Страница:
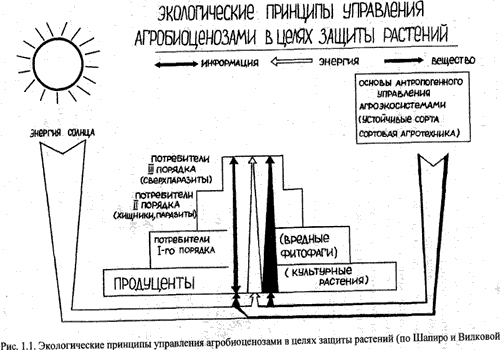
Все многообразие экологических взаимоотношений между организмами в биоценозах объединяется в три формы связей – вещественные, энергетические и информационные, т. е. по цепям питания передается запас вещества, энергии и информации. Передача вещества и энергии осуществляется в системе однонаправленно: от продуцента, т. е. растения, к потребителю, т. е. фитофагу, и определяет уровень жизнедеятельности вредителей. Взаимодействие по информационному каналу осуществляется по принципу обратной связи (рис. 1).
Большинство вредителей отличаются большой подвижностью и активностью, что позволяет им получать огромный объем разнообразной информации от растения. Контакты их с растениями в онтогенезе не постоянны во времени и для свободно живущих ограничиваются в основном актом еды и процессом откладки яиц. Хорошо развитые рецепторы обеспечивают фитофагам широкие возможности выбора оптимальных кормовых растений, их органов и тканей. Относительно малые размеры тела при общей высокой организации, подвижности в сочетании с большим воспроизводительным потенциалом, высокими темпами роста и скоростями развития и другие специфические особенности делают членистоногих вредителей весьма требовательными к пластическому и энергетическому обеспечению. Эта особенность превращает их в одно из главных звеньев, преобразующих вещество, энергию и информацию в экосистемах, как потребителей и вторичной биологической продукции.
Членистоногие благодаря совершенным органам чувств способны воспринимать разнообразную информацию о свойствах растений: внешнем строении, окраске, физиологическом состоянии, содержании различных по строению химических веществ и т. д. Так, насекомые хорошо различают растения по форме и уровню содержания физиологически активных веществ, в том числе фитогормонов, углеводов, белков, липидов. Положительный энергетический и пластический баланс животных зависит не только от общего содержания в пищевом субстрате нутриентов, но и от их молекулярного строения, оказывающего влияние на скорость, легкость и степень их переваривания.
1.8. Формы пищевой специализации вредителей
Большинство вредителей отличаются большой подвижностью и активностью, что позволяет им получать огромный объем разнообразной информации от растения. Контакты их с растениями в онтогенезе не постоянны во времени и для свободно живущих ограничиваются в основном актом еды и процессом откладки яиц. Хорошо развитые рецепторы обеспечивают фитофагам широкие возможности выбора оптимальных кормовых растений, их органов и тканей. Относительно малые размеры тела при общей высокой организации, подвижности в сочетании с большим воспроизводительным потенциалом, высокими темпами роста и скоростями развития и другие специфические особенности делают членистоногих вредителей весьма требовательными к пластическому и энергетическому обеспечению. Эта особенность превращает их в одно из главных звеньев, преобразующих вещество, энергию и информацию в экосистемах, как потребителей и вторичной биологической продукции.
Рис. 1. Экологические принципы управления агробиоценозами в целях защиты растений (по Шапиро и Вилковой)
Членистоногие благодаря совершенным органам чувств способны воспринимать разнообразную информацию о свойствах растений: внешнем строении, окраске, физиологическом состоянии, содержании различных по строению химических веществ и т. д. Так, насекомые хорошо различают растения по форме и уровню содержания физиологически активных веществ, в том числе фитогормонов, углеводов, белков, липидов. Положительный энергетический и пластический баланс животных зависит не только от общего содержания в пищевом субстрате нутриентов, но и от их молекулярного строения, оказывающего влияние на скорость, легкость и степень их переваривания.
