Страница:
Через сфинктер Одди небольшое количество желчи выходит в двенадцатиперстную кишку. Этот период первичной реакции желчевыделительного аппарата длится 7-10 мин. За ним следует период опорожнения желчного пузыря, во время которого сокращения пузыря чередуются с расслаблением, и через открытый сфинктер Одди переходит в двенадцатиперстную кишку сначала в основном пузырная желчь, а позже – печеночная. Длительность латентного и эвакуаторного периодов, количество выделенной желчи различны и зависят от вида принятой пищи. Сильными возбудителями желчевыделения являются яичные желтки, молоко, мясо и жиры. Через 3–6 ч после приема пищи наблюдают понижение желчевыделения, затухание сократительной деятельности желчного пузыря, в котором начинает вновь депонироваться печеночная желчь.
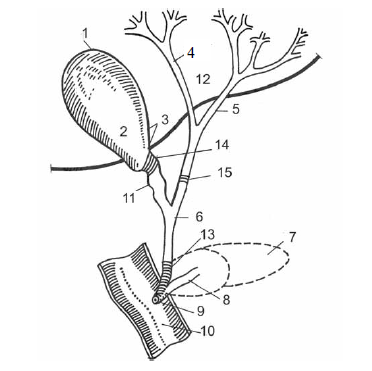
Рис. 2.6. Внепеченочные желчные пути и сфинктеры (схема)(по: Г. И. Косицкий, 1985).
1 – дно желчного пузыря; 2 – желчный пузырь; 3 – шейка пузыря; 4, 5 – ветви печеночного протока; 6 – общий желчный проток; 7 – поджелудочная железа; 8 – проток поджелудочной железы; 9 – сосок двенадцатиперстной кишки (фатеров); 10 – двенадцатиперстная кишка; 11 – пузырный проток; 12 – печень; 13 – сфинктер Одци; 14 – сфинктер Люткенса; 15 – сфинктер Мирицци.
Недостаточное поступление желчи в кишечник называется гипохолией, полное прекращение ее поступления – ахолией. Эти явления возможны при закупорке или сдавлении общего желчного протока, при нарушении желчеобразовательной функции печени. При ахолии резко страдает переваривание и всасывание жира, так как липаза поджелудочного сока в отсутствие желчи малоактивна, а жиры не эмульгированы и контакт их с липолитическим ферментом затруднен. При недостатке желчи страдает всасывание жирных кислот, холестерина, жирорастворимых витаминов. Вследствие недостаточного переваривания и всасывания жира развивается стеаторея – избыточное содержание жира в испражнениях. С калом при этом выводится до 70–80 % жира. В кишечнике нерасщепленный жир обволакивает пищевые массы и затрудняет действие на них панкреатических протеолитических ферментов и α-амилазы. Гидролитическая активность ферментов при недостатке желчи падает, так как желчь принимает участие в нейтрализации кислого желудочного содержимого и способствует поддержанию в двенадцатиперстной кишке щелочной среды, являющейся оптимальной для ферментов дуоденального сока.
Наконец, желчные кислоты стимулируют сорбционные свойства кишечного эпителия, поэтому при их дефиците уменьшается адсорбция кишечными клетками ферментов из химуса и нарушается мембранное пищеварение. Таким образом, при гипо– и ахолии нарушаются также начальные и промежуточные этапы переваривания белков и углеводов в тонкой кишке.
Вместе с неусвоенными жирами из кишечника выводятся и жирорастворимые витамины. Развиваются гиповитаминозы, в частности, нередко понижается свертываемость крови в связи с дефицитом витамина К.
При гипо– и ахолии ослабляется перистальтика кишечника, что в сочетании с выпадением бактерицидного действия желчи ведет к усилению в кишечнике процессов брожения и гниения, к метеоризму – развивается дисбактериоз кишечника. В результате наступает интоксикация организма.
Рефлекторные влияния на желчевыделительный аппарат осуществляются условно– и безусловно-рефлекторно с участием многочисленных рефлексогенных зон, в том числе рецепторов полости рта, желудка и двенадцатиперстной кишки. Велика роль гуморальной регуляции желчевыделения. Большую роль в качестве гуморального стимулятора желчевыделения играет холецистокинин, вызывающий сокращение желчного пузыря. Сокращения желчного пузыря, хотя и слабые, вызывают также гастрин, секретин, бомбезин. Тормозят сокращения желчного пузыря глюкагон, кальцитонин, антихолецистокинин, ВИП,ПП.
Глава 3 Базисные сведения о важнейших видах обмена веществ
Белки
Углеводы
Рис. 2.6. Внепеченочные желчные пути и сфинктеры (схема)(по: Г. И. Косицкий, 1985).
