципитации, иммунном лизисе, фиксации комплемента. Однако IgG могут образовываться и при первой беременности. Это чаще происходит при АВО несовместимости крови матери и плода.
При вторичном иммунном ответе антиген наиболее эффективно презентируется В-клетками, которые имеют антигенные рецепторы с высоким сродством (аффинностью) к иммуноглобулину. Т-хелперы распознают антиген, находящийся в комплексе с молекулами HLA II класса. Связь пептида с HLA молекулами зависит от сопоставимости (комплементарности) пептида с пазом (бороздой) на поверхности молекулы HLA. При этом сама способность молекул HLA связывать пептиды различных антигенов находится под генетическим контролем.
Т-хелперы экспрессируют рецепторы, распознающие пептиды, которые расположены в пазах HLA молекул. Считают, что эти пептиды с одной своей стороны состоят из нескольких аминокислот, которые связаны с молекулами HLA II класса, а с другой стороны – из нескольких аминокислот, связанных с рецепторами Т-хелперов. Таким образом, Т-клетки прежде опознают эпитопы антигенов, которые образованы последовательностью простых аминокислот, находящихся в бороздах HLA клеток, до того как эти эпитопы будут распознаны В-клетками и начнется выработка антител.
Взаимодействие между антиген-презентирующими клетками и Т-хелперами облегчается процессами клеточной адгезии и костимулирующими молекулами. Как только антиген презентируется, происходит активация Т-хелперной клетки и секреторного цитокина, такого как интерлейкин-2, присутствие которого необходимо для деления Т-клеток. Этот процесс, в свою очередь, приводит к высвобождению интерлейкинов, которые вызывают процессы дифференцировки В-клеток в клоны, продуцирующие антитела.
В-клетки также могут воспринимать антиген и презентировать его Т-хелперам в связанном с молекулами HLA II класса виде. Антигены, расположенные на поверхности В-клеток, представляют собой тот спектр антигенов, которые В-клетки могут узнавать и выбирать с помощью специфических мембранных антител. Это значит, что Т-хелперы активируют те В-клетки, которые связаны с антигеном, ответственным за первичную активацию Т-хелперов.
При взаимодействии между В-клетками и Т-хелперами инициируются некоторые другие рецептор-лиганд-зависимые взаимодействия и процессы обмена интерлейкинов. Расположенные на В-клетках комплексы пептидов с молекулами II класса HLA и CD40 лигандные рецепторные молекулы связываются с рецепторами Т-клеток и CD40 лигандными молекулами на Т-хелперах. Это является сигналом к пролиферации антиген-специфических В-клеток и их дифференцировке в плазматические клетки, продуцирующие антитела и В-клетки памяти. В конечном итоге антиген-зависимый процесс, направленный на повышение сродства, приводит к гипермутациям В-клеток и секреции антител с повышенным сродством к антигену.
На рисунке 1 представлены этапы гуморального иммунного ответа, развивающегося в организме матери при попадании в ее кровоток чужеродного антигена/антигенов.
Процесс возбуждения и презентации антигенов Т-хелперам происходит с помощью дендритных клеток. Пролиферация Т-хелперов наблюдается при наличии необходимых вторичных сигналов, которые, в свою очередь, дают сигнал пролиферировать и секретировать антитела антиген-специфичным В-клеткам. В этих процессах участвуют костимулирующие молекулы и цитокины.
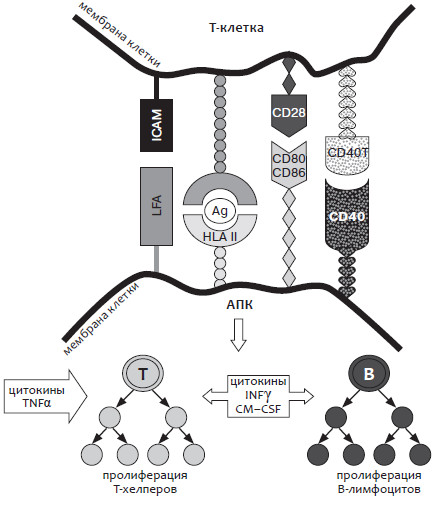
Рис. 1. Этапы гуморального иммунного ответа
АПК – антиген-презентирующая клетка
Как представлено на рисунке 1, костимулирующие молекулы на антиген-презентирующих клетках связываются со своими лигандами на Т-хелперах: так, лейкоцитарный функциональный антиген 3 (LFA-3) связывается с CD2; внутриклеточная адгезивная молекула-1 (ICAM-1) связывается с LFA-1, a CD80/CD86 – с CD28.
В активацию Т-хелперов включаются цитокины, такие как IL-1, IL-6, IL-12, IL-15 и TNFa. При этом в пролиферации Т-клеток важную роль выполняет IL-2. Цитокины, вовлеченные в активацию антиген-презентирующих клеток, такие как интерферон у (IFNy), гранулоцито-макрофагоколоний-стимулирующий фактор (GM-CSF), IL-4 и TNFP обеспечивают возрастание экспрессии таких молекул как HLA класса II и других.
При активации Т-хелперов костимулирующие молекулы связываются с их лигандами на В-клетках. Так, CD28 связывается с CD80/CD86, CD40 лиганд – с CD40, а LFA-1 – с ICAM-1 или ICAM-3, а CD2 связывается IL-3.
Активацию и дифференциацию В-клеток обеспечивают IL-4 и IL-6, а поддерживают активацию Т-хелперов IL-1 и IL-6.
Трансплацентарный перенос антител к плоду
Иммуноглобулины класса G, образующиеся при вторичном иммунном ответе, состоят из 4 субклассов, которые значительно отличаются по степени их агрессивности к эритроцитам. Все четыре субкласса иммуноглобулинов активно переносятся к плоду при помощи клеток синцитиотрофобласта плаценты, которые экспрессируют рецепторы для Fc-доменов IgG.
Считают, что транспорт IgG через плаценту до 24-й недели беременности происходит медленно, поэтому развитие заболевания у плода до этого срока наблюдается сравнительно редко. Однако этот транспорт значительно усиливается во второй половине беременности и достигает максимума перед родами, когда значения IgG у плода могут даже превышать таковые у матери. Скорость транспорта иммуноглобулина через плаценту, видимо, является основным фактором, определяющим тяжесть развивающегося у плода заболевания.
В исследованиях последних лет был выявлен механизм транспорта IgG. В его основе лежит способность рецептора FcRn связывать IgG с относительно высокой аффинностью в условиях кислой pH и с незначительной аффинностью при pH близкой к 7,4, т. е. к pH крови.
Молекула FcRn состоит из a-цепи, которая гомологична молекуле HLA I класса, связанной с β2-микроглобулином. На первом этапе транспорта осуществляется неспецифический жидкофазный пиноцитоз IgG синцитиотрофобластом. Он подобен таковому, осуществляемому при транспорте других пептидов плазмы. Затем наблюдается транспорт в апикальные пузырьки IgG, где pH, соответствующая кислой среде, способствует его связыванию с находящимися там рецепторами FcRn.
FcRn распознает определенные аминокислоты, находящиеся на поверхности IgG, имеющего две цепи, и, следовательно, связанными оказываются две молекулы FcRn. В дальнейшем IgG в связанном состоянии транспортируется через клетку синцитиотрофобласта внутри пузырьков, которые, в конечном счете, растворяются в клетках базальной мембраны. Этот процесс приводит к соприкосновению содержимого пузырьков с pH, соответствующей нейтральной среде. При этом IgG отделяется от FcRn и попадает в плодовую циркуляцию.
Считают, что антитела как бы вакцинируют эритроциты, обеспечивая их узнавание макрофагами, которые экспрессируют рецепторы для Fc части IgG. Сенсибилизированные таким образом клетки крови далее могут уничтожаться макрофагами путем окончательного экстраваскулярного лизиса.
У трети женщин, имеющих анти-D-антитела, они представлены только IgG1 У оставшейся части сенсибилизированных пациенток имеется смесь IgG1 и IgG3. Агрессивными являются иммуноглобулины третьего (IgG3) и первого (IgGj) субклассов, причем IgG3 наиболее активны. Эти антитела обладают способностью активировать комплемент и легко взаимодействовать с Fc-рецепторами фагоцитирующих клеток. Эти свойства отсутствуют у IgG2 и IgG4.
Разрушение или подавление выработки клеток крови плода под действием иммунных механизмов.