1.8. Формы пищевой специализации вредителей
Рассмотрение взаимоприспособлений фитофагов и их кормовых растений в связи с системой иммуногенетической защиты растений вызывает необходимость анализа пищевой специализации вредителей. Еще выдающийся советский ученый Н.И. Вавилов (1919, 1964) рассматривал явления иммунитета растений как синоним понятия «пищевая специализация паразита». В развитие идей Н.И. Вавилова современные иммунологи в качестве первоочередных задач в решении проблемы фитоиммунитета выдвигают изучение специализации патогенных организмов и их адаптации к антибиотическому воздействию растений, что, в свою очередь, вызывает необходимость изучения эволюции паразитизма патогенов (Вердеревский, 1966; Дунин, 1966, и др.).
Круг явлений, определяющих выбор или отвергание насекомыми растений для питания и откладки яиц, степень благоприятности корма, его усвояемость и т. д., дает представление, с одной стороны, об особенностях пищевой специализации фитофагов и, с другой – об основных факторах иммунитета.
Одним из наиболее сложных для решения и широко обсуждаемых в специальной мировой литературе является вопрос о механизмах, определяющих пищевую специализацию фитофагов.
Своеобразие современных систем «кормовое растение (повреждаемая культура) – фитофаг» определяется длительностью сопряженной эволюции организмов, обусловившей формирование адаптивных черт у партнеров.
Известно, что организмы в биоценозах связаны как единством жизненного субстрата, так и единством эволюционного процесса.
В условиях трофической эволюции основное направление отбора способствует развитию стабилизирующих регуляторных механизмов продуктивности популяций, населяющих биоценоз, и усиливает взаимосвязи между ними. В ходе сопряженной эволюции растений и их потребителей-фитофагов действие отбора было направлено на сохранение и совершенствование связей, повышающих устойчивость и прочность этой экологической системы. С экологической точки зрения специализацию фитофагов по отношению к кормовым растениям можно рассматривать как один из механизмов поддержания и сохранения устойчивости рассматриваемой системы.
Живая система всегда оказывается приспособленной к условиям существования, что считают наиболее характерной чертой жизни. При этом любой адаптационный процесс, по свидетельству Г. Селье (1960) и многих других, по своей сути есть процесс энергетический, не отделимый от других энергетических процессов организма и требующий постоянного притока энергии извне.
По мере повышения уровня организации организмов наряду с совершенствованием использования энергии повышается их зависимость от источников энергии. У гетеротрофов пища служит единственным источником пластического и энергетического обеспечения. Насекомые благодаря своим специфическим особенностям могут быть отнесены к организмам с очень высоким уровнем энергетических затрат.
Видовые адаптации, по свидетельству С.С. Шварца (1969), весьма разнообразны и необозримо совершенны, что определяется не только их функциональной активностью, но и энергетической стоимостью. Отсюда становится очевидным значение экономизации обмена и совершенствования способности к накоплению энергетических резервов как важнейших для адаптации. Известно, что в норме функциональные механизмы организма действуют с минимальной затратой энергии.
Экономия энергии, даже, казалось бы, самая ничтожная, может иметь решающее значение в жизнедеятельности и эволюции организмов, в том числе и насекомых. Особенно важным, по мнению С.С. Шварца (1969), следует считать соотношение энергетических потребностей животного с затратами энергии на добывание корма, что во многих случаях предопределяет пути микроэволюционных преобразований. В сопряженной эволюции с консументами основным направлением приспособительной изменчивости у растения-хозяина было развитие защитных механизмов от многочисленных врагов, в том числе и насекомых. С этим, очевидно, связана часто проявляющаяся общность факторов резистентности по отношению ко многим повреждающим агентам. В то же время адаптации у фитофагов оказались направленными на приспособление к наиболее рациональному использованию материально-энергетических ресурсов, поставляемых кормовым растением. Эта тенденция выразилась у фитофагов в многочисленных приспособлениях, затронувших различные уровни и организации. Это привело к становлению гостальной, топической и онтогенетической специализации. У насекомых, в свою очередь, эволюционировали и механизмы защиты от отрицательных воздействий хозяина.
Применительно к фитофагам различают несколько категорий пищевой специализации:
1) гостальную специализацию, приводящую к приобретению способности нормально существовать и развиваться лишь на растениях-хозяевах, относящихся к определенным систематическим группам;
2) топическую специализацию, при которой виды оказываются способными развиваться лишь при условии особой локализации на хозяевах;
3) онтогенетическую специализацию, приводящую к возникновению способности развиваться на органах хозяина, находящихся в определенном для каждого вида фитофага возрасте и морфофизиологическом состоянии.