1 – дно желчного пузыря; 2 – желчный пузырь; 3 – шейка пузыря; 4, 5 – ветви печеночного протока; 6 – общий желчный проток; 7 – поджелудочная железа; 8 – проток поджелудочной железы; 9 – сосок двенадцатиперстной кишки (фатеров); 10 – двенадцатиперстная кишка; 11 – пузырный проток; 12 – печень; 13 – сфинктер Одци; 14 – сфинктер Люткенса; 15 – сфинктер Мирицци.
Недостаточное поступление желчи в кишечник называется гипохолией, полное прекращение ее поступления – ахолией. Эти явления возможны при закупорке или сдавлении общего желчного протока, при нарушении желчеобразовательной функции печени. При ахолии резко страдает переваривание и всасывание жира, так как липаза поджелудочного сока в отсутствие желчи малоактивна, а жиры не эмульгированы и контакт их с липолитическим ферментом затруднен. При недостатке желчи страдает всасывание жирных кислот, холестерина, жирорастворимых витаминов. Вследствие недостаточного переваривания и всасывания жира развивается стеаторея – избыточное содержание жира в испражнениях. С калом при этом выводится до 70–80 % жира. В кишечнике нерасщепленный жир обволакивает пищевые массы и затрудняет действие на них панкреатических протеолитических ферментов и α-амилазы. Гидролитическая активность ферментов при недостатке желчи падает, так как желчь принимает участие в нейтрализации кислого желудочного содержимого и способствует поддержанию в двенадцатиперстной кишке щелочной среды, являющейся оптимальной для ферментов дуоденального сока.
Наконец, желчные кислоты стимулируют сорбционные свойства кишечного эпителия, поэтому при их дефиците уменьшается адсорбция кишечными клетками ферментов из химуса и нарушается мембранное пищеварение. Таким образом, при гипо– и ахолии нарушаются также начальные и промежуточные этапы переваривания белков и углеводов в тонкой кишке.
Вместе с неусвоенными жирами из кишечника выводятся и жирорастворимые витамины. Развиваются гиповитаминозы, в частности, нередко понижается свертываемость крови в связи с дефицитом витамина К.
При гипо– и ахолии ослабляется перистальтика кишечника, что в сочетании с выпадением бактерицидного действия желчи ведет к усилению в кишечнике процессов брожения и гниения, к метеоризму – развивается дисбактериоз кишечника. В результате наступает интоксикация организма.
Рефлекторные влияния на желчевыделительный аппарат осуществляются условно– и безусловно-рефлекторно с участием многочисленных рефлексогенных зон, в том числе рецепторов полости рта, желудка и двенадцатиперстной кишки. Велика роль гуморальной регуляции желчевыделения. Большую роль в качестве гуморального стимулятора желчевыделения играет холецистокинин, вызывающий сокращение желчного пузыря. Сокращения желчного пузыря, хотя и слабые, вызывают также гастрин, секретин, бомбезин. Тормозят сокращения желчного пузыря глюкагон, кальцитонин, антихолецистокинин, ВИП,ПП.
Глава 3 Базисные сведения о важнейших видах обмена веществ
Метаболизм основных видов нутриентов в норме и при патологии определяет потребности организма в них и пути их введения. Поэтому знания о процессах ассимиляции различных питательных веществ являются основой диетической терапии.
Белки
Белки – сложные азотсодержащие биополимеры, мономерами которых служат аминокислоты (органические соединения, содержащие карбоксильную и аминогруппы). Их биологическая роль многообразна. Белки выполняют в организме пластические, каталитические, гормональные, транспортные и другие функции, а также обеспечивают специфичность. Значение белкового компонента питания заключается, прежде всего, в том, что он служит источником аминокислот.
Аминокислоты делятся на эссенциальные и неэссенциальные в зависимости от того, возможно ли их образование в организме из предшественников. К незаменимым аминокислотам относятся гистидин, лейцин, изолейцин, лизин, метионин, фенилаланин, триптофан и валин, а также цистеин и тирозин, синтезируемые соответственно из метионина и фенилаланина. 9 заменимых аминокислот (аланин, аргинин, аспаргиновая и глутаминовая кислоты, глутамин, глицин, пролин и серии) могут отсутствовать в рационе, так как способны образовываться из других веществ. В организме также существуют аминокислоты, которые продуцируются путем модификации боковых цепей вышеперечисленных (например, компонент коллагена – гидроксипролин – и сократительных белков мышц – 3-метилгистидин).
Большинство аминокислот имеют изомеры (D– и L-формы), из которых только L-формы входят в состав белков человеческого организма. D-формы могут участвовать в метаболизме, превращаясь в L-формы, однако утилизируются гораздо менее эффективно.
