Страница:
Рис. 24. Поперечный разрез зерна гречихи: а – эндосперм; б – семенная оболочка; в – плодовая оболочка; г – зародыш
Гречиха имеет высокую пленчатость, колеблющуюся от 20 до 46 %. По показателю пленчатости у нее установлены следующие градации:
высокопленчатая – от 22 % и выше;
среднепленчатая – от 20 до 22 %;
низкопленчатая – ниже 20 %.
На содержание плодовых оболочек заметное влияние оказывает величина зерна. Крупные фракции имеют несколько большую пленчатость.
Размеры зерна у гречихи колеблются в довольно широких пределах: длина от 5 до 7,6 мм, ширина от 2,8 до 4 мм, толщина от 2,9 до 5,2 мм. Ее размеры в практике работы с зерном принято выражать сходом с сита с круглыми отверстиями: крупной ш 3,9–4,2 мм, средней ш 3,6–3,7 мм и мелкой ш 2,2 мм.
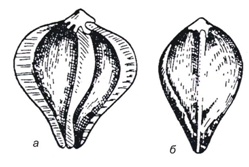 Рис. 25. Разновидность гречихи: а – крылатая; б – бескрылая
Рис. 25. Разновидность гречихи: а – крылатая; б – бескрылая
Повышенную пленчатость имеет так называемая «крылатая гречиха» с удлиненными гранями плодовых оболочек (рис. 25). Эти пленки предохраняют ядро от воздействия микроорганизмов, но они достаточно хрупки, способны к растрескиванию по месту соединения (граням). При нарушении целостности пленок создаются предпосылки как для механического повреждения ядра, имеющего мучнистый эндосперм и рыхлое строение клеток, так и для микробиологической порчи. Поэтому гречиха, особенно свежеубранная, нестойка при хранении.
Невызревшие и имеющие повышенную влажность зерна гречихи обладают низкими товарно-технологическими показателями. В свежеубранной партии зерна гречихи количество таких зерен обычно составляет 20–25 %. При подработке зернового вороха неполноценные зерна проходом через сито с отверстиями ш 3,0–3,4 мм уходят в отходы.
В процессе обмолота и послеуборочной подработки жесткая плодовая оболочка гречихи, неплотно прилегающая к ядру, легко отделяется. Ядра без плодовой оболочки плохо хранятся, их удаляют в процессе послеуборочной подработки гречихи.
Наиболее часто встречающиеся виды механических повреждений семян гречихи:
повреждение плодовой оболочки;
раскрытие «замка» – расхождение до 0,5 мм лепестков оболочки по граням в верхней части зерна;
шелушенное зерно – ядро без плодовой оболочки;
трещины в оболочке – трещины на лепестках оболочки;
срывы оболочек – частичная утрата лепестков оболочек;
повреждение ядра;
битое ядро;
частично расколотое ядро – кусочки ядра не менее 1/2 и не более 2/3 ядра;
трещины в ядре – форма ядра сохраняется, но видны неглубокие трещины в эндосперме.
По данным В. Б. Лебедева, от общего количества повреждений до 75 % приходится на плодовые оболочки в основном от раскрытия «замка», а также на образование трещин в плодовой оболочке и ее срыв. Обнаженное частично или полностью ядро в процессе хранения подвергается воздействию микрофлоры и вредителей.
Рис имеет пленчатую зерновку, не имеющую бороздки. Зерновка ребристая, часто покрыта волосками, сжатая с боков, реже округлая в разрезе, охватывается цветочными пленками, не удаляемыми при обмолоте. Цветочные пленки могут иметь разную окраску: желтую разных оттенков, буро-фиолетовую или буро-красную. Ребра и грани зерновки также могут иметь разную окраску. Полностью охватывая ядро, цветочная пленка риса срастается с ним только в некоторых участках. Как и у всех пленчатых культур, плодовая и семенная оболочки тонкие. Алейроновый слой состоит из одного ряда клеток (рис. 26).
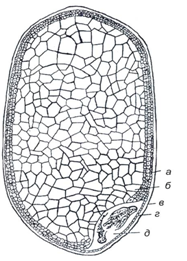 Рис. 26. Зерновка риса (продольный разрез): а – алейроновый слой; б – эндосперм; в – плодовая оболочка; г– семенная оболочка; д – зародыш
Рис. 26. Зерновка риса (продольный разрез): а – алейроновый слой; б – эндосперм; в – плодовая оболочка; г– семенная оболочка; д – зародыш
Длина зерновки колеблется от 4,4 до 10 мм, шир ина от 1,3 до 3,5 мм. Форма зерновки различна и является одним из признаков товарной классификации риса: 1-й тип – зерно продолговатое широкое; 2-й тип – зерно продолговатое тонкое, узкое; 3-й тип – зерно округлое.
С учетом длины и ширины зерновки различают рис индийской ветви – с узкими зерновками, длина превышает ширину в 3 раза и более и китайско-японскую ветвь, с отношением длины к ширине 1,4: 2,9.
У риса на долю цветковых пленок приходится 17–25 % массы зерновки, на долю оболочек 4–5 %, алейронового слоя 12–14 %, зародыша 2–3 %, эндосперма 65–67 %. По пленчатости сорта риса можно разделить на три группы: низкая пленчатость – 16–17 %, средняя – 18–19 %, высокая – 20 % и более. При обмолоте в зерновую массу попадают недозревшие (зеленые и меловые) зерна, что является следствием неравномерного развития и созревания зерна в различных частях метелки.
Культура риса – типичный гидрофит, дает урожаи зерна на периодически затопляемых полях, к моменту уборки его зерновая масса всегда имеет высокую влажность. В процессе послеуборочной подработки и хранения отдельные зерновки риса, вследствие меланоидинообразования желтеют, снижаются товарные достоинства вырабатываемой из такого зерна крупы.