При этом гостальная специфичность выступает как основная категория специализации, производными от которой являются две другие взаимосвязанные категории: топическая и онтогенетическая,
С точки зрения понимания природы устойчивости растений большой интерес представляет деление гостальной специфичности в соответствии с различиями таксономического ранга хозяев на:
1) специфичность крупных таксонов потребителей к крупным таксонам хозяев;
2) специфичность семейств или основных надсемейственньгх таксонов по отношению к роду, семейству или основным надсемейственным таксонам хозяев;
3) специфичность к виду и надвидовым таксонам хозяев;
4) специфичность вида-паразита или симбионта к внутривидовым группам, видам или родам хозяев (Слепян, 1973). Заслуживает внимания при рассмотрении гостальной специализации наличие существенных различий между так называемой реальной и потенциальной специфичностью, отмеченной у паразитов по отношению к их хозяевам (Киршенблат, 1941; Догель, 1962).
Различия между растениями, относящимися к группе потенциально возможных для питания и растениями оптимальными, зависят от следующих причин:
1) форм поведения, в которых выявляются избирательные отношения насекомого к определенным растениям, обусловленные реакцией на тот или иной признак последних;
2) влияния растения на насекомое при питании. Например, клопы (вредная черепашка) меняют место питания в зависимости от этапа органогенеза растений (рис. 2). Сочетание этих условий и определяет широкий спектр внутривидового разнообразия градаций устойчивости растений к вредителям.
Общепринятое деление фитофагов по широте пищевых связей на поли-, олиго– и монофагов, по существу, не отражает существования в каждой группе предпочтения определенного круга растений, в том числе и внутривидовых форм, которыми изобилуют культурные виды растений. Среди фитофагов наиболее широко представлены поли– и олигофага, включающие виды с более широкой или узкой специализацией. Доместикация и селекция растений привели к широкому их полиморфизму, что, в свою очередь, углубило процессы пищевой специализации фитофагов. Часто генетическая природа этих форм бывает гораздо сложнее, чем у диких видов.
Необходимость вычленения специфичности фитофагов к внутривидовым формам растений (сортам, гибридам, линиям) совершенно очевидна. Этот уровень пищевой специализации определяется факторами сортовой устойчивости растений. Н.И. Вавилов (1939) придавал большое значение наряду с родовым и видовым иммунитетом разнообразию представленности иммунитета полиморфных видов – от практически полной их резистентности до сильной восприимчивости.
Таким образом, напряжение иммунитета к вредителям на посевах культурных растений поддерживается иммуногенной защитой разных таксономических категорий растений: семейств, подсемейств, триб, родов, видов и внутривидовых форм. Выделение этих форм иммунитета растений базируется на соответствующих уровнях гостальной специализации фитофагов. Устойчивость растений к поли-, олиго– и монофагам может определяться каждой из названных форм иммунитета или их сочетанием. Характерно, что для полифагов большое значение имеют факторы, определяющие внутривидовую (сортовую) устойчивость. Вместе с тем на первых этапах изучения природы иммунитета растений к вредителям сложилось мнение о трудности нахождением устойчивых к многоядным вредителям форм растений и невозможности их создания (Щеголев, 1938). Однако, как показывает мировой опыт, наиболее значительные успехи были достигнуты в селекции устойчивых к полифагам и широким олигофагам сортов сельскохозяйственных культур.
В отношении гостальной специфичности, особенно видов с широким кругом кормовых растений, можно выделить три уровня специализации:
1) специализация, ограничивающая круг растений, возможных для питания в пределах относительно крупных их таксономических групп;
2) специализация, выражающаяся в предпочтении определенных форм растений из числа возможных;
3) специализация, выражающаяся в предпочтении внутривидовых форм полиморфных видов растений.
Поиски причин подобной специализации, предпринятые многими исследователями, породили различные гипотезы, трактующие избирательность насекомых по отношению к растениям с самых разных позиций.