Развитие у плода клинических проявлений заболевания
IgG3 и IgG1, проникая вкровотокрезусположител ьного плода, осаждаются на мембране эритроцитов. В большинстве случаев именно эти субклассы антител способны вызывать гемолиз эритроцитов.
Отсутствуют однозначные данные о том, имеется ли связь между тяжестью гемолитической болезни плода и новорожденного и спектром имеющихся у матери агрессивных антител – только IgG1 или смеси IgG1 и IgG3. При этом известно, что концентрация антител в материнском и плодовом кровотоке аналогична.
По данным некоторых авторов, для развития вторичного иммунного ответа, достаточно попадания в кровоток матери 0,1 мл крови плода. Иммуноглобулины класса G вступают в реакцию взаимодействия с Fc-рецепторами фагоцитирующих клеток, в большом количестве расположенных на поверхности плаценты. Для того чтобы вызвать фагоцитоз, клетки должны нести на своей поверхности около 1000–4000 молекул IgGj и всего лишь 135–500 молекул IgG3, что характеризует более высокую агрессивность IgG3. Происходит реакция антиген-антитело. Этот процесс продолжается до тех пор, пока по закону действия масс не наступит равновесие, после чего избыток антител проникает непосредственно к плоду и частично – в околоплодные воды, где происходит их обратное связывание.
Ряд факторов влияет на способность антител вызывать внутриутробный гемолиз эритроцитов. Прежде всего, это непосредственно сама возможность антител, вызывать гемолиз эритроцитов плода, опознавать антигены эритроцитов и связываться с ними. При этом в том случае, если антигены широко представлены в тканях плода, антитела связываются именно с ними, а не с теми антигенами, которые расположены на эритроцитах.
Другим фактором, влияющим на патогенез гемолитической болезни, является специфичность антигенов. Например, антитела к антигенам системы Келл, которые экспрессируются на ранних стадиях развития эритроцитов, могут вызывать анемию путем угнетения эритропоэза или путем стимуляции иммунного разрушения клеток-предшественников эритроцитов. При этом у плода, как было уже упомянуто выше, развивается не гемолитическая, а тяжелая апластическая анемия.
Еще одним важным фактором иммунного ответа является количество антигенов, а, следовательно, плотность их распределения на мембране эритроцитов. Так, некоторые антигены, например системы Резус, хорошо экспрессируются на эритроцитах плода уже на ранних этапах беременности. Другие антигены (например, системы Лютеран) плохо представлены на клетках плода. Антитела, которые представлены на эритроцитах плода с небольшой плотностью, не способны вызывать заболевание. Так, например, тяжесть гемолитической болезни, обусловленной анти-А и анти-В антителами системы АВО, зависит от количества на эритроцитах антигенов А и В.
Кроме того, значимым фактором силы иммунного ответа является сама структура антигена. Так, антитела к антигенам, расположенным относительно далеко от двойной липидной мембраны, усиливают распознавание эритроцитов моноцитами. По этой причине, например, анти-К антитела более эффективно способствуют адгезии, фагоцитозу и лизису эритроцитов, чем анти-D-антитела.
Совокупность вышеперечисленных факторов объясняет, в том числе редкую встречаемость тяжелых форм гемолитической болезни плода и новорожденного при АВО несовместимости крови плода и матери. Она объясняется тем, что:
1) большое количество анти-А и анти-В антител связываются с А и В антигенами, растворенными в тканях плаценты, плазме крови, околоплодных водах;
2) структура А и В антигенов фетальных эритроцитов позволяет связывать только небольшое количество антител, даже если их много;
3) преимущественным наличием анти-А и анти-В антител в виде IgG2.
Таким образом, становится понятно, что не только идентификация антиэритроцитарных антител в крови матери, но и определение их специфичности имеет большое значение для прогноза развития патологического процесса у плода.
Массивное разрушение эритроцитов, наблюдающееся у плода при гемолитическом характере аллоиммунной анемии, сопровождается образованием в большом количестве непрямого билирубина. При этом процессы его перевода в прямой билирубин нарушаются.
Известно, что непрямой билирубин обладает выраженным токсическим действием на ткани, богатые липидами, такие как ткани печени и мозга. Кроме того, непрямой билирубин не выводится почками. У плода он активно элиминируется из кровеносного русла через плаценту в кровоток матери, где происходит его инактивация. Однако после рождения токсическое влияние непрямого билирубина, образующегося в результате продолжающегося гемолиза эритроцитов под влиянием циркулирующих в крови новорожденного антител, может приводить к развитию у него печеночной недостаточности и выраженной энцефалопатии.
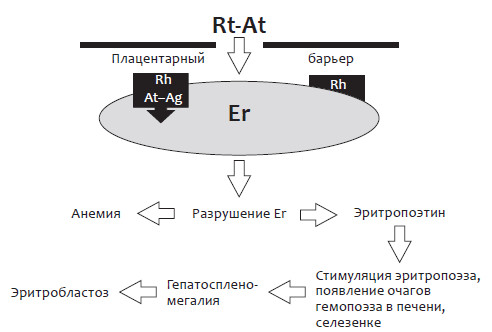
Рис. 2. Патогенез анемии и эритробластоза у плода при гемолитической болезни
На рисунке 2 показан каскад патологических процессов, развивающихся у плода под влиянием материнских антител.
При гемолизе эритроцитов высвобождается большое количество эритропоэтина, который активизирует экстрамедуллярный гемопоэз в органах экстрамедуллярного кроветворения плода – печени и селезенке.
Этот процесс носит адаптивно-приспособительный характер и направлен на компенсацию прогрессирующей анемии, сопровождающейся снижением кислородной емкости крови. Однако подобный тип кроветворения характеризуется незавершенным течением, в результате которого формируется большое количество бластных форм эритроцитов. Эта компенсаторная реакция, наблюдающаяся в крови плода и новорожденного, легла в основу диагностического теста, свидетельствующего о наличии гемолитической болезни у плода и новорожденного. Она же дала одно из названий самого заболевания – эритробластоз.
В печени плода очаги экстрамедуллярного кроветворения постепенно увеличиваются, сливаются между собой и начинают занимать большую часть ее паренхимы, вызывая нарушение работы гепатоцитов и их ферментативных систем, а также окклюзию желчевыводящих протоков. Печень перегружается железом, билирубиновыми пигментами, нарушается ее функция, прежде всего, белковосинтетическая. Следствием этого является прогрессирующая гипопротеинемия.
При разрушении эритроцитов высвобождается большое количество биологически активных веществ, включая ферменты тромбогенеза и фибринолиза. Они способствуют развитию у плода и новорожденного ДВС-синдрома, вследствие которого нарушается циркуляция крови, появляются кровоизлияния, нарушается трофика тканей.
При развитии хронической гипоксии и ацидоза, наблюдается компенсаторное увеличение сердечного выброса и минутного объема. Наблюдается гипертрофия миокарда, гипертензия в портальных и пуповинных венах, усиливается проницаемость сосудов. У плода формируется компенсаторно-приспособительная реакция – гипердинамический тип кровообращения, обеспечивающий защиту жизненно важных органов от нарастающей гипоксии.
Повышается центральное венозное давление, затрудняется ток лимфы по магистральным лимфатическим сосудам, нарушается отток интерстициальной жидкости. В тканях и серозных полостях у плода накапливается жидкость, развивается генерализованный отек. Ему способствует и прогрессирующая сердечная недостаточность, сопровождающаяся застоем в большом круге кровообращения.
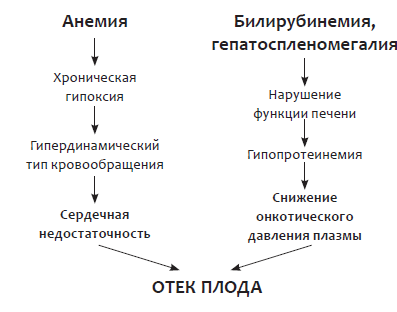
Рис. 3. Патогенез отечного синдрома у плода при гемолитической болезни
При отсутствии адекватного лечения тяжелой анемии, гематокрит в крови плода снижается ниже 12–14 %. Следствием этих процессов являются накопление жидкости в тканях и полостях – асцит, выпот в перикарде и плевральных полостях вплоть до развития у плода анасарки. Патогенез отечного синдрома, развивающегося у плода при изоиммунизации, представлен на рисунке 3.
Отечная форма заболевания, как правило, сопровождается антенатальной или постнатальной гибелью плода и новорожденного. При этом у матери может рано развиваться гестоз, часто наблюдаются симптомы плацентарной недостаточности.
