Страница:
2. Разнообразие клетки
У многоклеточных организмов разные клетки (например, нервные, мышечные, клетки крови) выполняют разные функции («разделение труда») и поэтому различаются по своей структуре. Несмотря на это, многообразие форм и организация клеток подчинены единым структурным принципам.Форма клеток необычайно разнообразна – от простейшей шаровидной (одноклеточные организмы; среди бактерий – кокки) до самой причудливой. Микрококки имеют диаметр 0,2 мкм, нервные клетки достигают в длину 1 м, а млечные сосуды растений – даже нескольких метров.
3. Структура клетки
Живое содержимое клетки, протоплазма, отделено от окружающей среды плазматической мембраной (плазмолеммой) и может быть, кроме того, окружено прочной клеточной стенкой. Протоплазма представляет собой студнеобразную неоднородную массу с множеством различных органелл и параплазматических включений. Последние только условно причисляются к живой протоплазме и содержат вещества, подлежащие накоплению или выделению.4. Структурные элементы клетки
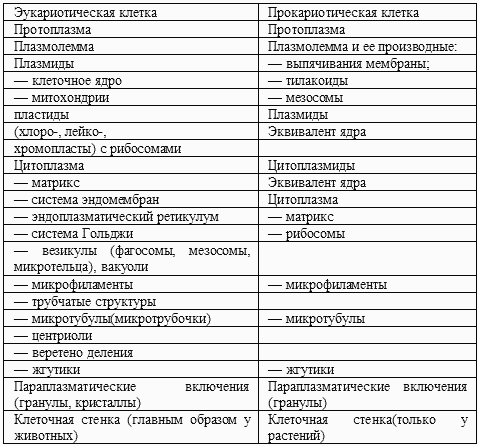
Существуют две ступени организации клетки: прокариотическая клетка (у прокариот – бактерий и синезеленых водорослей, в большинстве своем одноклеточных) и эукариотическая клетка (у эукариот, т. е. всех остальных одно– и многоклеточных организмов – растений, грибов и животных).Таблица 2. Структурные элементы клетки
Вопрос 15. Эукариотические и прокариотические клетки
1. Характеристика эукариотических клеток
Средняя величина эукариотической клетки – около 13 мкм (но существуют большие колебания в размерах). Клетка разделена внутренними мембранами на различные компартменты (реакционные пространства). Три вида органелл (пласты) четко отграничены от остальной протоплазмы (цитоплазмы) оболочкой из двух мембран: клеточное ядро, митохондрии и пластиды (последние только у растений). Пластиды служат главным образом для фотосинтеза, а митохондрии – для выработки энергии. Все пласты содержат ДНК в качестве носителя генетической информации.Цитоплазма содержит различные органеллы, большей частью видимые только с помощью электронного микроскопа, в том числе рибосомы, которые имеются также в пластидах и митохондриях. Все органеллы лежат в матриксе (это та часть цитоплазмы, которая даже в электронном микроскопе представляется гомогенной).
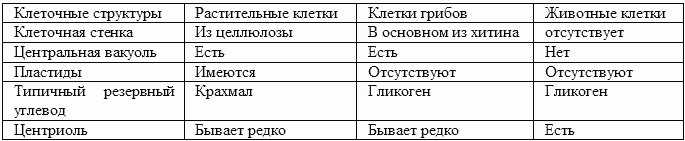
2. Основные формы эукариотических клеток
Существуют три основные формы эукариотических клеток: растительные клетки, клетки грибов и животные клетки.Таблица 3. Основные формы эукариотических клеток
3. Характеристика прокариотических клеток
Средняя величина прокариотических клеток составляет 5 мкм. У них нет никаких внутренних мембран, кроме выпячиваний внутренних мембран и плазматической мембраны. Пласты отсутствуют. Вместо клеточного ядра имеется его эквивалент (нуклеоид), лишенный оболочки и состоящий из одной-единственной молекулы ДНК. Кроме того, бактерии могут содержать ДНК в форме крошечных плазмид, сходных с внеядерными ДНК эукариот.В прокариотических клетках, способных к фотосинтезу (синезеленые водоросли, зеленые и пурпурные бактерии), имеются различно структурированные крупные выпячивания мембраны – тилакоиды, по своей функции соответствующие пластидам эукариот. Эти же тилакоиды или – в бесцветных клетках – более мелкие выпячивания мембраны (а иногда даже сама плазматическая мембрана) в функциональном отношении заменяют митохондрии. Другие сложно дифференцированные выпячивания мембраны называют мезосомами; их функция неясна. Только некоторые органеллы прокариотической клетки гомологичны соответствующим органеллам эукариот. Для прокариот характерно наличие муреннового мешка – механически прочного элемента клеточной стенки.