Топическая и онтогенетическая специализация фитофагов выступают для каждого вида растений и животных как неизбежность биологического прогресса, в связи с чем становится очевидным, что избирательное отношение фитофагов к видам и внутривидовым формам кормовых растений следует рассматривать лишь как ступень пищевой специализации насекомых.
Рассмотрение гостальной специализации фитофагов создает некоторую иллюзию их широких возможностей в использовании разнообразных пищевых субстратов, представляемых различными органами растений. Однако из огромного числа фитофагов лишь ограниченное число видов способно питаться на всех без исключения органах растений. Среди фитофагов четко выделяются строго специализированные группы: филофаги, ксилофаги, ризофаги, антофаги, карпофаги др. Становление топической специализации, помимо органотропности – приуроченности к определенным органам и системам органов, проявляется на тканевом (гистротропность) и клеточном (цитотропность) уровнях, когда в пределах каждого органа растения фитофаги предпочитают питаться определенными тканями и их структурами.
Можно полагать, что обилие видов фитофагов, приуроченных к вегетативным органам растений, отражает особенности прогрессивной эволюции покрытосеменных. Ароморфоз цветковых растений, по мнению ряда исследователей, базировался в первую очередь на постоянных, т. е. вегетативных органах (Крыжановский, 1958; Яценко-Хмелевский, 1958; Попов, 1963; Проханов, 1965; Тихомиров, 1965, и др.). Очевидно, в связи с этим высокого совершенства достигла и иммуногенная система вегетативной сферы растений. Важные физиологические функции, выполняемые вегетативными органами, и обилие массы имели, как можно полагать, решающее значение для приспособительной эволюции фитофагов. Существенным в становлении специфичности фитофагов следует считать и тот факт, что «основной линией прогрессивной эволюции высших растений является не усовершенствование структур, а убыстрение темпов роста и развития». Это свойство, присущее покрытосеменным, усилило значение ростового барьера как одного из важных элементов иммунной системы растений. В то же время ускорение темпов роста и происходящую в связи с этим легкость регенерации точек роста у покрытосеменных М.Г. Попов (1963) связывает с увеличением и усложнением белковой молекулы. Следствием этих существенных преобразований белка явился синтез самых разнообразных форм побочных продуктов жизнедеятельности растений: алкалоидов, эфирных масел, органических кислот и т. д. Эти соединения, обладая широким спектром антибиотического действия, играют огромную роль в иммуногенной системе растений.
Существенными во взаимосвязях с насекомыми следует признать такие онтогенетические особенности растений, как способность к восстановлению органов благодаря специфическим тканям – первичной и вторичной меристемам – и способность к ослаблению связей с внешней средой или сведение этого взаимодействия к минимуму в отдельные периоды онтогенеза, выраженные физической и физиологической изоляцией.
Экологическое своеобразие системы «насекомое – фитофаг – кормовое растение» определяется необходимостью периодического возобновления взаимосвязей между составляющими ее элементами – растением и насекомым. Особенно яркую выраженность онтогенетической периодичности становления система получила во взаимоотношениях фитофагов с культурными растениями, где в результате ротации культур в севообороте происходят частые смены стаций. В связи с этим результативность взаимодействия растений и их потребителей – фитофагов – в значительной степени зависит от синхронизации развития партнеров.
Надежность и устойчивость взаимосвязей фитофагов с кормовыми растениями обеспечивается согласованностью в пространстве и времени определенных морфофункциональных изменений тех и других организмов (Шапиро, 1962,1985; Вилкова, 1980).
Следствием этого явилось становление у насекомых особых типов сопряженности циклов развития с циклами развития хозяев. Приуроченность к определенным возрастным периодам развития растений, в основе которой лежит топическая специфичность, свойственна всем группам фитофагов. Однако более четко выраженная зависимость от определенных этапов органогенеза кормового растения проявляется у фитофагов, приспособившихся к внутритканевому образу жизни.