Клинические симптомы заболевания у новорожденных зависят от патогенетического варианта течения аллоиммунной эритропении. При желтушном и желтушно-анемическом вариантах гемолитической болезни в крови новорожденного наблюдается рост содержания билирубина, темп которого зависит от агрессивности иммунного процесса.
При ABO несовместимости крови плода и матери у новорожденного преобладает легкое течение гемолитической болезни, проявляющееся желтушным окрашиванием кожных покровов, умеренным, часто длительным повышением билирубина в крови. Печень и селезенка у ребенка, как правило, не увеличиваются. В редких случаях наблюдается умеренная анемия.
При тяжелом течении заболевания, развивающегося, преимущественно, при Rh-несовместимости крови матери и плода, клинические проявления могут наблюдаться и нарастать уже с первых часов жизни новорожденного. При этом у ребенка наблюдается бледно-желтушный цвет кожных покровов и слизистых, увеличивается печень и селезенка. В крови быстро накапливается непрямой билирубин, обладающий выраженным токсическим действием на ткани, богатые липидами, такие как мозг и печень. При этом непрямой билирубин не выводится почками. Накопление непрямого билирубина обусловлено нарушением процессов его перевода в прямой билирубин. Рефлексы и мышечный тонус у новорожденного, как правило, снижены. В клинической картине заболевания преобладают при анемической форме симптомы, обусловленные эритро– и тромбоцитопенией, при желтушной и желтушноанемической форме – гипербилирубинемией.
При отсутствии лечения гипербилирубинемия может нарастать. Считают, что патогенное действие непрямого билирубина на организм основывается на выключении из метаболизма ферментов, обеспечивающих тканевое дыхание. В результате этого прекращается окислительное фосфорилирование. При крайней степени тяжести заболевания «гепатотоксичный гемопоэз» может стать причиной развития печеночной недостаточности и билирубиновой энцефалопатии. При этом жирорастворимый неконъюгированый билирубин накапливается в базальных ядрах мозга. В крайне тяжелых случаях возможно развитие ядерной желтухи, первыми симптомами которой у новорожденного являются снижение сосательного рефлекса и ригидность затылочных мышц. В дальнейшем, при отсутствии лечения, у ребенка прогрессируют гиперестезия, беспокойство, появляются глазодвигательные нарушения («симптом заходящего солнца»), расстройства дыхания и сердцебиения на фоне судорог. Температура тела повышается до 40–41 °C. Она обусловлена пирогенным влиянием билирубина.
В агональном периоде у новорожденных наблюдаются геморрагические явления в виде кровоизлияний в кожу, кишечник, легкие с развитием отека и пневмонии с последующим летальным исходом.
В отдельных случаях может наблюдаться относительное выздоровление. Однако в последующем у ребенка проявляются неврологические нарушения, обусловленные токсическим влиянием билирубина, от минимальных мозговых дисфункций и небольших моторных нарушений до значительных расстройств, сочетающихся с глухотой и нарушением интеллекта. При отечной форме гемолитической болезни в случае живорождения, чаще в результате преждевременных родов, в клинической картине заболевания преобладает отечный синдром – выраженный асцит, сердечная недостаточность с наличием гидроперикарда, отека легких с гидротораксом, а также значительное увеличение печени и селезенки, резкая бледность кожных покровов. Новорожденный часто погибает на фоне развития полиорганной недостаточности.
Таким образом, неблагоприятные перинатальные исходы при изоиммунизации во время беременности главным образом обусловлены в антенатальном периоде развитием у плода тяжелого анемического синдрома, а в постнатальной жизни – гипербилирубинемией, сопровождающейся поражением центральной нервной системы с последующей полиорганной недостаточностью.
Литература к главе I
Глава II Антенатальная диагностика аллоиммунных эритропений
Объем плодово-материнских трансфузий может значительно увеличиваться на всех этапах беременности при наличии у беременной акушерских и экстрагенитальных заболеваний, которые сопровождаются как повышением проницаемости плаценты, так и ее отслойкой частичной или тотальной. Прежде всего, к такой патологии следует отнести угрозу прерывания беременности, особенно при наличии у беременной клинических признаков начавшегося выкидыша (кровомазания или кровотечения); чистый и сочетанный гестоз; экстрагенитальные заболевания, в генезе которых наблюдается поражение стенок сосудов (заболевания почек, гипертоническая болезнь и другие сердечно-сосудистые заболевания, сахарный диабет различного типа и др.). В этих случаях в кровоток беременной попадает больший объем фетальных эритроцитов, что, при их антигенной несовместимости с эритроцитами матери, может привести к раннему развитию заболевания у плода, в том числе его тяжелых форм, особенно у пациенток, уже имеющих отягощенный иммунологический анамнез.
При попадании антигена в кровоток матери развивается гуморальный иммунный ответ, зависящий от того, первично или повторно антиген попадает в организм матери.
При первичном иммунном ответе у матери образуются иммуноглобулины класса М (IgM), которые в силу своей высокой молекулярной массы не способны проникать к плоду через плацентарный барьер и вызывать аллоиммунную цитопению у плода. При первичном ответе антитела в крови матери не определяются или наблюдаются в невысоком титре.
Иммунный ответ начинается с неспецифического восприятия антигена антиген-презентирующими (антиген-представляющими) клетками, локализующимися в лимфоидных центрах организма матери, таких как селезенка и лимфатические узлы. В фаголизосомах этих клеток под влиянием протеолитических ферментов антигены подвергаются частичному распаду на пептидные фрагменты. Затем эти пептиды, связываются с молекулами II класса HLA системы и возвращаются на поверхность клетки. В результате образуются интердигитатные дендритные клетки, которые при первичном иммунном ответе являются единственными, способными эффективно презентировать антиген Т-хелперам.
Вторичный иммунный ответ возникает, как правило, при повторной беременности. Он характеризуется появлением антител в значительно более высоком титре. При этом образуются иммуноглобулины класса G (IgG), которые участвуют в реакциях агглютинации, преципитации, иммунном лизисе, фиксации комплемента. Однако IgG могут образовываться и при первой беременности. Это чаще происходит при АВО несовместимости крови матери и плода.
При вторичном иммунном ответе антиген наиболее эффективно презентируется В-клетками, которые имеют антигенные рецепторы с высоким сродством (аффинностью) к иммуноглобулину. Т-хелперы распознают антиген, находящийся в комплексе с молекулами HLA II класса. Связь пептида с HLA молекулами зависит от сопоставимости (комплементарности) пептида с пазом (бороздой) на поверхности молекулы HLA. При этом сама способность молекул HLA связывать пептиды различных антигенов находится под генетическим контролем.
Т-хелперы экспрессируют рецепторы, распознающие пептиды, которые расположены в пазах HLA молекул. Считают, что эти пептиды с одной своей стороны состоят из нескольких аминокислот, которые связаны с молекулами HLA II класса, а с другой стороны – из нескольких аминокислот, связанных с рецепторами Т-хелперов. Таким образом, Т-клетки прежде опознают эпитопы антигенов, которые образованы последовательностью простых аминокислот, находящихся в бороздах HLA клеток, до того как эти эпитопы будут распознаны В-клетками и начнется выработка антител.
Взаимодействие между антиген-презентирующими клетками и Т-хелперами облегчается процессами клеточной адгезии и костимулирующими молекулами. Как только антиген презентируется, происходит активация Т-хелперной клетки и секреторного цитокина, такого как интерлейкин-2, присутствие которого необходимо для деления Т-клеток. Этот процесс, в свою очередь, приводит к высвобождению интерлейкинов, которые вызывают процессы дифференцировки В-клеток в клоны, продуцирующие антитела.
В-клетки также могут воспринимать антиген и презентировать его Т-хелперам в связанном с молекулами HLA II класса виде. Антигены, расположенные на поверхности В-клеток, представляют собой тот спектр антигенов, которые В-клетки могут узнавать и выбирать с помощью специфических мембранных антител. Это значит, что Т-хелперы активируют те В-клетки, которые связаны с антигеном, ответственным за первичную активацию Т-хелперов.
При взаимодействии между В-клетками и Т-хелперами инициируются некоторые другие рецептор-лиганд-зависимые взаимодействия и процессы обмена интерлейкинов. Расположенные на В-клетках комплексы пептидов с молекулами II класса HLA и CD40 лигандные рецепторные молекулы связываются с рецепторами Т-клеток и CD40 лигандными молекулами на Т-хелперах. Это является сигналом к пролиферации антиген-специфических В-клеток и их дифференцировке в плазматические клетки, продуцирующие антитела и В-клетки памяти. В конечном итоге антиген-зависимый процесс, направленный на повышение сродства, приводит к гипермутациям В-клеток и секреции антител с повышенным сродством к антигену.