Вопрос 16. Вирусы
1. Характеристика вирусов
Вирусы представляют собой неклеточные образования – очень мелкие частицы (вирионы), состоящие из нуклеиновой кислоты (ДНК или РНК, одно– или двуцепочечной, служащей генетическим материалом) и белковой оболочки, иногда содержащей липиды.Оболочка (капсид) построена из субъединиц (капсомеров), которые состоят из одной или нескольких идентичных или разных полипептидных цепей.
Вирусы видоспецифичны и размножаются только в живых клетках-хозяевах. Существуют бактериальные вирусы (фаги), вирусы растений и вирусы животных. Вне клетки-хозяина вирионы не осуществляют обмена веществ и не проявляют никаких других признаков жизни.
2. Проникновение вируса в организм хозяина
В клетку-хозяин проникает вирион или только его нуклеиновая кислота. Там эта нуклеиновая кислота, используя систему репликации и белоксинтезирующий аппарат клетки хозяина, размножается (реплицируется) и обеспечивает синтез вирусного белка.У вирулентных вирусов образующиеся вирионы освобождаются постепенно или же все сразу в результате разрушения клетки. У умеренных фагов ДНК может быть встроена в ДНК клетки-хозяина в качестве провируса и реплицируется вместе с ней. Образование вирионов происходит лишь при его индукции различными факторами (облучение, химические агенты, повышенная температура).
Вирусы служат возбудителями болезней, поскольку при своем освобождении они разрушают клетку-хозяина или вызывают нарушение ее метаболизма.
Вопрос 17. Цитоплазма. Рибосомы и плазмиды
1. Состав цитоплазмы
Цитоплазмой мы называем живое содержимое клетки без пластов или эквивалента ядра. Цитоплазма представляет собой вязко-упругий тиксотропный гель.Вязко-упругие свойства и тиксотропность возможны только тогда, когда молекулы образуют сплошную сеть, которая может разрушаться и возникать вновь. Разрушение молекулярной сети приводит к проявлению жидкостных свойств, а ее восстановление – к свойствам, характерным для твердых тел. В цитоплазме элементами, способными сплетаться в сеть, служат длинные нитевидные микрофиламенты из белка актина, которые, вероятно, удерживаются вместе с помощью какого-то другого белка. При отщеплении молекул этого белка сеть распадается (состояние золя). Теперь микрофиламенты могут двигаться, и таким образом возникает течение протоплазмы, которое можно обнаружить в большинстве клеток.
2. Строение матрикса цитоплазмы
Матрикс цитоплазмы предоставляет собой гомогенную (при исследовании в электронном микроскопе) субстанцию между микрофиламентами. Она состоит из воды и множества растворенных неорганических и органических веществ, в частности ферментов и других белков. Матрикс цитоплазмы служит средой для диффузии многих промежуточных продуктов обмена, а также местом, где протекают важнейшие метаболические процессы, например гликолиз и пентозофосфатный цикл.Понятие «цитозоль» означает неосаждаемую при ультрацентрифугировании фракцию гомогената, которая содержит матрикс цитоплазмы и очень легкие структуры, такие как микрофиламенты. Оно применимо также к соответствующей фракции интактных клеток, хотя в клетке матрикс – не золь, а так же, как и остальная цитоплазма, представляет собой вязко-эластичный тиксотропный гель.