Таким образом, в сопряженной эволюции с растением-хозяином формирование приспособительных черт фитофагов проходило на фоне сложных кинетических изменений, происходящих в онтогенезе растений, носящих характер циклических – ростовых и органообразовательных процессов, в основе которых лежат физико-химические превращения энергетических и пластических веществ в растениях. В связи с этим приспособительная эволюция фитофагов была подчинена особенностям морфоанатомической конституции, характеру и темпам формирования и дифференциации различных органов и тканей растений в онтогенезе последних, особенностям синтеза, транспорта и резервирования растениями веществ основного и «вторичного» обменов. В связи с этим можно полагать, что адаптации фитофагов были направлены в сторону морфофизиологических приспособлений, обеспечивающих потребителям относительно гомеостатические условия использования в пищу непрерывно меняющихся в онтогенезе растений форм и уровня содержания пластических и энергетических ресурсов (Шапиро, Вилкова, 1973).
Этот процесс нашел свое выражение в становлении у насекомых строгой приуроченности к использованию лишь определенных органов и тканей, находящихся на тех или иных этапах формирования. Так, для многих видов насекомых хорошо известна приуроченность к питанию осевыми органами, находящимися на определенных этапах онтогенеза. Еще более выражена приуроченность многих видов насекомых к питанию репродуктивными органами лишь на тех или иных этапах органогенеза. Во многих случаях относительно гомеостатические условия питания насекомых обеспечиваются путем приспособления к питанию строго определенными тканями растений. Это особенно ярко проявляется в приспособлении многих видов сосущих насекомых к высасыванию соков из эмбриональных тканей, паренхимы, флоэмы и других специализированных тканей растений. Принцип анализа сопряженности онтогенезов фитофагов и их кормовых растений позволил подойти к пониманию сущности топической и онтогенетической специализации многих видов.
Приобретению фитофагами в процессе эволюции приуроченности к оптимальным кормовым растениям, определенного морфофизиологического состояния, способствовала необходимость преодоления барьеров иммунной системы растений – тканевого (эпидерма, пробка, склеренхима, древесина), осмотического, ростового, органогенетического (большая скорость роста и дифференциации органов и тканей) и метаболического – наличие токсических соединений. Этот процесс нашел выражение в становлении у насекомых строгой приуроченности к использованию определенных органов тканей растений, находящихся на тех или иных этапах формирования.
Круг явлений, определяющих выбор или отвергание насекомыми растений для питания и откладки яиц, степень благоприятности корма, его усвояемость и т. д., дает представление, с одной стороны, об особенностях пищевой специализации фитофагов и, с другой – об основных факторах иммунитета.
Одним из наиболее сложных для решения и широко обсуждаемых в специальной мировой литературе является вопрос о механизмах, определяющих пищевую специализацию фитофагов.
Своеобразие современных систем «кормовое растение (повреждаемая культура) – фитофаг» определяется длительностью сопряженной эволюции организмов, обусловившей формирование адаптивных черт у партнеров.
Известно, что организмы в биоценозах связаны как единством жизненного субстрата, так и единством эволюционного процесса.
В условиях трофической эволюции основное направление отбора способствует развитию стабилизирующих регуляторных механизмов продуктивности популяций, населяющих биоценоз, и усиливает взаимосвязи между ними. В ходе сопряженной эволюции растений и их потребителей-фитофагов действие отбора было направлено на сохранение и совершенствование связей, повышающих устойчивость и прочность этой экологической системы. С экологической точки зрения специализацию фитофагов по отношению к кормовым растениям можно рассматривать как один из механизмов поддержания и сохранения устойчивости рассматриваемой системы.
Живая система всегда оказывается приспособленной к условиям существования, что считают наиболее характерной чертой жизни. При этом любой адаптационный процесс, по свидетельству Г. Селье (1960) и многих других, по своей сути есть процесс энергетический, не отделимый от других энергетических процессов организма и требующий постоянного притока энергии извне.
По мере повышения уровня организации организмов наряду с совершенствованием использования энергии повышается их зависимость от источников энергии. У гетеротрофов пища служит единственным источником пластического и энергетического обеспечения. Насекомые благодаря своим специфическим особенностям могут быть отнесены к организмам с очень высоким уровнем энергетических затрат.
Видовые адаптации, по свидетельству С.С. Шварца (1969), весьма разнообразны и необозримо совершенны, что определяется не только их функциональной активностью, но и энергетической стоимостью. Отсюда становится очевидным значение экономизации обмена и совершенствования способности к накоплению энергетических резервов как важнейших для адаптации. Известно, что в норме функциональные механизмы организма действуют с минимальной затратой энергии.