На рисунке 1 представлены этапы гуморального иммунного ответа, развивающегося в организме матери при попадании в ее кровоток чужеродного антигена/антигенов.
Процесс возбуждения и презентации антигенов Т-хелперам происходит с помощью дендритных клеток. Пролиферация Т-хелперов наблюдается при наличии необходимых вторичных сигналов, которые, в свою очередь, дают сигнал пролиферировать и секретировать антитела антиген-специфичным В-клеткам. В этих процессах участвуют костимулирующие молекулы и цитокины.
Рис. 1. Этапы гуморального иммунного ответа
АПК – антиген-презентирующая клетка
Как представлено на рисунке 1, костимулирующие молекулы на антиген-презентирующих клетках связываются со своими лигандами на Т-хелперах: так, лейкоцитарный функциональный антиген 3 (LFA-3) связывается с CD2; внутриклеточная адгезивная молекула-1 (ICAM-1) связывается с LFA-1, a CD80/CD86 – с CD28.
В активацию Т-хелперов включаются цитокины, такие как IL-1, IL-6, IL-12, IL-15 и TNFa. При этом в пролиферации Т-клеток важную роль выполняет IL-2. Цитокины, вовлеченные в активацию антиген-презентирующих клеток, такие как интерферон у (IFNy), гранулоцито-макрофагоколоний-стимулирующий фактор (GM-CSF), IL-4 и TNFP обеспечивают возрастание экспрессии таких молекул как HLA класса II и других.
При активации Т-хелперов костимулирующие молекулы связываются с их лигандами на В-клетках. Так, CD28 связывается с CD80/CD86, CD40 лиганд – с CD40, а LFA-1 – с ICAM-1 или ICAM-3, а CD2 связывается IL-3.
Активацию и дифференциацию В-клеток обеспечивают IL-4 и IL-6, а поддерживают активацию Т-хелперов IL-1 и IL-6.
Трансплацентарный перенос антител к плоду
Иммуноглобулины класса G, образующиеся при вторичном иммунном ответе, состоят из 4 субклассов, которые значительно отличаются по степени их агрессивности к эритроцитам. Все четыре субкласса иммуноглобулинов активно переносятся к плоду при помощи клеток синцитиотрофобласта плаценты, которые экспрессируют рецепторы для Fc-доменов IgG.
Считают, что транспорт IgG через плаценту до 24-й недели беременности происходит медленно, поэтому развитие заболевания у плода до этого срока наблюдается сравнительно редко. Однако этот транспорт значительно усиливается во второй половине беременности и достигает максимума перед родами, когда значения IgG у плода могут даже превышать таковые у матери. Скорость транспорта иммуноглобулина через плаценту, видимо, является основным фактором, определяющим тяжесть развивающегося у плода заболевания.
В исследованиях последних лет был выявлен механизм транспорта IgG. В его основе лежит способность рецептора FcRn связывать IgG с относительно высокой аффинностью в условиях кислой pH и с незначительной аффинностью при pH близкой к 7,4, т. е. к pH крови.
Молекула FcRn состоит из a-цепи, которая гомологична молекуле HLA I класса, связанной с β2-микроглобулином. На первом этапе транспорта осуществляется неспецифический жидкофазный пиноцитоз IgG синцитиотрофобластом. Он подобен таковому, осуществляемому при транспорте других пептидов плазмы. Затем наблюдается транспорт в апикальные пузырьки IgG, где pH, соответствующая кислой среде, способствует его связыванию с находящимися там рецепторами FcRn.
FcRn распознает определенные аминокислоты, находящиеся на поверхности IgG, имеющего две цепи, и, следовательно, связанными оказываются две молекулы FcRn. В дальнейшем IgG в связанном состоянии транспортируется через клетку синцитиотрофобласта внутри пузырьков, которые, в конечном счете, растворяются в клетках базальной мембраны. Этот процесс приводит к соприкосновению содержимого пузырьков с pH, соответствующей нейтральной среде. При этом IgG отделяется от FcRn и попадает в плодовую циркуляцию.
Считают, что антитела как бы вакцинируют эритроциты, обеспечивая их узнавание макрофагами, которые экспрессируют рецепторы для Fc части IgG. Сенсибилизированные таким образом клетки крови далее могут уничтожаться макрофагами путем окончательного экстраваскулярного лизиса.
У трети женщин, имеющих анти-D-антитела, они представлены только IgG1 У оставшейся части сенсибилизированных пациенток имеется смесь IgG1 и IgG3. Агрессивными являются иммуноглобулины третьего (IgG3) и первого (IgGj) субклассов, причем IgG3 наиболее активны. Эти антитела обладают способностью активировать комплемент и легко взаимодействовать с Fc-рецепторами фагоцитирующих клеток. Эти свойства отсутствуют у IgG2 и IgG4.
Разрушение или подавление выработки клеток крови плода под действием иммунных механизмов.
Развитие у плода клинических проявлений заболевания
IgG3 и IgG1, проникая вкровотокрезусположител ьного плода, осаждаются на мембране эритроцитов. В большинстве случаев именно эти субклассы антител способны вызывать гемолиз эритроцитов.
Отсутствуют однозначные данные о том, имеется ли связь между тяжестью гемолитической болезни плода и новорожденного и спектром имеющихся у матери агрессивных антител – только IgG1 или смеси IgG1 и IgG3. При этом известно, что концентрация антител в материнском и плодовом кровотоке аналогична.
По данным некоторых авторов, для развития вторичного иммунного ответа, достаточно попадания в кровоток матери 0,1 мл крови плода. Иммуноглобулины класса G вступают в реакцию взаимодействия с Fc-рецепторами фагоцитирующих клеток, в большом количестве расположенных на поверхности плаценты. Для того чтобы вызвать фагоцитоз, клетки должны нести на своей поверхности около 1000–4000 молекул IgGj и всего лишь 135–500 молекул IgG3, что характеризует более высокую агрессивность IgG3. Происходит реакция антиген-антитело. Этот процесс продолжается до тех пор, пока по закону действия масс не наступит равновесие, после чего избыток антител проникает непосредственно к плоду и частично – в околоплодные воды, где происходит их обратное связывание.
Ряд факторов влияет на способность антител вызывать внутриутробный гемолиз эритроцитов. Прежде всего, это непосредственно сама возможность антител, вызывать гемолиз эритроцитов плода, опознавать антигены эритроцитов и связываться с ними. При этом в том случае, если антигены широко представлены в тканях плода, антитела связываются именно с ними, а не с теми антигенами, которые расположены на эритроцитах.
Другим фактором, влияющим на патогенез гемолитической болезни, является специфичность антигенов. Например, антитела к антигенам системы Келл, которые экспрессируются на ранних стадиях развития эритроцитов, могут вызывать анемию путем угнетения эритропоэза или путем стимуляции иммунного разрушения клеток-предшественников эритроцитов. При этом у плода, как было уже упомянуто выше, развивается не гемолитическая, а тяжелая апластическая анемия.
Еще одним важным фактором иммунного ответа является количество антигенов, а, следовательно, плотность их распределения на мембране эритроцитов. Так, некоторые антигены, например системы Резус, хорошо экспрессируются на эритроцитах плода уже на ранних этапах беременности. Другие антигены (например, системы Лютеран) плохо представлены на клетках плода. Антитела, которые представлены на эритроцитах плода с небольшой плотностью, не способны вызывать заболевание. Так, например, тяжесть гемолитической болезни, обусловленной анти-А и анти-В антителами системы АВО, зависит от количества на эритроцитах антигенов А и В.
Кроме того, значимым фактором силы иммунного ответа является сама структура антигена. Так, антитела к антигенам, расположенным относительно далеко от двойной липидной мембраны, усиливают распознавание эритроцитов моноцитами. По этой причине, например, анти-К антитела более эффективно способствуют адгезии, фагоцитозу и лизису эритроцитов, чем анти-D-антитела.
Совокупность вышеперечисленных факторов объясняет, в том числе редкую встречаемость тяжелых форм гемолитической болезни плода и новорожденного при АВО несовместимости крови плода и матери. Она объясняется тем, что:
1) большое количество анти-А и анти-В антител связываются с А и В антигенами, растворенными в тканях плаценты, плазме крови, околоплодных водах;
2) структура А и В антигенов фетальных эритроцитов позволяет связывать только небольшое количество антител, даже если их много;
3) преимущественным наличием анти-А и анти-В антител в виде IgG2.