3. Характеристика рибосом
Рибосомы осуществляют биосинтез белка, реализуя таким образом генетическую информацию. Каждая клетка обладает десятками тысяч или миллионами этих крошечных, размером 20–30 нм, округлых рибонуклепротеидных частиц. Рибосома состоит из двух неодинаковых субчастиц. Они образуются отдельно и объединяются на и-РНК, что происходит по эксцентрически расположенному каналу между субчастицами и доставляет информацию для биосинтеза белка. При этом несколько рибосом могут быть связаны нитевидной молекулой и-РНК в полисому (полирибосому) наподобие нитки жемчуга.Более крупные рибосомы мы находим в цитоплазме эукариотических клеток. Они могут быть вместе с и-РНК связаны с эндоплазматическим ретикулом. Их субчастицы синтезируются в клеточном ядре. Прокариотические клетки обладают более мелкими рибосомами. Рибосомы чрезвычайно богаты магнием.
4. Плазмиды
Плазмиды – это находящиеся вне генома очень короткие двойные спирали ДНК, замкнутые в кольцо (длиной от нескольких до ста тысяч пар оснований), с одним или несколькими генами, а иногда и совсем без генов. Они реплицируются в большинстве случаев независимо от остального генетического материала и часто переходят из одной клетки в другую. В настоящее время они обнаружены у бактерий и дрожжей, а также в митохондриях эукариотических клеток. Некоторые бактериальные плазмиды могут включаться в геном и снова отделяться от него.
Вопрос 18. Мембраны, их молекулярная структура
1. Строение
Протоплазма ограничена наружной мембраной – плазмолеммой и содержит систему внутренних мембран (эндомембран). Митохондрии и пластиды, тоже имеющие внутренние мембраны и клеточное ядро, окружены двумя мембранами.Толщина мембраны чаще всего составляет 6—12 нм. Мембраны ограничивают замкнутые объемы различной величины и формы, например пузырьки, уплощенные полости или целые клетки. Таким образом, создавая препятствие для диффузии, они образуют отдельные реакционные объемы (компартменты). С другой стороны, мембраны способны избирательно пропускать некоторые вещества и активно накачивать другие, что связано с затратой энергии. Как полагают, каждая мембрана отделяет протоплазматическое пространство от неплазматического: плазмолемма – от окружающей клетку среды, мембраны пузырьков – от неплазматического содержимого этих пузырьков, обе мембраны ядерной оболочки – от неплазматического пространства, находящегося между ними.
Мембраны (за исключением мембран митохондрий и пластид) используются в процессах онтогенеза и могут превращаться друг в друга (течение мембран). Например, из эндоплазматического ретикулума образуются мембраны аппарата Гольджи, а последние служат материалом для регенерации плазмолеммы.
2. Белки и липиды в составе мембраны
Мембраны представляют собой двумерные жидкокристаллические растворы глобулярных белков в липидах. Структурную основу мембран составляют липиды, среди которых преобладают фосфолипиды (например, лецитин), а в мембранах пластид – гликолипиды. Белки в мембранах выполняют определенные функции: они являются, например, ферментами или транспортными белками. Кроме того, в состав мембран входят стерины (у животных а основном холестерин), гликопротеиды и некоторые неорганические соли.3. Основная структура мембран
Основная структура всех мембран представляет собой два параллельных слоя липидов (бимолекулярный слой). Мембранные липиды – амфипатические молекулы, имеющие гидрофобную часть (углеводородные остатки жирных кислот и сфингозина) и гидрофильную часть (фосфат, холин, комамин, сахар и т. п.). Такие молекулы образуют на водной поверхности мономолекулярный слой. В водном окружении и в клетке образуются бимолекулярные слои: гидрофобные части различных молекул повернуты дальше от водного окружения, т. е. друг к другу, и удерживаются вместе сильными гидрофобными взаимодействиями и слабыми силами Ван-дер-Ваальса.Таким образом, мембраны на обеих наружных поверхностях гидрофильны, а внутри – гидрофобны. Поскольку гидрофильные части молекул поглощают электроны, они видны в электронном микроскопе как два темных слоя.