Экономия энергии, даже, казалось бы, самая ничтожная, может иметь решающее значение в жизнедеятельности и эволюции организмов, в том числе и насекомых. Особенно важным, по мнению С.С. Шварца (1969), следует считать соотношение энергетических потребностей животного с затратами энергии на добывание корма, что во многих случаях предопределяет пути микроэволюционных преобразований. В сопряженной эволюции с консументами основным направлением приспособительной изменчивости у растения-хозяина было развитие защитных механизмов от многочисленных врагов, в том числе и насекомых. С этим, очевидно, связана часто проявляющаяся общность факторов резистентности по отношению ко многим повреждающим агентам. В то же время адаптации у фитофагов оказались направленными на приспособление к наиболее рациональному использованию материально-энергетических ресурсов, поставляемых кормовым растением. Эта тенденция выразилась у фитофагов в многочисленных приспособлениях, затронувших различные уровни и организации. Это привело к становлению гостальной, топической и онтогенетической специализации. У насекомых, в свою очередь, эволюционировали и механизмы защиты от отрицательных воздействий хозяина.
Применительно к фитофагам различают несколько категорий пищевой специализации:
1) гостальную специализацию, приводящую к приобретению способности нормально существовать и развиваться лишь на растениях-хозяевах, относящихся к определенным систематическим группам;
2) топическую специализацию, при которой виды оказываются способными развиваться лишь при условии особой локализации на хозяевах;
3) онтогенетическую специализацию, приводящую к возникновению способности развиваться на органах хозяина, находящихся в определенном для каждого вида фитофага возрасте и морфофизиологическом состоянии.
При этом гостальная специфичность выступает как основная категория специализации, производными от которой являются две другие взаимосвязанные категории: топическая и онтогенетическая,
С точки зрения понимания природы устойчивости растений большой интерес представляет деление гостальной специфичности в соответствии с различиями таксономического ранга хозяев на:
1) специфичность крупных таксонов потребителей к крупным таксонам хозяев;
2) специфичность семейств или основных надсемейственньгх таксонов по отношению к роду, семейству или основным надсемейственным таксонам хозяев;
3) специфичность к виду и надвидовым таксонам хозяев;
4) специфичность вида-паразита или симбионта к внутривидовым группам, видам или родам хозяев (Слепян, 1973). Заслуживает внимания при рассмотрении гостальной специализации наличие существенных различий между так называемой реальной и потенциальной специфичностью, отмеченной у паразитов по отношению к их хозяевам (Киршенблат, 1941; Догель, 1962).
Различия между растениями, относящимися к группе потенциально возможных для питания и растениями оптимальными, зависят от следующих причин:
1) форм поведения, в которых выявляются избирательные отношения насекомого к определенным растениям, обусловленные реакцией на тот или иной признак последних;
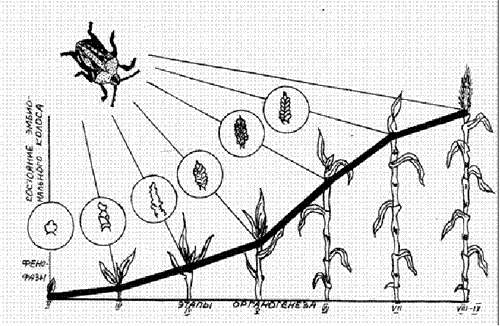
2) влияния растения на насекомое при питании. Например, клопы (вредная черепашка) меняют место питания в зависимости от этапа органогенеза растений (рис. 2). Сочетание этих условий и определяет широкий спектр внутривидового разнообразия градаций устойчивости растений к вредителям.
Рис. 2. Изменение мест питания вредной черепашки в связи с этапами органогенеза растений (по К.Е. Воронину, 1988).
Общепринятое деление фитофагов по широте пищевых связей на поли-, олиго– и монофагов, по существу, не отражает существования в каждой группе предпочтения определенного круга растений, в том числе и внутривидовых форм, которыми изобилуют культурные виды растений. Среди фитофагов наиболее широко представлены поли– и олигофага, включающие виды с более широкой или узкой специализацией. Доместикация и селекция растений привели к широкому их полиморфизму, что, в свою очередь, углубило процессы пищевой специализации фитофагов. Часто генетическая природа этих форм бывает гораздо сложнее, чем у диких видов.