Таким образом, становится понятно, что не только идентификация антиэритроцитарных антител в крови матери, но и определение их специфичности имеет большое значение для прогноза развития патологического процесса у плода.
Массивное разрушение эритроцитов, наблюдающееся у плода при гемолитическом характере аллоиммунной анемии, сопровождается образованием в большом количестве непрямого билирубина. При этом процессы его перевода в прямой билирубин нарушаются.
Известно, что непрямой билирубин обладает выраженным токсическим действием на ткани, богатые липидами, такие как ткани печени и мозга. Кроме того, непрямой билирубин не выводится почками. У плода он активно элиминируется из кровеносного русла через плаценту в кровоток матери, где происходит его инактивация. Однако после рождения токсическое влияние непрямого билирубина, образующегося в результате продолжающегося гемолиза эритроцитов под влиянием циркулирующих в крови новорожденного антител, может приводить к развитию у него печеночной недостаточности и выраженной энцефалопатии.
Рис. 2. Патогенез анемии и эритробластоза у плода при гемолитической болезни
На рисунке 2 показан каскад патологических процессов, развивающихся у плода под влиянием материнских антител.
При гемолизе эритроцитов высвобождается большое количество эритропоэтина, который активизирует экстрамедуллярный гемопоэз в органах экстрамедуллярного кроветворения плода – печени и селезенке.
Этот процесс носит адаптивно-приспособительный характер и направлен на компенсацию прогрессирующей анемии, сопровождающейся снижением кислородной емкости крови. Однако подобный тип кроветворения характеризуется незавершенным течением, в результате которого формируется большое количество бластных форм эритроцитов. Эта компенсаторная реакция, наблюдающаяся в крови плода и новорожденного, легла в основу диагностического теста, свидетельствующего о наличии гемолитической болезни у плода и новорожденного. Она же дала одно из названий самого заболевания – эритробластоз.
В печени плода очаги экстрамедуллярного кроветворения постепенно увеличиваются, сливаются между собой и начинают занимать большую часть ее паренхимы, вызывая нарушение работы гепатоцитов и их ферментативных систем, а также окклюзию желчевыводящих протоков. Печень перегружается железом, билирубиновыми пигментами, нарушается ее функция, прежде всего, белковосинтетическая. Следствием этого является прогрессирующая гипопротеинемия.
При разрушении эритроцитов высвобождается большое количество биологически активных веществ, включая ферменты тромбогенеза и фибринолиза. Они способствуют развитию у плода и новорожденного ДВС-синдрома, вследствие которого нарушается циркуляция крови, появляются кровоизлияния, нарушается трофика тканей.
При развитии хронической гипоксии и ацидоза, наблюдается компенсаторное увеличение сердечного выброса и минутного объема. Наблюдается гипертрофия миокарда, гипертензия в портальных и пуповинных венах, усиливается проницаемость сосудов. У плода формируется компенсаторно-приспособительная реакция – гипердинамический тип кровообращения, обеспечивающий защиту жизненно важных органов от нарастающей гипоксии.
Повышается центральное венозное давление, затрудняется ток лимфы по магистральным лимфатическим сосудам, нарушается отток интерстициальной жидкости. В тканях и серозных полостях у плода накапливается жидкость, развивается генерализованный отек. Ему способствует и прогрессирующая сердечная недостаточность, сопровождающаяся застоем в большом круге кровообращения.
Рис. 3. Патогенез отечного синдрома у плода при гемолитической болезни
При отсутствии адекватного лечения тяжелой анемии, гематокрит в крови плода снижается ниже 12–14 %. Следствием этих процессов являются накопление жидкости в тканях и полостях – асцит, выпот в перикарде и плевральных полостях вплоть до развития у плода анасарки. Патогенез отечного синдрома, развивающегося у плода при изоиммунизации, представлен на рисунке 3.
Отечная форма заболевания, как правило, сопровождается антенатальной или постнатальной гибелью плода и новорожденного. При этом у матери может рано развиваться гестоз, часто наблюдаются симптомы плацентарной недостаточности.
Клинические симптомы заболевания у новорожденных зависят от патогенетического варианта течения аллоиммунной эритропении. При желтушном и желтушно-анемическом вариантах гемолитической болезни в крови новорожденного наблюдается рост содержания билирубина, темп которого зависит от агрессивности иммунного процесса.
При ABO несовместимости крови плода и матери у новорожденного преобладает легкое течение гемолитической болезни, проявляющееся желтушным окрашиванием кожных покровов, умеренным, часто длительным повышением билирубина в крови. Печень и селезенка у ребенка, как правило, не увеличиваются. В редких случаях наблюдается умеренная анемия.
При тяжелом течении заболевания, развивающегося, преимущественно, при Rh-несовместимости крови матери и плода, клинические проявления могут наблюдаться и нарастать уже с первых часов жизни новорожденного. При этом у ребенка наблюдается бледно-желтушный цвет кожных покровов и слизистых, увеличивается печень и селезенка. В крови быстро накапливается непрямой билирубин, обладающий выраженным токсическим действием на ткани, богатые липидами, такие как мозг и печень. При этом непрямой билирубин не выводится почками. Накопление непрямого билирубина обусловлено нарушением процессов его перевода в прямой билирубин. Рефлексы и мышечный тонус у новорожденного, как правило, снижены. В клинической картине заболевания преобладают при анемической форме симптомы, обусловленные эритро– и тромбоцитопенией, при желтушной и желтушноанемической форме – гипербилирубинемией.
При отсутствии лечения гипербилирубинемия может нарастать. Считают, что патогенное действие непрямого билирубина на организм основывается на выключении из метаболизма ферментов, обеспечивающих тканевое дыхание. В результате этого прекращается окислительное фосфорилирование. При крайней степени тяжести заболевания «гепатотоксичный гемопоэз» может стать причиной развития печеночной недостаточности и билирубиновой энцефалопатии. При этом жирорастворимый неконъюгированый билирубин накапливается в базальных ядрах мозга. В крайне тяжелых случаях возможно развитие ядерной желтухи, первыми симптомами которой у новорожденного являются снижение сосательного рефлекса и ригидность затылочных мышц. В дальнейшем, при отсутствии лечения, у ребенка прогрессируют гиперестезия, беспокойство, появляются глазодвигательные нарушения («симптом заходящего солнца»), расстройства дыхания и сердцебиения на фоне судорог. Температура тела повышается до 40–41 °C. Она обусловлена пирогенным влиянием билирубина.
В агональном периоде у новорожденных наблюдаются геморрагические явления в виде кровоизлияний в кожу, кишечник, легкие с развитием отека и пневмонии с последующим летальным исходом.
В отдельных случаях может наблюдаться относительное выздоровление. Однако в последующем у ребенка проявляются неврологические нарушения, обусловленные токсическим влиянием билирубина, от минимальных мозговых дисфункций и небольших моторных нарушений до значительных расстройств, сочетающихся с глухотой и нарушением интеллекта. При отечной форме гемолитической болезни в случае живорождения, чаще в результате преждевременных родов, в клинической картине заболевания преобладает отечный синдром – выраженный асцит, сердечная недостаточность с наличием гидроперикарда, отека легких с гидротораксом, а также значительное увеличение печени и селезенки, резкая бледность кожных покровов. Новорожденный часто погибает на фоне развития полиорганной недостаточности.
Таким образом, неблагоприятные перинатальные исходы при изоиммунизации во время беременности главным образом обусловлены в антенатальном периоде развитием у плода тяжелого анемического синдрома, а в постнатальной жизни – гипербилирубинемией, сопровождающейся поражением центральной нервной системы с последующей полиорганной недостаточностью.
Литература к главе I
1. Акушерство: национальное руководство / ред. Э. К. Айламазян, В. И. Кулаков, В. Е. Радзинский, Г. М. Савельева. – М.: ГЭОТАР-Медиа, 2007.– 1200 с.
2. Акушерство: учебник для медицинских вузов / Э. К. Айламазян [и др.].– 6-е изд. – СПб.: Спецлит, 2007. – 528 с.
3. Минеева Н. В. Группы крови человека. Основы иммуногематологии. – СПб., 2004. – 188 с.
4. Сидельникова В. М., Антонов А. Г. Гемолитическая болезнь плода и новорожденного. – М.: Триада-Х, 2004.– 191 с.