4. Влияние температуры на мембрану
При низких температурах углеводородные остатки образуют подобие кристаллической решетки, и мембраны переходят в состояние геля. При физиологических температурах мембраны находятся в жидкокристаллическом состоянии: углеводородные остатки вращаются вокруг своей продольной оси и диффундируют в плоскости слоя; реже перескакивают из одного слоя в другой, не нарушая прочных гидрофобных связей.Периферические белки мембран гидрофильны, так как на поверхности их глобулярной молекулы преобладают гидрофильные аминокислоты (с полярными группами). Они относительно непрочно связаны с гидрофильными поверхностями мембран в основном электростатическими силами, т. е. ионными связями.
Интегральные мембранные белки гидрофобны (по крайней мере частично), так как на поверхности их молекул находятся главным образом гидрофобные аминокислотные остатки.
Эти белки прочно укреплены в гидрофобной толще мембраны гидрофобными взаимодействиями, а гидрофильные части молекул выступают из мембраны наружу. Некоторые интегральные белки мембран способны, как и липидные молекулы, диффундировать в плоскости мембраны, другие встроены неподвижно.
Описанная жидкостно-мозаичная модель структуры мембраны (модель Сингера) заменила принятую ранее модель Даниели (без интегральных белков).
Благодаря гидрофобным взаимодействиям мембраны способны растягиваться (расти) при включении новых молекул, а в случае разрыва образовавшиеся края могут снова смыкаться.
Мембраны полупроницаемы; они должны обладать мельчайшими порами, через которые диффундируют вода и другие небольшие гидрофильные молекулы. Вероятно, для этого используются внутренние гидрофильные области интегральных мембранных белков или отверстия между соприкасающимися интегральными белками (туннельные белки).
Вопрос 19. Плазматическая мембрана
1. Характеристика плазмолемм
Плазмолемма, толщина которой около 8 нм, выполняет роль барьера для диффузии веществ из клетки; это существенно и для растительных клеток, так как клеточная стенка, как правило, проницаема. Встроенные в мембрану транспортные молекулы осуществляют перенос определенных веществ. Мембранные ферменты принимают лишь ограниченное участие в метаболизме. У растений плазмолемма участвует в обмене компонентов клеточной стенки, в нервных клетках – в проведении импульсов.При клеточном делении дочерние клетки получают плазмолемму от материнской клетки. При росте плазмолеммы (связанном с делением и ростом клеток) и при ее регенерации она образуется из пузырьков Гольджи (течение мембран).
Плазматическая мембрана животных клеток покрыта снаружи полисахаридным слоем толщиной от 10 до 20 нм – гликокаликсом. Разветвленные остатки полисахаридов ковалентно связаны с белками и сфингозинсодержащими липидами. Полисахариды состоят главным образом из галактозы, маннозы, фукозы, N-ацетилгалактозамина, N-ацетилглюкозамина и (в концевых положениях) остатков сиаловой кислоты. Сиаловыми кислотами называют N-гликозил– и N-ацетилнейраминовые кислоты; нейраминовая кислота – это циклический конденсат маннозы и пирувата.
Из компонентов гликокаликса хорошо изучен гликопротеид гликофорин в мембранах эритроцитов. Он состоит на 60 % из углеводов и несет (подобно другим гликопротеидам и гликолипидам плазматических мембран животных клеток) специфические антигены групп крови, а также участки, связывающие различные вирусы и лектины.
Карбоксильный конец полипептидной цепи выступает из мембраны с ее внутренней стороны, а с наружной стороны находится аминный конец с многочисленными сильно разветвленными боковыми цепями полисахаридов.