Из-за высокого содержания крахмала (58–80 %) эндосперм у риса непрочный, хрупкий, а из-за неравномерного распределения влаги в зерновке в ядре возникают трещины. Они появляются вследствие того, что центральная часть эндосперма, по сравнению с внешними слоями, имеет повышенное содержание влаги. При сушке зерна эти более сухие слои зерновки сжимаются, происходит их усадка, что вызывает появление на поверхности зерновки трещин. Микроповреждение зерновок риса происходит в период вегетации растений, а затем влага, адсорбированная микротрещинами и проникшая в эндосперм под действием капиллярного давления, завершает процесс образования повреждений. Зерновка риса легко повреждается и при обмолоте. Чаще в зерновке риса образуются поперечные трещины, расположенные перпендикулярно к продольной оси зерновки.
Отмеченные особенности зерновок риса свидетельствуют о его нестойкости в хранении и необходимости проведения тщательных, разносторонних технологических мероприятий по подготовке к хранению, доведению показателей качества до базисных кондиций.
Подсолнечник имеет плод семянку, состоящую из околоплодника (плодовой оболочки, кожуры, лузги) и заключенного в нем семени (ядра) (рис. 27).
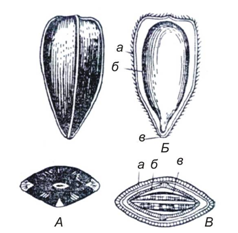 Рис. 27. Строение семянки подсолнечника: А – общий вид семянки сбоку и сверху; Б – продольный разрез: а – плодовая оболочка; б – ядро; в – место прикрепления семени;
Рис. 27. Строение семянки подсолнечника: А – общий вид семянки сбоку и сверху; Б – продольный разрез: а – плодовая оболочка; б – ядро; в – место прикрепления семени;
В – поперечный разрез: а – плодовая оболочка; б – семенная оболочка; в – семядоли
Околоплодник состоит из эпидермиса (кожицы), пробковой ткани и нескольких слоев одревесневших клеток склеренхимы. В верхних клетках склеренхимы может содержаться вещество черного цвета – фитомелан. Состоя на 76 % из углерода, фитомелан образует между пробковым слоем и склеренхимой панцирный слой, предохраняющий семянки от опасного для подсолнечника вредителя – подсолнечной моли (рис. 28). Семенная оболочка представлена тонкой пленкой, состоящей из внешней (бахромчатой) и внутренней (эпидермиса) тканей.
Семядоли подсолнечника толстые, мясистые, в них сконцентрированы все запасные липиды и белки.
Лузжистость семянок масленичного подсолнечника колеблется в пределах 35–45 %.
Из-за повышенного содержания в семянках жира (27 % и более) подсолнечник не стоек в хранении и требует к себе особого внимания.
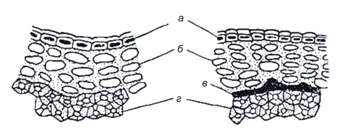 Рис. 28. Разрез плодовой оболочки семянки подсолнечника (слева – беспанцирного, справа – панцирного): а – эпидермис; б – пробковая ткань; в – панцирный слой; г – склеренхима
Рис. 28. Разрез плодовой оболочки семянки подсолнечника (слева – беспанцирного, справа – панцирного): а – эпидермис; б – пробковая ткань; в – панцирный слой; г – склеренхима
1.3. Физико-механические свойства единичных зерен и семян
1.4. Покой и долговечность зерна и семян
Гречиха имеет высокую пленчатость, колеблющуюся от 20 до 46 %. По показателю пленчатости у нее установлены следующие градации:
высокопленчатая – от 22 % и выше;
среднепленчатая – от 20 до 22 %;
низкопленчатая – ниже 20 %.
На содержание плодовых оболочек заметное влияние оказывает величина зерна. Крупные фракции имеют несколько большую пленчатость.
Размеры зерна у гречихи колеблются в довольно широких пределах: длина от 5 до 7,6 мм, ширина от 2,8 до 4 мм, толщина от 2,9 до 5,2 мм. Ее размеры в практике работы с зерном принято выражать сходом с сита с круглыми отверстиями: крупной ш 3,9–4,2 мм, средней ш 3,6–3,7 мм и мелкой ш 2,2 мм.
Повышенную пленчатость имеет так называемая «крылатая гречиха» с удлиненными гранями плодовых оболочек (рис. 25). Эти пленки предохраняют ядро от воздействия микроорганизмов, но они достаточно хрупки, способны к растрескиванию по месту соединения (граням). При нарушении целостности пленок создаются предпосылки как для механического повреждения ядра, имеющего мучнистый эндосперм и рыхлое строение клеток, так и для микробиологической порчи. Поэтому гречиха, особенно свежеубранная, нестойка при хранении.
Невызревшие и имеющие повышенную влажность зерна гречихи обладают низкими товарно-технологическими показателями. В свежеубранной партии зерна гречихи количество таких зерен обычно составляет 20–25 %. При подработке зернового вороха неполноценные зерна проходом через сито с отверстиями ш 3,0–3,4 мм уходят в отходы.
В процессе обмолота и послеуборочной подработки жесткая плодовая оболочка гречихи, неплотно прилегающая к ядру, легко отделяется. Ядра без плодовой оболочки плохо хранятся, их удаляют в процессе послеуборочной подработки гречихи.
Наиболее часто встречающиеся виды механических повреждений семян гречихи:
повреждение плодовой оболочки;
раскрытие «замка» – расхождение до 0,5 мм лепестков оболочки по граням в верхней части зерна;
шелушенное зерно – ядро без плодовой оболочки;
трещины в оболочке – трещины на лепестках оболочки;
срывы оболочек – частичная утрата лепестков оболочек;
повреждение ядра;
битое ядро;
частично расколотое ядро – кусочки ядра не менее 1/2 и не более 2/3 ядра;
трещины в ядре – форма ядра сохраняется, но видны неглубокие трещины в эндосперме.
По данным В. Б. Лебедева, от общего количества повреждений до 75 % приходится на плодовые оболочки в основном от раскрытия «замка», а также на образование трещин в плодовой оболочке и ее срыв. Обнаженное частично или полностью ядро в процессе хранения подвергается воздействию микрофлоры и вредителей.