5. Analysis of immunoglobullin class, IgG subclass and titre of HPA-la antibodies in alloimmunized mothers giving birth to babies with or without neonatal alloimmune thrombocytopenia / Proulx C. [et al.] // Br. J. Haematology. – 1994.– Vol. 87.– P. 813–817.
6. Anti-D concentration in mother and child in haemolytic disease of the newborn / Hughes-Jones N. C. [et al.] // Vox Sanguinis. – 1971.—Vol. 21.—P. 135–140.
7. Anti-D concentrations in fetal and maternal serum and amniotic fluid in rhesus allo-immunised pregnaneies / Economides D. L. [et al.] // Br. J. of Obstet. and Gynaecol. – 1993. – Vol. 100. – P. 923–926.
8. Antigen topography is critical for interaction of IgG2 anti-red-cell antibodies with Fey receptors / KumpelB.M. [et al.] // Br. J. of Haematology. – 1996. —Vol. 94.—P. 175–183.
9. Billington W.D. The normal fetomaternal immune relationship // Bailliere’s Clinics Obstet. Gynaecol. – 1992.– Vol. 6.—P. 417–438.
10. Blockade of clearance of immune complexes by an anti-FcR monoclonal antibody / Clarkson S. B. [et al.] // J. Experim. Medicine. – 1986. – Vol. 164. – P. 474–489
11. Clinical significance of IgG Fc receptors and FcyR-directed immunotherapies / Deo Y. M. [et al.] // Immunology Today. – 1997. – Vol. 18. —P. 127–135.
12. Correlation of serological, quantitative and cell-mediated functional assays of maternal alloantibodies with the severity of haemolytic disease of the newborn / Hadley A. G. [et al.] // Br. J. of Haematology. – 1991. —Vol. 77.—P. 221–228.
13. Daniels G.L., Hadley A. G., Green C.A. Fetal anaemia due to anti-K may result from immune destruction of early eryth-roid progenitors // Transfusion Medicine. – 1999.– Vol. 9, suppl. 1.– P. 16.
14. Davenport R. D., Kunkel S. L. IgG receptor roles in red cell binding to monocytes and macrophages // Transfusion. – 1994.– Vol. 34.—P. 795.
15. Denomme GRyan G., Fernandes B. The FcyRIIa-Hisl31 allotype is overexpressed in infants with ABO hemolytic disease of the newborn // Blood. – 1997. – Vol. 90. – P. 472–473.
16. Detection of fetal erythrocytes in maternal blood postpartum with the fluorescence-activated cell sorter / Medearis A. L. [et al.] // Am. J. of Obstet. Gynecol. – 1984.—Vol. 148.—P. 290–295.
17. Dooren M.C., Engelfriet C. P. Protection against RhD-haemolytic disease of the newborn by a diminished transport of maternal IgG to the fetus // Vox Sanguinis. – 1993.– Vol. 65.—P. 59–61.
18. Erythropoietic suppression in fetal anaemia because of Kell alloimmunization / Vaughan J.L. [et al.] // Am. J. Obstet. Gynecol. – 1994.—Vol. 171. —P. 247-52.
19. Galactosylation of human IgG monoclonal anti-D produced by EBV-transformed B-lymphoblastoid lines is dependent on culture method and affects Fey receptor-mediated functional activity / Kumpel В. M. [et al.] // Human Antibodies and Hy-bridomas. – 1994. – Vol. 5.– P. 143–151.
20. Ghetie V., Ward E. S. FcRn, the MHC class I-related receptor that is more than an IgG transporter // Immunology Today. – 1997.—Vol. 18.—P. 592–598.
21. Hilden J.O., Gottvall I, Lindblom B. HLA phenotypes and severe Rh (D) immunization // Tissue Antigens. – 1995.– Vol. 46.—P. 3131–3315.
22. HulettM. О., Hogarth P.M. Molecular basis of Fc receptor function // Advances Immunology. – 1994.– Vol. 57.– P. 1–127.
23. Human IgG subclasses in maternal and fetal serum / Morell A. [et al.] // Vox Sanguinis. – 1971. – Vol. 21. – P. 481–492.
24. Inhibition of the monocyte chemiluminescent response to anti-D sensitized red cells by Fey receptor I-blocking antibodies which ameliorate the severity of haemolytic disease of the newborn / Shepard S.L. [et al.] // Vox Sanguinis. – 1996.– Vol. 70.—P. 157–163.
25. Jefferis R., Lund J., Pound J.D. IgG-Fc-mediated effector functions, molecular definition of interaction sites for effector ligands and the role of glycosylation // Immunology Reviews. – 1998. —Vol. 163.—P. 59–76.
26. Kohler P. F., Farr R. S. Elevation of cord over maternal IgG immunoglobulin: evidence for an active placental IgG transport // Nature. – 1966. – Vol. 210.– P. 1070–1071.
27. Kurlander R. J. Blockade of Fc receptor-mediated binding to U-937 cells by murine monoclonal antibodies directed against a variety of surface antigens // J. of Immunology. – 1983.– Vol. 131.—P. 140–147.
28. Lubenko A., Contreras М., Rodeck С. H. Transplacental IgG subclass concentrations in pregnancies at risk of haemolytic disease of the newborn // Vox Sanguinis. – 1994. – Vol. 67. – P. 291–298.
29. Male fetuses are particularly affected by maternal alloimmunization to D antigen / Ulm B. [et al.] // Transfusion. – 1999. —Vol. 39. —P. 169–173.
30. Mapping the site on human IgG for binding of the MHC class I-related receptor, FcRn / Kim J-K. [et al.] // Eur. J. Immunology. – 1999.– Vol. 29.– P. 2819–2825.
31. Maternal antibodies against fetal blood group antigens A or B, lytic activity of IgG subclasses in monocyte-driven cytotoxicity and correlation with ABO haemolytic disease of the newborn / Brouwers H. A. [et al.] // Br. J. Haematology. – 1988. – Vol. 70.—P. 465–469.
32. Misleading results in the determination of haemolytic disease of the newborn using antibody titration and ADCC in a woman with anti-Lub / Novotny V. M. J. [et al.] // Vox Sanguinis. – 1992. —Vol. 62. —P. 49–52.
33. Molecular basis for a polymorphism of human FcyRII (CD32) / Warmerdam P. A. M. [et al.] // J. Experim. Medicine. – 1990. —Vol. 172.—P. 19–25.
34. Nance S. J., Arndt P. A., Garratty G. Correlation of IgG subclass with the severity of hemolytic disease of the newborn // Transfusion. – 1990.– Vol. 30.– P. 381–382.
35. Parinaud J. [et al.] IgG subclasses and Gm allotypes of anti-D antibodies during pregnancy: correlation with the gravity of fetal disease // Am. J. Obstet. and Gynecol. – 1985.– Vol. 151. —P. 1111–1115.
36. Pollock J. М., Bowman J.M. Anti-Rh (D) IgG subclasses and severity of Rh hemolytic disease of the newborn // Vox Sanguinis. – 1990.—Vol. 59.—P. 176–179.
37. Protection against immune haemolytic disease of newborn infants by maternal monocyte-reactive IgG alloantibodies (anti HLA-DR) / DoorenM. C. [et al.] // Lancet. – 1992.– Vol. 339.—P. 1067–1070.
38. Ramsey G., Sherman L. A. Anti-D in pregnancy: value of antibody titres and effect of fetal gender // Transfusion. – 1999. – Vol. 39.—P. 105.
39. Ravetch J. V, Perussia B. Alternative membrane forms of FcyRIII (CDI6) on human NK cells and neutrophils, cell-type specific expression of two genes which differ in single nucleotide substitutions // J. Experim. Medicine. – 1989.– Vol. 170.—P. 481–491.
40. Renkonen К. O., Seppala M. The sex of the sensitizing Rh-positive child // Annals of Medicine. – 1962.– Vol. 40.– P. 108–118.
41. Renkonen К. O., Timonen S. Factors influencing the immunization of Rh-negative mothers // J. of Medical Genetics. – 1967.—Vol. 4.—P. 166–176.
42. Rhesus Du incompatibility in a newborn with high levels of anti-D and a benign clinical course / Dias R. [et al.] // Vox Sanguinis. – 1986. – Vol. 50. – R 52–53.
43. Rochna E., Hughes-Jones N. C. The use of purified 1251-la-belled anti-y globullin in the determination of the number of D antigen sites on red cells of different phenotypes // Vox Sanguinis. – 1965.—Vol. 10.—P. 675–686.