2. Отличие плазматической мембраны в прокариотических клетках
Плазматическая мембрана прокариотических клеток отличается тем, что содержит в качестве интегральных белков переносчики электронов и ферменты дыхательной цепи и образует разного рода выпячивания. Некоторые выпячивания осуществляют дыхание, другие – фотосинтез и дыхание. Мезосомы бактерий представляют собой пластинчатые, трубчатые или везикулярные тельца, лежащие в карманах мембраны. Внутреннее пространство мезосом частично сообщается с внеклеточной средой. Мезосомы образуются в результате сложного складывания и слияния впяченных участков мембраны. Их функция неизвестна. Сходные структуры описаны у синезеленых водорослей и в клетках грибов (хотя последние относятся к эукариотам).
Вопрос 20. Эндоплазматический ретикулум (ЭР)
1. Характеристика ЭР
Трубчатые или уплотненные цистерны ЭР пронизывают всю цитоплазму и окружают клеточное ядро, образуя ядерную оболочку. Пузыревидные расширения достигают 100 нм в диаметре. Многие или даже все цистерны связаны между собой и с ядерной оболочкой, а их внутреннее пространство сообщается с перинуклеарным пространством. У растений трубчатые цистерны проходят сквозь клеточную стенку в соседние клетки (десмотубулы в десмосомах).Цистерны нельзя выделить целиком, так как при гомогенизации они разрушаются до микросом – фрагментов величиной с рибосому. Биохимический анализ ЭР проводят чаще всего на препаратах микросом.
Мембраны цистерн имеют толщину около 6 нм. Составляющие их липиды – главным образом глицерофосфатиды (90–95 %), в частности лецитин (55 %).
2. Гранулярный ЭР
Гранулярный (шероховатый) ЭР густо усеян полисомами, а гладкий (агранулярный) ЭР, состоящий в основном из трубчатых элементов, не связан с ними. Плотные слои цистерн гранулярного ЭР – так называемая эргастоплазма – окрашиваются основными красителями благодаря высокому содержанию нуклеиновых кислот, поэтому скопления этих цистерн видны в световой микроскоп, особенно в клетках, секретирующих белки (в слюнных железах и поджелудочной железе).В гранулярном ЭР происходит синтез определенных белков. Рибосомы, прикрепленные своими большими субчастицами к мембране, проталкивают вновь синтезируемые полипептидные цепи в цистерны, откуда белки выводятся из клетки, чаще всего с помощью трубчатых цистерн гладкого ЭР.
3. Гладкий ЭР
В гладком ЭР протекают различные этапы обмена углеводов, жирных кислот, жиров, терпеноидов и других веществ. Прежде всего это центр синтеза липидов и мембранных стероидов (холестерола) и тем самым начальный пункт течения мембран, т. е. образования и регенерации всей системы эндомембран и плазматической мембраны. В мышечных клетках ЭР, называемый здесь саркоплазматическим ретикулумом, обслуживает двигательную функцию.В быстро растущих животных клетках (эмбриональных, раковых) в цитоплазме и в клеточном ядре встречаются кольчатые мембраны, сходные по структуре с ядерной оболочкой, – короткие и плоские изолированные фрагменты двойной мембраны с порами.
Цистерны ЭР могут «размножаться», синтезируя собственные структурные компоненты. Кроме того, они образуются, по-видимому, и из других мембран (например, цистерн Гольджи) или в результате слияния пузырьков, отшнуровывающихся от других частей ЭР.
Вопрос 21. Система Гольджи
1. Характеристика системы Гольджи
Система Гольджи используется в клетке для образования роста и регенерации плазматической мембраны и для образования экскретов в самом широком смысле (прежде всего углеводов и белков).Диктиосомы – это стопки из 3—12 дискообразных замкнутых цистерн Гольджи (диаметром в большинстве случаев до 0,2–0,5 мкм), от краев которых отшнуровываются пузырьки Гольджи с диаметром около 20 нм. Как правило, их бывает от нескольких сотен до нескольких тысяч на клетку. Более старые цистерны продырявлены; между ними могут появляться тонкие параллельные трубчатые или фибриллярные элементы.