Рис имеет пленчатую зерновку, не имеющую бороздки. Зерновка ребристая, часто покрыта волосками, сжатая с боков, реже округлая в разрезе, охватывается цветочными пленками, не удаляемыми при обмолоте. Цветочные пленки могут иметь разную окраску: желтую разных оттенков, буро-фиолетовую или буро-красную. Ребра и грани зерновки также могут иметь разную окраску. Полностью охватывая ядро, цветочная пленка риса срастается с ним только в некоторых участках. Как и у всех пленчатых культур, плодовая и семенная оболочки тонкие. Алейроновый слой состоит из одного ряда клеток (рис. 26).
Длина зерновки колеблется от 4,4 до 10 мм, шир ина от 1,3 до 3,5 мм. Форма зерновки различна и является одним из признаков товарной классификации риса: 1-й тип – зерно продолговатое широкое; 2-й тип – зерно продолговатое тонкое, узкое; 3-й тип – зерно округлое.
С учетом длины и ширины зерновки различают рис индийской ветви – с узкими зерновками, длина превышает ширину в 3 раза и более и китайско-японскую ветвь, с отношением длины к ширине 1,4: 2,9.
У риса на долю цветковых пленок приходится 17–25 % массы зерновки, на долю оболочек 4–5 %, алейронового слоя 12–14 %, зародыша 2–3 %, эндосперма 65–67 %. По пленчатости сорта риса можно разделить на три группы: низкая пленчатость – 16–17 %, средняя – 18–19 %, высокая – 20 % и более. При обмолоте в зерновую массу попадают недозревшие (зеленые и меловые) зерна, что является следствием неравномерного развития и созревания зерна в различных частях метелки.
Культура риса – типичный гидрофит, дает урожаи зерна на периодически затопляемых полях, к моменту уборки его зерновая масса всегда имеет высокую влажность. В процессе послеуборочной подработки и хранения отдельные зерновки риса, вследствие меланоидинообразования желтеют, снижаются товарные достоинства вырабатываемой из такого зерна крупы.
Из-за высокого содержания крахмала (58–80 %) эндосперм у риса непрочный, хрупкий, а из-за неравномерного распределения влаги в зерновке в ядре возникают трещины. Они появляются вследствие того, что центральная часть эндосперма, по сравнению с внешними слоями, имеет повышенное содержание влаги. При сушке зерна эти более сухие слои зерновки сжимаются, происходит их усадка, что вызывает появление на поверхности зерновки трещин. Микроповреждение зерновок риса происходит в период вегетации растений, а затем влага, адсорбированная микротрещинами и проникшая в эндосперм под действием капиллярного давления, завершает процесс образования повреждений. Зерновка риса легко повреждается и при обмолоте. Чаще в зерновке риса образуются поперечные трещины, расположенные перпендикулярно к продольной оси зерновки.
Отмеченные особенности зерновок риса свидетельствуют о его нестойкости в хранении и необходимости проведения тщательных, разносторонних технологических мероприятий по подготовке к хранению, доведению показателей качества до базисных кондиций.
Подсолнечник имеет плод семянку, состоящую из околоплодника (плодовой оболочки, кожуры, лузги) и заключенного в нем семени (ядра) (рис. 27).
В – поперечный разрез: а – плодовая оболочка; б – семенная оболочка; в – семядоли
Околоплодник состоит из эпидермиса (кожицы), пробковой ткани и нескольких слоев одревесневших клеток склеренхимы. В верхних клетках склеренхимы может содержаться вещество черного цвета – фитомелан. Состоя на 76 % из углерода, фитомелан образует между пробковым слоем и склеренхимой панцирный слой, предохраняющий семянки от опасного для подсолнечника вредителя – подсолнечной моли (рис. 28). Семенная оболочка представлена тонкой пленкой, состоящей из внешней (бахромчатой) и внутренней (эпидермиса) тканей.
Семядоли подсолнечника толстые, мясистые, в них сконцентрированы все запасные липиды и белки.
Лузжистость семянок масленичного подсолнечника колеблется в пределах 35–45 %.
Из-за повышенного содержания в семянках жира (27 % и более) подсолнечник не стоек в хранении и требует к себе особого внимания.
1.3. Физико-механические свойства единичных зерен и семян
Геометрическая форма и линейные размеры – длина (наибольший размер), ширина (наибольший поперечный размер), толщина (наименьший поперечный размер) – это физико-механические свойства единичных семян и зерен.
Совокупность линейных размеров получила название крупности зерна. Крупное зерно в технологическом отношении является наиболее ценным, т. к. у него более высокое содержание эндосперма, следовательно, и повышенный выход муки и крупы.
Зерно, однородное по крупности, получило название выравненного. Крупность и выравненность зерна определяются путем просеивания навесок на ситах с отверстиями различного сечения. При этом, если основная масса зерна остается на каком-либо одном сите или на двух смежных ситах, то такая партия зерна считается выравненной. Для выравненного зерна при его очистке на зерноочистительных машинах легче подобрать сита, отрегулировать воздушный поток, и в отходах меньше попадет зерна основной культуры.
Форма зерна определяется как соотношением размеров (длины, ширины, толщины), так и выполненностью, т. е. объемом и очертаниями. Форма зерен и семян весьма разнообразна, присуща данной культуре, виду, разновидности и сорту. Даже у одних и тех же культур форма семян весьма разнообразна. Например, семена фасоли могут быть овальными, почкообразными, цилиндрическими, яйцевидными. Разные виды пшеницы также различаются по форме. У вида мягкой пшеницы зерна округло-удлиненные, у твердой – более удлиненные. Выполненные зерна пшеницы более овальные, а щуплые более удлиненные.
Форма зерна может быть удлиненно-овальной (у пшеницы), удлиненной (у ржи, овса), яйцевидной (у пшеницы, фасоли), шарообразной (у гороха, горчицы, рапса), округлой (у проса, сорго), чечевицеобразной (у чечевицы), эллипсоида вращения (у фасоли, сои, бобов), треугольной (у гречихи).
Для характеристики формы зерна введены понятия «коэффициент формы» и «показатель сферичности».
Коэффициент формы – это отношение площади поверхности зерновки несферической формы к поверхности эквивалентного шара, объем которого равен объему зерновки.