44. Selective placental transport of maternal IgG to the fetus / Williams P. J. [et al.] // Placenta. – 1995. – Vol. 16. – P. 749–756.
45. Shepard S. L., Hadley A. G. Monocyte-bound monoclonal antibodies inhibit the FcyRI-mediated phagocytosis of red cells: the efficiency and mechanism of inhibition are determined by the nature of the antigen // Immunology. – 1997.—Vol. 90.– P. 314–322.
46. Skidmore I., Hadley A. G. The effect of specificity on the functional activity of red cell-bound blood group antibodies // Transfusion Medicine. – 1996. – Vol. 6, suppl. 2.– P. 26.
47. Story C.M., Mikulska J. E., Simister N.E. A major histocompatibility complex class I-like Fc receptor cloned from human placenta: possible role in transfer of immunoglobulin G from mother to fetus // J. Experim. Medicine. – 1994.– Vol. 180.—P. 2377–2381.
48. TaslimiM.M. [et al.] Immunoglobulin G subclasses and isoimmunized pregnancy outcome // Am. J. of Obstet. and Gynecol. – 1986.—Vol. 154. —P. 1327–1332.
49. The alloreactive and autoreactive T helper cytokine responses to the RhD protein differ / Hall A. M. [et al.] // Transfusion Medicine. – 1999. – Vol. 9, suppl. 1. – P. 17.
50. The expression of IgG Fc receptors on circulating leucocytes in the fetus and new-born / Mawas P. [et al.] // Transfusion Medicine. – 1994.– Vol. 4.– P. 25–33.
51. Urbaniak S.J., Greiss M.A. ADCC (К-cell) lysis of human erythrocytes sensitized with Rhesus alloantibodies. III. Comparison of IgG anti-D agglutinating and lytic (ADCC) activity and the role of IgG subclasses // Br. J. of Haematology. – 1980. – Vol. 46. – P. 447–453.
52. Vietor Н.Е., Kanhai Brand A. Induction of additional red cell alloantibodies after intrauterine transfusions // Transfusion. – 1994.—Vol. 34. —P. 970–974.
53. Woodrow J. C. Rh immunisation and its prevention // Ser. Haematol. -1970. —Vol. 3, N 3.– P. 1–151.
54. Woodrow J. C., Finn R. Transplacental haemorrhage // Br. J. Haematology. – 1966. —Vol. 12.—P. 297–307.
55. Wu S., Maslanka K, Gorski J. An integrin polymorphism that defines reactivity with alloantibodies generates an anchor for MHC class II peptide binding: a model for unindirectional al-loimmune responses // J. Immunology. – 1997. – Vol. 158. – P. 3221–3226.
56. Zupanska B. [et al.] Serological and immunological characteristics of maternal anti-Rh (D) antibodies in predicting the severity of haemolytic disease of the newborn // Vox Sanguinis. – 1989. —Vol. 56.—P. 247–253.
57. Bux J. Nomenclature of granulocyte alloantigens // Transfusion. – 1999. —Vol. 39.—P. 662–663.
58. KumpelB.M., Hadley A. G. Functional interactions of red cells sensitised by IgGl and IgG3 human monoclonal anti-D with enzyme-modified human monocytes and FcR-bearing cell lines // Molecular Immunology. – 1990.– Vol. 27.– P. 247–256.
59. Murray S., Knox E. G., Walker W. Rhesus haemolytic disease of the newborn and the ABO groups // Vox Sanguinis. – 1965.—Vol. 10. —P. 6-31.
60. Mononuclear phagocyte assays, AutoAnalyzer quantitation and IgG subclasses of maternal anti-RhD in the prediction of the severity of haemolytic disease in the fetus before 32 weeks gestation / Garner S. F. [et al.] // Br. J. of Haematology. – 1992.—Vol. 80.—P. 97-101.
2. Акушерство: учебник для медицинских вузов / Э. К. Айламазян [и др.].– 6-е изд. – СПб.: Спецлит, 2007. – 528 с.
3. Минеева Н. В. Группы крови человека. Основы иммуногематологии. – СПб., 2004. – 188 с.
4. Сидельникова В. М., Антонов А. Г. Гемолитическая болезнь плода и новорожденного. – М.: Триада-Х, 2004.– 191 с.
5. Analysis of immunoglobullin class, IgG subclass and titre of HPA-la antibodies in alloimmunized mothers giving birth to babies with or without neonatal alloimmune thrombocytopenia / Proulx C. [et al.] // Br. J. Haematology. – 1994.– Vol. 87.– P. 813–817.
6. Anti-D concentration in mother and child in haemolytic disease of the newborn / Hughes-Jones N. C. [et al.] // Vox Sanguinis. – 1971.—Vol. 21.—P. 135–140.
7. Anti-D concentrations in fetal and maternal serum and amniotic fluid in rhesus allo-immunised pregnaneies / Economides D. L. [et al.] // Br. J. of Obstet. and Gynaecol. – 1993. – Vol. 100. – P. 923–926.
8. Antigen topography is critical for interaction of IgG2 anti-red-cell antibodies with Fey receptors / KumpelB.M. [et al.] // Br. J. of Haematology. – 1996. —Vol. 94.—P. 175–183.
9. Billington W.D. The normal fetomaternal immune relationship // Bailliere’s Clinics Obstet. Gynaecol. – 1992.– Vol. 6.—P. 417–438.
10. Blockade of clearance of immune complexes by an anti-FcR monoclonal antibody / Clarkson S. B. [et al.] // J. Experim. Medicine. – 1986. – Vol. 164. – P. 474–489
11. Clinical significance of IgG Fc receptors and FcyR-directed immunotherapies / Deo Y. M. [et al.] // Immunology Today. – 1997. – Vol. 18. —P. 127–135.
12. Correlation of serological, quantitative and cell-mediated functional assays of maternal alloantibodies with the severity of haemolytic disease of the newborn / Hadley A. G. [et al.] // Br. J. of Haematology. – 1991. —Vol. 77.—P. 221–228.
13. Daniels G.L., Hadley A. G., Green C.A. Fetal anaemia due to anti-K may result from immune destruction of early eryth-roid progenitors // Transfusion Medicine. – 1999.– Vol. 9, suppl. 1.– P. 16.
14. Davenport R. D., Kunkel S. L. IgG receptor roles in red cell binding to monocytes and macrophages // Transfusion. – 1994.– Vol. 34.—P. 795.
15. Denomme GRyan G., Fernandes B. The FcyRIIa-Hisl31 allotype is overexpressed in infants with ABO hemolytic disease of the newborn // Blood. – 1997. – Vol. 90. – P. 472–473.
16. Detection of fetal erythrocytes in maternal blood postpartum with the fluorescence-activated cell sorter / Medearis A. L. [et al.] // Am. J. of Obstet. Gynecol. – 1984.—Vol. 148.—P. 290–295.
17. Dooren M.C., Engelfriet C. P. Protection against RhD-haemolytic disease of the newborn by a diminished transport of maternal IgG to the fetus // Vox Sanguinis. – 1993.– Vol. 65.—P. 59–61.
18. Erythropoietic suppression in fetal anaemia because of Kell alloimmunization / Vaughan J.L. [et al.] // Am. J. Obstet. Gynecol. – 1994.—Vol. 171. —P. 247-52.
19. Galactosylation of human IgG monoclonal anti-D produced by EBV-transformed B-lymphoblastoid lines is dependent on culture method and affects Fey receptor-mediated functional activity / Kumpel В. M. [et al.] // Human Antibodies and Hy-bridomas. – 1994. – Vol. 5.– P. 143–151.
20. Ghetie V., Ward E. S. FcRn, the MHC class I-related receptor that is more than an IgG transporter // Immunology Today. – 1997.—Vol. 18.—P. 592–598.
21. Hilden J.O., Gottvall I, Lindblom B. HLA phenotypes and severe Rh (D) immunization // Tissue Antigens. – 1995.– Vol. 46.—P. 3131–3315.
22. HulettM. О., Hogarth P.M. Molecular basis of Fc receptor function // Advances Immunology. – 1994.– Vol. 57.– P. 1–127.
23. Human IgG subclasses in maternal and fetal serum / Morell A. [et al.] // Vox Sanguinis. – 1971. – Vol. 21. – P. 481–492.
24. Inhibition of the monocyte chemiluminescent response to anti-D sensitized red cells by Fey receptor I-blocking antibodies which ameliorate the severity of haemolytic disease of the newborn / Shepard S.L. [et al.] // Vox Sanguinis. – 1996.– Vol. 70.—P. 157–163.