Необходимость вычленения специфичности фитофагов к внутривидовым формам растений (сортам, гибридам, линиям) совершенно очевидна. Этот уровень пищевой специализации определяется факторами сортовой устойчивости растений. Н.И. Вавилов (1939) придавал большое значение наряду с родовым и видовым иммунитетом разнообразию представленности иммунитета полиморфных видов – от практически полной их резистентности до сильной восприимчивости.
Таким образом, напряжение иммунитета к вредителям на посевах культурных растений поддерживается иммуногенной защитой разных таксономических категорий растений: семейств, подсемейств, триб, родов, видов и внутривидовых форм. Выделение этих форм иммунитета растений базируется на соответствующих уровнях гостальной специализации фитофагов. Устойчивость растений к поли-, олиго– и монофагам может определяться каждой из названных форм иммунитета или их сочетанием. Характерно, что для полифагов большое значение имеют факторы, определяющие внутривидовую (сортовую) устойчивость. Вместе с тем на первых этапах изучения природы иммунитета растений к вредителям сложилось мнение о трудности нахождением устойчивых к многоядным вредителям форм растений и невозможности их создания (Щеголев, 1938). Однако, как показывает мировой опыт, наиболее значительные успехи были достигнуты в селекции устойчивых к полифагам и широким олигофагам сортов сельскохозяйственных культур.
В отношении гостальной специфичности, особенно видов с широким кругом кормовых растений, можно выделить три уровня специализации:
1) специализация, ограничивающая круг растений, возможных для питания в пределах относительно крупных их таксономических групп;
2) специализация, выражающаяся в предпочтении определенных форм растений из числа возможных;
3) специализация, выражающаяся в предпочтении внутривидовых форм полиморфных видов растений.
Поиски причин подобной специализации, предпринятые многими исследователями, породили различные гипотезы, трактующие избирательность насекомых по отношению к растениям с самых разных позиций.
Топическая и онтогенетическая специализация фитофагов выступают для каждого вида растений и животных как неизбежность биологического прогресса, в связи с чем становится очевидным, что избирательное отношение фитофагов к видам и внутривидовым формам кормовых растений следует рассматривать лишь как ступень пищевой специализации насекомых.
Рассмотрение гостальной специализации фитофагов создает некоторую иллюзию их широких возможностей в использовании разнообразных пищевых субстратов, представляемых различными органами растений. Однако из огромного числа фитофагов лишь ограниченное число видов способно питаться на всех без исключения органах растений. Среди фитофагов четко выделяются строго специализированные группы: филофаги, ксилофаги, ризофаги, антофаги, карпофаги др. Становление топической специализации, помимо органотропности – приуроченности к определенным органам и системам органов, проявляется на тканевом (гистротропность) и клеточном (цитотропность) уровнях, когда в пределах каждого органа растения фитофаги предпочитают питаться определенными тканями и их структурами.
Можно полагать, что обилие видов фитофагов, приуроченных к вегетативным органам растений, отражает особенности прогрессивной эволюции покрытосеменных. Ароморфоз цветковых растений, по мнению ряда исследователей, базировался в первую очередь на постоянных, т. е. вегетативных органах (Крыжановский, 1958; Яценко-Хмелевский, 1958; Попов, 1963; Проханов, 1965; Тихомиров, 1965, и др.). Очевидно, в связи с этим высокого совершенства достигла и иммуногенная система вегетативной сферы растений. Важные физиологические функции, выполняемые вегетативными органами, и обилие массы имели, как можно полагать, решающее значение для приспособительной эволюции фитофагов. Существенным в становлении специфичности фитофагов следует считать и тот факт, что «основной линией прогрессивной эволюции высших растений является не усовершенствование структур, а убыстрение темпов роста и развития». Это свойство, присущее покрытосеменным, усилило значение ростового барьера как одного из важных элементов иммунной системы растений. В то же время ускорение темпов роста и происходящую в связи с этим легкость регенерации точек роста у покрытосеменных М.Г. Попов (1963) связывает с увеличением и усложнением белковой молекулы. Следствием этих существенных преобразований белка явился синтез самых разнообразных форм побочных продуктов жизнедеятельности растений: алкалоидов, эфирных масел, органических кислот и т. д. Эти соединения, обладая широким спектром антибиотического действия, играют огромную роль в иммуногенной системе растений.