Показатель сферичности – это отношение площади поверхности равновеликого по объему шара к площади внешней поверхности зерновки несферической формы. Для шарообразных зерен эти характеристики равны 1, для зерна иной формы они всегда больше 1, а показатель сферичности меньше 1.
Форма зерна тесно связана с содержанием пленок и оболочек. У зерен, по форме близких к шару, при прочих равных условиях обычно ниже пленчатость и содержание оболочек. Форма зерна влияет и на свойства зерновой массы. Зерновая масса, состоящая из зерен шаровидной формы, имеет большую сыпучесть, но меньшую скважистость по сравнению с зерновой массой, состоящей из зерен иной формы.
Форма и размер зерна и семян положены в основу очистки зерна от примесей и выделения крупных фракций, имеющих повышенные посевные и технологические показатели качества. Размеры семян и зерна варьируются довольно широко (табл. 5).
Таблица 5
Размеры зерен и семян различных культур, мм
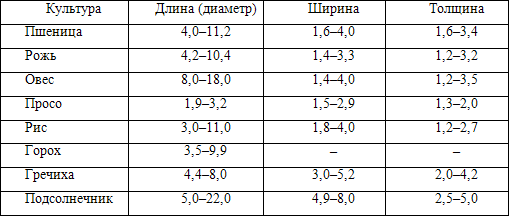 Влажные зерна всегда крупнее, чем сухие, по ширине и толщине, но практически не отличаются от сухих по длине.
Влажные зерна всегда крупнее, чем сухие, по ширине и толщине, но практически не отличаются от сухих по длине.
Совокупность линейных размеров получила название крупности зерна. Крупное зерно в технологическом отношении является наиболее ценным, т. к. у него более высокое содержание эндосперма, следовательно, и повышенный выход муки и крупы.
Зерно, однородное по крупности, получило название выравненного. Крупность и выравненность зерна определяются путем просеивания навесок на ситах с отверстиями различного сечения. При этом, если основная масса зерна остается на каком-либо одном сите или на двух смежных ситах, то такая партия зерна считается выравненной. Для выравненного зерна при его очистке на зерноочистительных машинах легче подобрать сита, отрегулировать воздушный поток, и в отходах меньше попадет зерна основной культуры.
Форма зерна определяется как соотношением размеров (длины, ширины, толщины), так и выполненностью, т. е. объемом и очертаниями. Форма зерен и семян весьма разнообразна, присуща данной культуре, виду, разновидности и сорту. Даже у одних и тех же культур форма семян весьма разнообразна. Например, семена фасоли могут быть овальными, почкообразными, цилиндрическими, яйцевидными. Разные виды пшеницы также различаются по форме. У вида мягкой пшеницы зерна округло-удлиненные, у твердой – более удлиненные. Выполненные зерна пшеницы более овальные, а щуплые более удлиненные.
Форма зерна может быть удлиненно-овальной (у пшеницы), удлиненной (у ржи, овса), яйцевидной (у пшеницы, фасоли), шарообразной (у гороха, горчицы, рапса), округлой (у проса, сорго), чечевицеобразной (у чечевицы), эллипсоида вращения (у фасоли, сои, бобов), треугольной (у гречихи).
Для характеристики формы зерна введены понятия «коэффициент формы» и «показатель сферичности».
Коэффициент формы – это отношение площади поверхности зерновки несферической формы к поверхности эквивалентного шара, объем которого равен объему зерновки.
Показатель сферичности – это отношение площади поверхности равновеликого по объему шара к площади внешней поверхности зерновки несферической формы. Для шарообразных зерен эти характеристики равны 1, для зерна иной формы они всегда больше 1, а показатель сферичности меньше 1.
Форма зерна тесно связана с содержанием пленок и оболочек. У зерен, по форме близких к шару, при прочих равных условиях обычно ниже пленчатость и содержание оболочек. Форма зерна влияет и на свойства зерновой массы. Зерновая масса, состоящая из зерен шаровидной формы, имеет большую сыпучесть, но меньшую скважистость по сравнению с зерновой массой, состоящей из зерен иной формы.
Форма и размер зерна и семян положены в основу очистки зерна от примесей и выделения крупных фракций, имеющих повышенные посевные и технологические показатели качества. Размеры семян и зерна варьируются довольно широко (табл. 5).
Таблица 5
Размеры зерен и семян различных культур, мм
1.4. Покой и долговечность зерна и семян
Для хранящихся масс зерновых и масличных культур состояние покоя – непременное условие стабилизации всех процессов жизнедеятельности, связанных как с проявлением дыхания, так и полным отсутствием прорастания семян.
Зерно, достигшее полной спелости и высушенное до влажности ниже критической, находится в состоянии покоя, который может быть обозначен и как анабиоз, т. е. состояние организма, при котором отсутствуют видимые проявления жизни, обнаруживающиеся однако при благоприятных условиях влажности и температуры. Явление анабиоза имеет большое значение для практики сельского хозяйства и хранения, т. к. именно в состоянии анабиоза зерно и семена хранятся в течение длительного срока. Некоторые ученые считают, что по отношению к зерну и семенам масличных культур следует применять термин «хозяйственный анабиоз», т. е. неполный анабиоз, в состоянии которого семена сохраняют свои биологические свойства как посевной материал и как сырье для переработки.
Семена, обладая необычным свойством, длительное время находятся в состоянии покоя, однако сохраняют свою жизнеспособность. При этом они могут находиться в состоянии вынужденного или органического покоя. Вынужденный покой обуславливают различные факторы внешней среды, такие как неблагоприятная температура для прорастания или недостаток влаги.
Посевной материал, находящийся в хранилищах в течение зимы, пребывает в вынужденном покое, для его прорастания нет необходимых условий. Но если в хранилищах создать благоприятные факторы (влажность и температуру), семена быстро прорастут.
Разнообразны причины, вызывающие у семян явление покоя. Впервые довольно полную классификацию типов покоя семян предложил в 1916 г. американский ученый В. Крокер. В ее основу были положены известные в то время причины, вызывающие покой:
1) недозрелость зародыша;
2) непроницаемость кожуры для воды;
3) механическое сопротивление покровов росту зародыша;
4) низкая газопроницаемость покровов;
5) покой, связанный с существованием метаболического блока;
6) комбинирование упомянутых причин;
7) вторичный покой.