25. Jefferis R., Lund J., Pound J.D. IgG-Fc-mediated effector functions, molecular definition of interaction sites for effector ligands and the role of glycosylation // Immunology Reviews. – 1998. —Vol. 163.—P. 59–76.
26. Kohler P. F., Farr R. S. Elevation of cord over maternal IgG immunoglobulin: evidence for an active placental IgG transport // Nature. – 1966. – Vol. 210.– P. 1070–1071.
27. Kurlander R. J. Blockade of Fc receptor-mediated binding to U-937 cells by murine monoclonal antibodies directed against a variety of surface antigens // J. of Immunology. – 1983.– Vol. 131.—P. 140–147.
28. Lubenko A., Contreras М., Rodeck С. H. Transplacental IgG subclass concentrations in pregnancies at risk of haemolytic disease of the newborn // Vox Sanguinis. – 1994. – Vol. 67. – P. 291–298.
29. Male fetuses are particularly affected by maternal alloimmunization to D antigen / Ulm B. [et al.] // Transfusion. – 1999. —Vol. 39. —P. 169–173.
30. Mapping the site on human IgG for binding of the MHC class I-related receptor, FcRn / Kim J-K. [et al.] // Eur. J. Immunology. – 1999.– Vol. 29.– P. 2819–2825.
31. Maternal antibodies against fetal blood group antigens A or B, lytic activity of IgG subclasses in monocyte-driven cytotoxicity and correlation with ABO haemolytic disease of the newborn / Brouwers H. A. [et al.] // Br. J. Haematology. – 1988. – Vol. 70.—P. 465–469.
32. Misleading results in the determination of haemolytic disease of the newborn using antibody titration and ADCC in a woman with anti-Lub / Novotny V. M. J. [et al.] // Vox Sanguinis. – 1992. —Vol. 62. —P. 49–52.
33. Molecular basis for a polymorphism of human FcyRII (CD32) / Warmerdam P. A. M. [et al.] // J. Experim. Medicine. – 1990. —Vol. 172.—P. 19–25.
34. Nance S. J., Arndt P. A., Garratty G. Correlation of IgG subclass with the severity of hemolytic disease of the newborn // Transfusion. – 1990.– Vol. 30.– P. 381–382.
35. Parinaud J. [et al.] IgG subclasses and Gm allotypes of anti-D antibodies during pregnancy: correlation with the gravity of fetal disease // Am. J. Obstet. and Gynecol. – 1985.– Vol. 151. —P. 1111–1115.
36. Pollock J. М., Bowman J.M. Anti-Rh (D) IgG subclasses and severity of Rh hemolytic disease of the newborn // Vox Sanguinis. – 1990.—Vol. 59.—P. 176–179.
37. Protection against immune haemolytic disease of newborn infants by maternal monocyte-reactive IgG alloantibodies (anti HLA-DR) / DoorenM. C. [et al.] // Lancet. – 1992.– Vol. 339.—P. 1067–1070.
38. Ramsey G., Sherman L. A. Anti-D in pregnancy: value of antibody titres and effect of fetal gender // Transfusion. – 1999. – Vol. 39.—P. 105.
39. Ravetch J. V, Perussia B. Alternative membrane forms of FcyRIII (CDI6) on human NK cells and neutrophils, cell-type specific expression of two genes which differ in single nucleotide substitutions // J. Experim. Medicine. – 1989.– Vol. 170.—P. 481–491.
40. Renkonen К. O., Seppala M. The sex of the sensitizing Rh-positive child // Annals of Medicine. – 1962.– Vol. 40.– P. 108–118.
41. Renkonen К. O., Timonen S. Factors influencing the immunization of Rh-negative mothers // J. of Medical Genetics. – 1967.—Vol. 4.—P. 166–176.
42. Rhesus Du incompatibility in a newborn with high levels of anti-D and a benign clinical course / Dias R. [et al.] // Vox Sanguinis. – 1986. – Vol. 50. – R 52–53.
43. Rochna E., Hughes-Jones N. C. The use of purified 1251-la-belled anti-y globullin in the determination of the number of D antigen sites on red cells of different phenotypes // Vox Sanguinis. – 1965.—Vol. 10.—P. 675–686.
44. Selective placental transport of maternal IgG to the fetus / Williams P. J. [et al.] // Placenta. – 1995. – Vol. 16. – P. 749–756.
45. Shepard S. L., Hadley A. G. Monocyte-bound monoclonal antibodies inhibit the FcyRI-mediated phagocytosis of red cells: the efficiency and mechanism of inhibition are determined by the nature of the antigen // Immunology. – 1997.—Vol. 90.– P. 314–322.
46. Skidmore I., Hadley A. G. The effect of specificity on the functional activity of red cell-bound blood group antibodies // Transfusion Medicine. – 1996. – Vol. 6, suppl. 2.– P. 26.
47. Story C.M., Mikulska J. E., Simister N.E. A major histocompatibility complex class I-like Fc receptor cloned from human placenta: possible role in transfer of immunoglobulin G from mother to fetus // J. Experim. Medicine. – 1994.– Vol. 180.—P. 2377–2381.
48. TaslimiM.M. [et al.] Immunoglobulin G subclasses and isoimmunized pregnancy outcome // Am. J. of Obstet. and Gynecol. – 1986.—Vol. 154. —P. 1327–1332.
49. The alloreactive and autoreactive T helper cytokine responses to the RhD protein differ / Hall A. M. [et al.] // Transfusion Medicine. – 1999. – Vol. 9, suppl. 1. – P. 17.
50. The expression of IgG Fc receptors on circulating leucocytes in the fetus and new-born / Mawas P. [et al.] // Transfusion Medicine. – 1994.– Vol. 4.– P. 25–33.
51. Urbaniak S.J., Greiss M.A. ADCC (К-cell) lysis of human erythrocytes sensitized with Rhesus alloantibodies. III. Comparison of IgG anti-D agglutinating and lytic (ADCC) activity and the role of IgG subclasses // Br. J. of Haematology. – 1980. – Vol. 46. – P. 447–453.
52. Vietor Н.Е., Kanhai Brand A. Induction of additional red cell alloantibodies after intrauterine transfusions // Transfusion. – 1994.—Vol. 34. —P. 970–974.
53. Woodrow J. C. Rh immunisation and its prevention // Ser. Haematol. -1970. —Vol. 3, N 3.– P. 1–151.
54. Woodrow J. C., Finn R. Transplacental haemorrhage // Br. J. Haematology. – 1966. —Vol. 12.—P. 297–307.
55. Wu S., Maslanka K, Gorski J. An integrin polymorphism that defines reactivity with alloantibodies generates an anchor for MHC class II peptide binding: a model for unindirectional al-loimmune responses // J. Immunology. – 1997. – Vol. 158. – P. 3221–3226.
56. Zupanska B. [et al.] Serological and immunological characteristics of maternal anti-Rh (D) antibodies in predicting the severity of haemolytic disease of the newborn // Vox Sanguinis. – 1989. —Vol. 56.—P. 247–253.
57. Bux J. Nomenclature of granulocyte alloantigens // Transfusion. – 1999. —Vol. 39.—P. 662–663.
58. KumpelB.M., Hadley A. G. Functional interactions of red cells sensitised by IgGl and IgG3 human monoclonal anti-D with enzyme-modified human monocytes and FcR-bearing cell lines // Molecular Immunology. – 1990.– Vol. 27.– P. 247–256.
59. Murray S., Knox E. G., Walker W. Rhesus haemolytic disease of the newborn and the ABO groups // Vox Sanguinis. – 1965.—Vol. 10. —P. 6-31.
60. Mononuclear phagocyte assays, AutoAnalyzer quantitation and IgG subclasses of maternal anti-RhD in the prediction of the severity of haemolytic disease in the fetus before 32 weeks gestation / Garner S. F. [et al.] // Br. J. of Haematology. – 1992.—Vol. 80.—P. 97-101.
Глава II Антенатальная диагностика аллоиммунных эритропений
Как известно, у беременных группы риска по развитию изоиммунизации иммунопрофилактика до настоящего времени, к сожалению, не является, с одной стороны, всеобщей, а, с другой стороны, всеобъемлющей, поскольку она предотвращает развитие только наиболее распространенной анти-D-изоиммунизации. Поэтому, несмотря на снижение в мире частоты изоиммунизации
Конец бесплатного ознакомительного фрагмента