Существенными во взаимосвязях с насекомыми следует признать такие онтогенетические особенности растений, как способность к восстановлению органов благодаря специфическим тканям – первичной и вторичной меристемам – и способность к ослаблению связей с внешней средой или сведение этого взаимодействия к минимуму в отдельные периоды онтогенеза, выраженные физической и физиологической изоляцией.
Экологическое своеобразие системы «насекомое – фитофаг – кормовое растение» определяется необходимостью периодического возобновления взаимосвязей между составляющими ее элементами – растением и насекомым. Особенно яркую выраженность онтогенетической периодичности становления система получила во взаимоотношениях фитофагов с культурными растениями, где в результате ротации культур в севообороте происходят частые смены стаций. В связи с этим результативность взаимодействия растений и их потребителей – фитофагов – в значительной степени зависит от синхронизации развития партнеров.
Надежность и устойчивость взаимосвязей фитофагов с кормовыми растениями обеспечивается согласованностью в пространстве и времени определенных морфофункциональных изменений тех и других организмов (Шапиро, 1962,1985; Вилкова, 1980).
Следствием этого явилось становление у насекомых особых типов сопряженности циклов развития с циклами развития хозяев. Приуроченность к определенным возрастным периодам развития растений, в основе которой лежит топическая специфичность, свойственна всем группам фитофагов. Однако более четко выраженная зависимость от определенных этапов органогенеза кормового растения проявляется у фитофагов, приспособившихся к внутритканевому образу жизни.
Таким образом, в сопряженной эволюции с растением-хозяином формирование приспособительных черт фитофагов проходило на фоне сложных кинетических изменений, происходящих в онтогенезе растений, носящих характер циклических – ростовых и органообразовательных процессов, в основе которых лежат физико-химические превращения энергетических и пластических веществ в растениях. В связи с этим приспособительная эволюция фитофагов была подчинена особенностям морфоанатомической конституции, характеру и темпам формирования и дифференциации различных органов и тканей растений в онтогенезе последних, особенностям синтеза, транспорта и резервирования растениями веществ основного и «вторичного» обменов. В связи с этим можно полагать, что адаптации фитофагов были направлены в сторону морфофизиологических приспособлений, обеспечивающих потребителям относительно гомеостатические условия использования в пищу непрерывно меняющихся в онтогенезе растений форм и уровня содержания пластических и энергетических ресурсов (Шапиро, Вилкова, 1973).
Этот процесс нашел свое выражение в становлении у насекомых строгой приуроченности к использованию лишь определенных органов и тканей, находящихся на тех или иных этапах формирования. Так, для многих видов насекомых хорошо известна приуроченность к питанию осевыми органами, находящимися на определенных этапах онтогенеза. Еще более выражена приуроченность многих видов насекомых к питанию репродуктивными органами лишь на тех или иных этапах органогенеза. Во многих случаях относительно гомеостатические условия питания насекомых обеспечиваются путем приспособления к питанию строго определенными тканями растений. Это особенно ярко проявляется в приспособлении многих видов сосущих насекомых к высасыванию соков из эмбриональных тканей, паренхимы, флоэмы и других специализированных тканей растений. Принцип анализа сопряженности онтогенезов фитофагов и их кормовых растений позволил подойти к пониманию сущности топической и онтогенетической специализации многих видов.
Приобретению фитофагами в процессе эволюции приуроченности к оптимальным кормовым растениям, определенного морфофизиологического состояния, способствовала необходимость преодоления барьеров иммунной системы растений – тканевого (эпидерма, пробка, склеренхима, древесина), осмотического, ростового, органогенетического (большая скорость роста и дифференциации органов и тканей) и метаболического – наличие токсических соединений. Этот процесс нашел выражение в становлении у насекомых строгой приуроченности к использованию определенных органов тканей растений, находящихся на тех или иных этапах формирования.
Конец бесплатного ознакомительного фрагмента