Через 50 лет М. Г. Никалаева дала новую классификацию типов органического покоя семян, основанную на связи между причинами, обусловливающими покой, и условиями его преодоления. Ею были выделены три группы типов покоя семян: эндогенный, экзогенный и комбинированный. Причинами экзогенного покоя являются физические, механические, химические факторы; эндогенного – морфологические, физиологические и морфофизиологические факторы.
Органический покой характеризуется невозможностью прорастания, вызван свойствами зародыша или окружающих его тканей, а именно эндосперма и семенной кожуры, а также околоплодника или его частей.
Не прошедшие период послеуборочного дозревания семена находятся в так называемом первичном покое. В этом состоянии хорошо сформировавшиеся, жизнеспособные семена не способны прорастать. Такое «нежелание» семян прорастать при благоприятных для этого условиях – наличия влаги, тепла и кислорода атмосферного воздуха – вполне объяснимо с точки зрения этапов онтогенетического развития растений. Для семян яровых культур состояние покоя в зимний период – необходимый фактор для сохранения их в неблагоприятных условиях. Вызревшие сухие семена легко переносят низкие температуры. При замораживании в них не образуются кристаллы льда, разрушающие структуру клеток. Состояние покоя, как естественная стадия развития семян, является основополагающим фактором сохранения семян любой сельскохозяйственной культуры.
Свежеубранные сухие семена многих злаковых и других культур находятся в неглубоком физиологическом покое, который во времени обычно совпадает с периодом послеуборочного дозревания. При таком покое содержание влаги в семенах должно быть не выше критической величины (для хлебных злаков не более 15,5 %), которая характеризует в тканях только количество связанной влаги. При повышении влажности семян выше критической в структурах семян появляется свободная влага, гидролитические ферменты активизируются и начинается разрушение, происходит гидролиз запасных веществ семян.
Состояние покоя, его длительность у пшеницы М. И. Княгиничева связывает с пигментацией семян, накоплением флавоновых глюкозидов, которые тормозят прорастание. Впервые предположение о том, что белозерные сорта пшеницы имеют менее глубокий период покоя высказал еще в 1914 г. Нильсон-Элле. У этих сортов пшеницы короче и период послеуборочного дозревания.
Установлено, что продолжительность периода покоя, когда завершаются процессы послеуборочного дозревания, колеблется в зависимости от географических и метеорологических условий.
У озимой пшеницы в Ленинградской области он, например, длится до 60 дней, а в южных районах России от 5 до 20 дней. Из-за дождливой холодной погоды у пшеницы VI и XII этапы органогенеза обычно затягиваются, снижаются синтетические процессы, не образуются при этом некоторые, особенно физиологически активные, продукты синтеза. В такие годы семена характеризуются пониженной энергией прорастания и имеют длительный период дозревания и покоя.
Интенсивные осадки в период налива зерновок пшеницы приостанавливают синтетические процессы, происходит гидролиз крахмала зерна, оно становится щуплым, происходит вымывание дождевой водой из эндосперма растворимых углеводов.
Это явление, получившее название «стекание» зерна, приводит к снижению урожая пшеницы и резкому ухудшению семенных качеств. Такие зерна являются физиологически недозрелыми. В зернах пшеницы, не завершивших нормально XII этап органогенеза, длительное время идут процессы послеуборочного дозревания, затягивается вступление в состояние покоя.
Выявлено, что семена обычно переходят в состоянии покоя в период созревания на материнском растении, это так называемый первичный покой. Он обусловлен приспособлением растений к регулярно повторяющимся в определенные времена года неблагоприятным условиям. Растения реагируют на них задержкой развития и переходом в состояние покоя. Из внешних условий, влияющих на наступление покоя, можно выделить высокую температуру во время развития и созревания семян. Она ослабляет физиологическую деятельность созревающих семян.
Противоположным образом влияют низкие температуры, в результате чего в ранних фазах созревания семена, порой, не впадают в состояние покоя. Хранение таких семян сопряжено с определенными трудностями.
Покой семян пшеницы связывают с накоплением ряда веществ, тормозящих возможность прорастания зародыша зерновки в колосе вегетирующего растения. В процессе дозревания вещества, тормозящие рост, разрушаются, и у семян пшеницы появляется способность к прорастанию.
На длительность послеуборочного дозревания пшеницы оказывает влияние доступ воды и кислорода к зародышу, что в значительной мере связано со строением оболочек. Поступление воды к зародышу семян, как установила А. А. Алавдина еще в 1939 г., зависит от строения перикарпия зерновок. Кутикула, отделяющая перикарпий от семенной оболочки, непроницаема для воды. При снятии плодовой и семенной оболочек создается доступ кислорода к зародышу семени, и семена выходят из состояния покоя.
Ученые считают, что непроницаемость семенной кожуры оказывает влияние на изменение процесса дыхания, в результате чего образуются вещества – ингибиторы роста, тормозящие развитие семени. В качестве ингибиторов выступают образующиеся в семенной кожуре, эндосперме или околоплоднике различные химические соединения: фенолы, альдегиды, органические кислоты и т. д. Действие ингибиторов основано на блокировании систем ферментов и повышении энергии активации реакций, катализируемых этими ферментами, а также на инактивации ростовых веществ и на изменении структуры протоплазмы.
В процессе хранения большинство ингибиторов сами собой подвергаются распаду, и семена выходят из состояния покоя, всхожесть их повышается. Условия хранения семян зерновых культур оказывают непосредственное влияние на длительность их покоя, которая у зерновых резко сокращается в условиях хранения при температуре 38–40 °C и продлевается у семян, охлажденных до 2–3 °C.