Аппарат Гольджи в зрелых клетках позвоночных представляет собой результат слияния всех диктиосом, часто лежащий вблизи ядра, окрашивающийся и видимый в световой микроскоп. В яйцеклетках, некоторых эмбриональных клетках и во время клеточного деления еще встречаются диктиосомы, но при дифференцировке клеток они сливаются благодаря росту цистерн и их агрегации. Кроме пузырьков, в результате расширения цистерн образуются крупные вакуоли Гольджи.
2. Синтез системы Гольджи
Система Гольджи – производное эндоплазматического ретикулума. На одной стороне стопки цистерн слияние отдельных частей ЭР (пузырьков или фрагментов) ведет к формированию новых цистерн Гольджи. По мере дальнейшего поступления веществ из ЭР с пузырьками (везикулярный поток) или через трубчатые соединения в цистернах образуется секрет и одновременно происходит перестройка мембран: тонкая (до 6 нм) мембрана ЭР превращается в более толстую (8 нм) и более плотную мембрану с иным составом липидов и белков, сходную с плазматической. Липиды поступают из гладкого ЭР, а белки – частью из гранулярного ЭР, а частью – от свободных полисом.Зрелые цистерны на секреторной стороне стопки используются для формирования пузырьков, или вакуолей, Гольджи, заполненных секретом (у быстро работающих диктиосом весь процесс длится от 20 с до 2 мин). Пузырьки Гольджи подходят к плазматической мембране, сливаются с ней, изливают свое содержимое наружу (экзоцитоз), а их мембрана включается в плазматическую мембрану. Аналогичным образом они могут опорожняться и во внутренние компартменты, например в секреторные вакуоли у растений.
Диктиосомы образуются заново из частей ЭР, может быть, умножаются путем деления надвое; последнее весьма спорно.
Цистерны Гольджи активно извлекают моносахариды из основного вещества протоплазмы и синтезируют из них олиго– и полисахариды. У растений таким способом образуются протопектин и гемицеллюлоза для формирования клеточной стенки, реже – целлюлоза, а также полисахаридная слизь. При клеточном делении пузырьки Гольджи скапливаются на новой границе между клетками и сливаются, их содержимое образует первичную клеточную стенку, а мембраны – плазмолемму. Для последующего роста клеточной стенки новые пузырьки Гольджи путем эндоцитоза добавляют к ней свое содержимое.
3. Диктиосомы
У животных система Гольджи синтезирует гликопротеиды и гликолипиды гликокаликса. Гликозилирование начинается в эндоплазматическом ретикулуме; полисахаридные остатки, синтезируемые далее в цистернах Гольджи, выступают во внутреннее пространство этих цистерн, а после экзоцитоза попадают на наружную поверхность плазматической мембраны.«Экспортируемые» белки химически изменяются во внутреннем пространстве цистерн (и пузырьков) Гольджи. Они могут связываться с сахаром или сульфатом, как это происходит в слизистых клетках кишечного эпителия, или активируются в результате отщепления аминокислотных остатков (процессинг), как, например, в случае превращения проинсулина в инсулин в лангергансовых островках поджелудочной железы.
4. Синтез пищеварительных ферментов
В секреторных клетках поджелудочной железы пищеварительные ферменты (в частности, трипсиноген) синтезируются в эргастоплазме. Они появляются там между цистернами как мелкие просекреторные гранулы, не одетые мембраной. Затем они (возможно, в вакуолях Гольджи) попадают в аппарат Гольджи и сливаются там в очень крупные (0,5–1,5 мкм) секреторные пузырьки (зимогеновые гранулы). Последние при определенной стимуляции выбрасывают свое содержимое из клетки, а их мембраны сливаются друг с другом и с плазматической мембраной. Сходным образом вырабатываются и выделяются амилаза в слюнных железах, пептидные гормоны в гипофизе, коллаген в ряде тканей млекопитающих.Аппарат Гольджи участвует также в образовании белков молока в молочных железах, желчи в печени, веществ хрусталика, зубной эмали и т. п.