Для большинства семян злаков главным фактором, регулирующим покой, является доступ кислорода воздуха к зародышу. Основная масса поступающего внутрь семени кислорода используется на дыхание, из-за чего повышение интенсивности неферментативных окислительных реакций происходит постепенно. Это и обуславливает продолжительность состояния покоя. У зерновок злаковых культур семенные оболочки и алейроновый слой задерживают доступ кислорода к зародышу, создают дефицит кислорода внутри семени, вызывают состояние покоя. Но и в состоянии покоя жизненные процессы в семенах полностью не прекращаются. Идет очень слабый газообмен, осуществляется медленный обмен веществ, расходуются запасные вещества, необратимо нарушаются структуры биомембран клеток, и при длительном хранении семена гибнут.
В технологии хранения в состоянии покоя считаются те семена, которые вследствие послеуборочной подработки и в результате дозревания в благоприятных для этого условиях установили минимальный уровень обмена веществ.
Покоящиеся сухие семена, с сильно ослабленными метаболическими процессами способны долго храниться и прорастать через длительный период времени. По способности сохранять жизнеспособность в оптимальных условиях хранения семена подразделяют на семена, способные сохранять жизнедеятельность в течение нескольких лет, и семена, сохраняющие жизнедеятельность в течение нескольких десятков лет.
В литературе, однако, встречаются противоречивые данные о невероятном долголетии отдельных видов семян. Так, К. Е. Овчаров описывает случаи, когда семена лотоса пролежали в земле около 3000 лет и дали всходы, или же семена арктического лютика пролежали в иле реки Миллер Крик (Канада) несколько веков и не потеряли жизнеспособности.
Долговечность семян зависит от многих причин: газонепроницаемости покровных тканей, химического состава, степени обводненности семян к уборке урожая и содержания влаги в хранящихся семенах, начального уровня обмена веществ, уровня зараженности и состава микрофлоры семян, условий хранения.
Жизнеспособность семян масличных культур, находящихся в состоянии влажности ниже критической, выше жизнеспособности семян, содержащих крахмал и белки, вследствие более высоких энергетических запасов и более совершенной теплоизоляции. В оптимальных условиях жизнеспособность семян, засыпанных на хранение, сохраняется длительное время. Однако в них постоянно идут необратимые процессы, вызывающие старение семян.
Старение свойственно всем живым организмам, и согласно общебиологическим законам, процесс этот является следствием наложения ошибок в системе хранения и реализации генетической информации, происходит постепенное нарушение структуры и функции клеток ткани и органов, которые увеличивают вероятность глубоких аномалий, приводящих к гибели организмов.
По У. Ф. Харрингтону, причинами старения семян являются различные разнохарактерные процессы: потеря дыхательной активности, разрушение хромосом, увеличение проницаемости мембран, образование токсичных продуктов, разрушение гормонов, необходимых для прорастания, денатурация белков и самоокисление жиров, сопровождаемое образованием свободных радикалов.
По мнению У. Ф. Харрингтона, основной причиной старения семян является распад хромосом. При старении семян происходит и распад кодирующегося механизма ДНК-РНК, принимающего участие в синтезе белков и ферментов.
В хранящихся семенах протекают катаболические процессы, приводящие к истощению запасов пластических веществ, изменению активности ферментов, повышению кислотности жиров, проницаемости мембран и т. д.
Детальную классификацию причин гибели семян дает Е. Н. Робертс. Ими, по его мнению, могут служить внешние и внутренние факторы (рис. 29). К внешним факторам относят действие ионизирующего излучения и влияние различных токсических веществ, которые выделяются микроорганизмами, находящимися в контакте с семенами.
Рассмотрим влияние отдельных этих факторов на старение семян.
Как показали исследования, ионизирующее излучение ускоряет потерю жизнеспособности семян при повышенных дозах радиации, а при обычных условиях фоновый уровень радиации не оказывает существенного влияния на долголетие семян.
Так же и микроорганизмы, и в первую очередь грибы, способны ослабить жизнеспособность семян, что нередко наблюдается при хранении семенного материала с повышенной влажностью. Сухие семена устойчивы к микрофлоре, находящейся на поверхности семени.
Следовательно, внешние факторы при соблюдении режима хранения семян не оказывают негативного воздействия на потери жизнеспособности.
Изучение влияния внутренних факторов старения семян показало, что в хранящихся семенах накапливаются различные классы соединений, которые могут обладать токсическим эффектом. Однако до настоящего времени четко не выявлено, являются ли они следствием старения семян или их причиной.
Денатурация макромолекул наблюдается в хранящихся семенах в белках, липопротеидных мембранах и нуклеиновых кислотах. В стареющих семенах изменяется количественный и качественный состав свободных аминокислот.
Как показали исследования А. Г. Малышевой, в период хранения семян подсолнечника происходит расщепление липидов до свободных жирных кислот, которые, в свою очередь, подвергаются окислению. Эти явления, возможно, и оказывают непосредственное влияние на снижение всхожести семян.
Образующиеся при окислении ненасыщенных жирных кислот свободные радикалы вызывают деструктивные изменения в клеточных липопротеиновых мембранах и снижение всхожести семян.
В процессе хранения семян наблюдается, особенно интенсивно при повышенной температуре и влажности, денатурация нуклеиновых кислот, вызывающая спонтанные мутации, при этом теряются системы, ответственные за «починку и замену» органелл.
Одной из причин снижения жизнеспособности семян является истощение дыхательного субстрата, в первую очередь уменьшение содержания белка и нередуцирующих сахаров и увеличение, как считает К. Е. Овчаров, содержания свободных жирных кислот и редуцирующих сахаров.
Зерно, достигшее полной спелости и высушенное до влажности ниже критической, находится в состоянии покоя, который может быть обозначен и как анабиоз, т. е. состояние организма, при котором отсутствуют видимые проявления жизни, обнаруживающиеся однако при благоприятных условиях влажности и температуры. Явление анабиоза имеет большое значение для практики сельского хозяйства и хранения, т. к. именно в состоянии анабиоза зерно и семена хранятся в течение длительного срока. Некоторые ученые считают, что по отношению к зерну и семенам масличных культур следует применять термин «хозяйственный анабиоз», т. е. неполный анабиоз, в состоянии которого семена сохраняют свои биологические свойства как посевной материал и как сырье для переработки.
Семена, обладая необычным свойством, длительное время находятся в состоянии покоя, однако сохраняют свою жизнеспособность. При этом они могут находиться в состоянии вынужденного или органического покоя. Вынужденный покой обуславливают различные факторы внешней среды, такие как неблагоприятная температура для прорастания или недостаток влаги.
Посевной материал, находящийся в хранилищах в течение зимы, пребывает в вынужденном покое, для его прорастания нет необходимых условий. Но если в хранилищах создать благоприятные факторы (влажность и температуру), семена быстро прорастут.
Разнообразны причины, вызывающие у семян явление покоя. Впервые довольно полную классификацию типов покоя семян предложил в 1916 г. американский ученый В. Крокер. В ее основу были положены известные в то время причины, вызывающие покой:
1) недозрелость зародыша;
2) непроницаемость кожуры для воды;
3) механическое сопротивление покровов росту зародыша;
4) низкая газопроницаемость покровов;
5) покой, связанный с существованием метаболического блока;
6) комбинирование упомянутых причин;
7) вторичный покой.
Через 50 лет М. Г. Никалаева дала новую классификацию типов органического покоя семян, основанную на связи между причинами, обусловливающими покой, и условиями его преодоления. Ею были выделены три группы типов покоя семян: эндогенный, экзогенный и комбинированный. Причинами экзогенного покоя являются физические, механические, химические факторы; эндогенного – морфологические, физиологические и морфофизиологические факторы.
Органический покой характеризуется невозможностью прорастания, вызван свойствами зародыша или окружающих его тканей, а именно эндосперма и семенной кожуры, а также околоплодника или его частей.
Не прошедшие период послеуборочного дозревания семена находятся в так называемом первичном покое. В этом состоянии хорошо сформировавшиеся, жизнеспособные семена не способны прорастать. Такое «нежелание» семян прорастать при благоприятных для этого условиях – наличия влаги, тепла и кислорода атмосферного воздуха – вполне объяснимо с точки зрения этапов онтогенетического развития растений. Для семян яровых культур состояние покоя в зимний период – необходимый фактор для сохранения их в неблагоприятных условиях. Вызревшие сухие семена легко переносят низкие температуры. При замораживании в них не образуются кристаллы льда, разрушающие структуру клеток. Состояние покоя, как естественная стадия развития семян, является основополагающим фактором сохранения семян любой сельскохозяйственной культуры.
Свежеубранные сухие семена многих злаковых и других культур находятся в неглубоком физиологическом покое, который во времени обычно совпадает с периодом послеуборочного дозревания. При таком покое содержание влаги в семенах должно быть не выше критической величины (для хлебных злаков не более 15,5 %), которая характеризует в тканях только количество связанной влаги. При повышении влажности семян выше критической в структурах семян появляется свободная влага, гидролитические ферменты активизируются и начинается разрушение, происходит гидролиз запасных веществ семян.
Состояние покоя, его длительность у пшеницы М. И. Княгиничева связывает с пигментацией семян, накоплением флавоновых глюкозидов, которые тормозят прорастание. Впервые предположение о том, что белозерные сорта пшеницы имеют менее глубокий период покоя высказал еще в 1914 г. Нильсон-Элле. У этих сортов пшеницы короче и период послеуборочного дозревания.
Установлено, что продолжительность периода покоя, когда завершаются процессы послеуборочного дозревания, колеблется в зависимости от географических и метеорологических условий.
У озимой пшеницы в Ленинградской области он, например, длится до 60 дней, а в южных районах России от 5 до 20 дней. Из-за дождливой холодной погоды у пшеницы VI и XII этапы органогенеза обычно затягиваются, снижаются синтетические процессы, не образуются при этом некоторые, особенно физиологически активные, продукты синтеза. В такие годы семена характеризуются пониженной энергией прорастания и имеют длительный период дозревания и покоя.
Интенсивные осадки в период налива зерновок пшеницы приостанавливают синтетические процессы, происходит гидролиз крахмала зерна, оно становится щуплым, происходит вымывание дождевой водой из эндосперма растворимых углеводов.
Это явление, получившее название «стекание» зерна, приводит к снижению урожая пшеницы и резкому ухудшению семенных качеств. Такие зерна являются физиологически недозрелыми. В зернах пшеницы, не завершивших нормально XII этап органогенеза, длительное время идут процессы послеуборочного дозревания, затягивается вступление в состояние покоя.
Выявлено, что семена обычно переходят в состоянии покоя в период созревания на материнском растении, это так называемый первичный покой. Он обусловлен приспособлением растений к регулярно повторяющимся в определенные времена года неблагоприятным условиям. Растения реагируют на них задержкой развития и переходом в состояние покоя. Из внешних условий, влияющих на наступление покоя, можно выделить высокую температуру во время развития и созревания семян. Она ослабляет физиологическую деятельность созревающих семян.
Противоположным образом влияют низкие температуры, в результате чего в ранних фазах созревания семена, порой, не впадают в состояние покоя. Хранение таких семян сопряжено с определенными трудностями.
Покой семян пшеницы связывают с накоплением ряда веществ, тормозящих возможность прорастания зародыша зерновки в колосе вегетирующего растения. В процессе дозревания вещества, тормозящие рост, разрушаются, и у семян пшеницы появляется способность к прорастанию.
На длительность послеуборочного дозревания пшеницы оказывает влияние доступ воды и кислорода к зародышу, что в значительной мере связано со строением оболочек. Поступление воды к зародышу семян, как установила А. А. Алавдина еще в 1939 г., зависит от строения перикарпия зерновок. Кутикула, отделяющая перикарпий от семенной оболочки, непроницаема для воды. При снятии плодовой и семенной оболочек создается доступ кислорода к зародышу семени, и семена выходят из состояния покоя.
Ученые считают, что непроницаемость семенной кожуры оказывает влияние на изменение процесса дыхания, в результате чего образуются вещества – ингибиторы роста, тормозящие развитие семени. В качестве ингибиторов выступают образующиеся в семенной кожуре, эндосперме или околоплоднике различные химические соединения: фенолы, альдегиды, органические кислоты и т. д. Действие ингибиторов основано на блокировании систем ферментов и повышении энергии активации реакций, катализируемых этими ферментами, а также на инактивации ростовых веществ и на изменении структуры протоплазмы.