Вопрос 22. Пузырьки, эндо– и эктоцитоз
1. Эндоцитоз
Пузырьки – это округлые или овальные образования с одиночной мембраной. Они либо имеют гладкую стенку, либо покрыты снаружи волокнистой оболочкой из белка клатрина (окаймленные пузырьки).Эндоцитоз – это образование пузырьков путем выпячивания плазматической мембраны при поглощении твердых частиц (фагоцитоз) или растворенных веществ (пиноцитоз). Возникающие при этом гладкие или окаймленные эндоцитозные пузырьки называют также фагосомами или пиносомами. Путем эндоцитоза осуществляются, во-первых, питание (яйцеклетки поглощают таким способом желточные белки, фагосомами являются пищеварительные вакуоли простейших), во-вторых, защитные и иммунные реакции (лейкоциты поглощают чужеродные частицы и иммуноглобулины), в-третьих, транспорт (почечные канальцы всасывают белки из первичной мочи). Избирательный эндоцитоз определенных веществ (желточных белков, иммуноглобулинов и т. п.) происходит при контакте этих веществ с субстрат-специфическими рецепторными участками на плазматической мембране.
Материалы, попадающие в клетку путем эндоцитоза, расщепляются («перевариваются»), накапливаются (например, желточные белки) или снова выводятся с противоположной стороны клетки путем экзоцитоза.
2. Экзоцитоз
Экзоцитоз – процесс, противоположный эндоцитозу: различные пузырьки (например, из эндоплазматического ретикулума, аппарата Гольджи, эндоцитозные пузырьки, лизосомы) сливаются с плазматической мембраной, освобождая свое содержимое наружу. При этом мембрана пузырька может либо встраиваться в плазматическую мембрану, либо в форме пузырька возвращаться в цитоплазму. У растений экзоцитоз широко распространен; в отдельных случаях был обнаружен фагоцитоз, а пиноцитоз, по-видимому, не встречается.
Вопрос 23. Лизосомы
1. Характеристика лизосом
Лизосомы осуществляют внутриклеточное переваривание. Это пузырьки величиной до 2 мкм, бесструктурные или содержащие полупереваренный материал. Их главный наиболее характерный фермент – кислая фосфатаза, но, кроме того, в них имеется свыше 30 ферментов, осуществляющих гидролитическое расщепление белков, нуклеиновых кислот, углеводов и липидов.2. Гетерофагия
Гетерофагия – это расщепление чужеродного и поглощенного путем эндоцитоза материала в гетеролизосомах (фаголизосомах), а аутофагия – расщепление в аутосомах (цитолизосомах) собственных материалов, например, запасных веществ, а также макромолекул или органелл, утративших функциональную активность. Аутолиз – самопереваривание клеток после разрушения мембран лизосом, вызванного патологическими изменениями или старением.3. Активность лизосом
Первичные лизосомы еще неактивны, они не содержат перевариваемого субстрата. Это чаще всего мелкие, гладкостенные, реже – окаймленные пузырьки. Они отшнуровываются от цистерн Гольджи. Ферменты образуются в гранулярном ЭР и собираются чаще всего в цистернах Гольджи. Вторичные лизосомы обладают гидролитической активностью.Они образуются различными способами из первичных лизосом после поглощения субстрата:
• первичные лизосомы могут активно поглощать
макромолекулы из окружающей цитоплазмы (аутофагия);
• первичные лизосомы могут сливаться с эндоцитозными пузырьками (гетерофагия). Возможно повторение этого процесса – слияние вторичных лизосом с новыми эндоцитозными пузырьками, доставляющими субстрат, и с новыми первичными лизосомами, добавляющими свои ферменты;
Конец бесплатного ознакомительного фрагмента