В процессе хранения большинство ингибиторов сами собой подвергаются распаду, и семена выходят из состояния покоя, всхожесть их повышается. Условия хранения семян зерновых культур оказывают непосредственное влияние на длительность их покоя, которая у зерновых резко сокращается в условиях хранения при температуре 38–40 °C и продлевается у семян, охлажденных до 2–3 °C.
Для большинства семян злаков главным фактором, регулирующим покой, является доступ кислорода воздуха к зародышу. Основная масса поступающего внутрь семени кислорода используется на дыхание, из-за чего повышение интенсивности неферментативных окислительных реакций происходит постепенно. Это и обуславливает продолжительность состояния покоя. У зерновок злаковых культур семенные оболочки и алейроновый слой задерживают доступ кислорода к зародышу, создают дефицит кислорода внутри семени, вызывают состояние покоя. Но и в состоянии покоя жизненные процессы в семенах полностью не прекращаются. Идет очень слабый газообмен, осуществляется медленный обмен веществ, расходуются запасные вещества, необратимо нарушаются структуры биомембран клеток, и при длительном хранении семена гибнут.
В технологии хранения в состоянии покоя считаются те семена, которые вследствие послеуборочной подработки и в результате дозревания в благоприятных для этого условиях установили минимальный уровень обмена веществ.
Покоящиеся сухие семена, с сильно ослабленными метаболическими процессами способны долго храниться и прорастать через длительный период времени. По способности сохранять жизнеспособность в оптимальных условиях хранения семена подразделяют на семена, способные сохранять жизнедеятельность в течение нескольких лет, и семена, сохраняющие жизнедеятельность в течение нескольких десятков лет.
В литературе, однако, встречаются противоречивые данные о невероятном долголетии отдельных видов семян. Так, К. Е. Овчаров описывает случаи, когда семена лотоса пролежали в земле около 3000 лет и дали всходы, или же семена арктического лютика пролежали в иле реки Миллер Крик (Канада) несколько веков и не потеряли жизнеспособности.
Долговечность семян зависит от многих причин: газонепроницаемости покровных тканей, химического состава, степени обводненности семян к уборке урожая и содержания влаги в хранящихся семенах, начального уровня обмена веществ, уровня зараженности и состава микрофлоры семян, условий хранения.
Жизнеспособность семян масличных культур, находящихся в состоянии влажности ниже критической, выше жизнеспособности семян, содержащих крахмал и белки, вследствие более высоких энергетических запасов и более совершенной теплоизоляции. В оптимальных условиях жизнеспособность семян, засыпанных на хранение, сохраняется длительное время. Однако в них постоянно идут необратимые процессы, вызывающие старение семян.
Старение свойственно всем живым организмам, и согласно общебиологическим законам, процесс этот является следствием наложения ошибок в системе хранения и реализации генетической информации, происходит постепенное нарушение структуры и функции клеток ткани и органов, которые увеличивают вероятность глубоких аномалий, приводящих к гибели организмов.
По У. Ф. Харрингтону, причинами старения семян являются различные разнохарактерные процессы: потеря дыхательной активности, разрушение хромосом, увеличение проницаемости мембран, образование токсичных продуктов, разрушение гормонов, необходимых для прорастания, денатурация белков и самоокисление жиров, сопровождаемое образованием свободных радикалов.
По мнению У. Ф. Харрингтона, основной причиной старения семян является распад хромосом. При старении семян происходит и распад кодирующегося механизма ДНК-РНК, принимающего участие в синтезе белков и ферментов.
В хранящихся семенах протекают катаболические процессы, приводящие к истощению запасов пластических веществ, изменению активности ферментов, повышению кислотности жиров, проницаемости мембран и т. д.
Детальную классификацию причин гибели семян дает Е. Н. Робертс. Ими, по его мнению, могут служить внешние и внутренние факторы (рис. 29). К внешним факторам относят действие ионизирующего излучения и влияние различных токсических веществ, которые выделяются микроорганизмами, находящимися в контакте с семенами.
Рассмотрим влияние отдельных этих факторов на старение семян.
Как показали исследования, ионизирующее излучение ускоряет потерю жизнеспособности семян при повышенных дозах радиации, а при обычных условиях фоновый уровень радиации не оказывает существенного влияния на долголетие семян.
Так же и микроорганизмы, и в первую очередь грибы, способны ослабить жизнеспособность семян, что нередко наблюдается при хранении семенного материала с повышенной влажностью. Сухие семена устойчивы к микрофлоре, находящейся на поверхности семени.
Следовательно, внешние факторы при соблюдении режима хранения семян не оказывают негативного воздействия на потери жизнеспособности.
Изучение влияния внутренних факторов старения семян показало, что в хранящихся семенах накапливаются различные классы соединений, которые могут обладать токсическим эффектом. Однако до настоящего времени четко не выявлено, являются ли они следствием старения семян или их причиной.
Денатурация макромолекул наблюдается в хранящихся семенах в белках, липопротеидных мембранах и нуклеиновых кислотах. В стареющих семенах изменяется количественный и качественный состав свободных аминокислот.
Как показали исследования А. Г. Малышевой, в период хранения семян подсолнечника происходит расщепление липидов до свободных жирных кислот, которые, в свою очередь, подвергаются окислению. Эти явления, возможно, и оказывают непосредственное влияние на снижение всхожести семян.
Образующиеся при окислении ненасыщенных жирных кислот свободные радикалы вызывают деструктивные изменения в клеточных липопротеиновых мембранах и снижение всхожести семян.
В процессе хранения семян наблюдается, особенно интенсивно при повышенной температуре и влажности, денатурация нуклеиновых кислот, вызывающая спонтанные мутации, при этом теряются системы, ответственные за «починку и замену» органелл.
Одной из причин снижения жизнеспособности семян является истощение дыхательного субстрата, в первую очередь уменьшение содержания белка и нередуцирующих сахаров и увеличение, как считает К. Е. Овчаров, содержания свободных жирных кислот и редуцирующих сахаров.