По химическому строению аминокислоты делятся на двухосновные, двухкислотные и нейтральные с алифатическими и ароматическими боковыми цепями, что имеет важное значение для их транспорта, поскольку каждый класс аминокислот обладает специфическими переносчиками. Аминокислоты с аналогичным строением обычно вступают в сложные, часто конкурентные взаимоотношения.
Так, ароматические аминокислоты (фенилаланин, тирозин и триптофан) близкородственны между собой. Хотя фенилаланин является незаменимой, а тирозин синтезируемой из него заменимой аминокислотой, наличие тирозина в рационе как будто бы «сберегает» фенилаланин. Если фенилаланина недостаточно, или его метаболизм нарушен (например, при дефиците витамина С) – тирозин становится незаменимой аминокислотой. Подобные взаимоотношения характерны и для серосодержащих аминокислот: незаменимой – метионина, и образующегося из него цистеина.
Приведем другой пример. Триптофан в ходе превращений, для которых необходим витамин В6 (пиридоксин), включается в структуру НАД и НАДФ, то есть дублирует роль ниацина. Приблизительно половина обычной потребности в ниацине удовлетворяется за счет триптофана: 1 мг ниацина пищи эквивалентен 60 мг триптофана. Поэтому состояние пеллагры может развиваться не только при недостатке витамина РР в рационе, но и при нехватке триптофана или нарушении его обмена, в том числе вследствие дефицита пиридоксина.
Аминокислоты также делятся на глюкогенные и кетогенные, в зависимости от того, могут ли они при определенных условиях становиться предшественниками глюкозы или кетоновых тел (табл. 3.1).
Таблица 3.1
Классификация аминокислот

Примечания: Г – глюкогенные, К – кетогенные аминокислоты; * – гистидин незаменим у детей до года; ** – «условно-незаменимые» аминокислоты (могут синтезироваться из фенилаланина и метионина).
Поступление азотсодержащих веществ с пищей происходит в основном за счет белка и, в менее значимых количествах, свободных аминокислот и других соединений. В животной пище основное количество азота содержится в виде белка. В продуктах растительного происхождения большая часть азота представлена небелковыми соединениями, а также содержится множество аминокислот, которые не встречаются в организме человека и зачастую не могут метаболизироваться им.
Человек не нуждается в поступлении с пищей нуклеиновых кислот. Пуриновые и пиримидиновые основания синтезируются в печени из аминокислот, а избыток этих оснований, поступивший с пищей, выводится в виде мочевой кислоты. В синтезе пиримидиновых колец принимает участие витамин В12, для образования пуриновых структур необходима фолиевая кислота. Именно поэтому дефицит этих нутриентов отражается, прежде всего, на органе с высоким уровнем пролиферации, где идет наиболее интенсивный синтез нуклеиновых кислот – кроветворной ткани.
Обычный (но не оптимальный) ежедневный прием белка у среднестатистического человека составляет приблизительно 100 г. К ним присоединяется примерно 70 г белка, секретируемого в полость желудочно-кишечного тракта. Из этого количества абсорбируется около 160 г. Самим организмом в сутки синтезируется в среднем 240–250 г белка. Такая разница между поступлением и эндогенным преобразованием свидетельствует об активности процессов ресинтеза (рис. 3.1).
Для здорового человека характерно состояние азотного равновесия, когда потери белка (с мочой, калом, эпидермисом и т. п.) соответствуют его количеству, поступившему с пищей. При преобладании катаболических процессов возникает отрицательный азотный баланс, который характерен для низкого потребления азотсодержащих веществ (низкобелковые рационы, голодание, нарушение абсорбции белка) и многих патологических процессов, вызывающих интенсификацию распада (опухоли, ожоговая болезнь и т. п.). При доминировании синтетических процессов количество вводимого азота преобладает над его выведением и возникает положительный азотный баланс, характерный для детей, беременных женщин и реконвалесцентов после тяжелых заболеваний.
После прохождения энтерального барьера белки поступают в кровь в виде свободных аминокислот. Следует отметить, что клетки слизистой оболочки желудочно-кишечного тракта могут метаболизировать некоторые аминокислоты (в том числе глутаминовую кислоту и аспаргиновую кислоту в аланин). Способность энтероцитов видоизменять эти аминокислоты, возможно, позволяет избежать токсического эффекта при их избыточном введении.
Аминокислоты, как поступившие в кровь при переваривании белка, так и синтезированные в клетках, в крови образуют постоянно обновляющийся свободный пул аминокислот, который составляет около 100 г.
75 % аминокислот, находящихся в системной циркуляции, представлены аминокислотами с ветвящимися цепями (лейцином, изолейцином и валином). Из мышечной ткани в кровоток выделяются аланин, который является основным предшественником синтеза глюкозы, и глутамин.
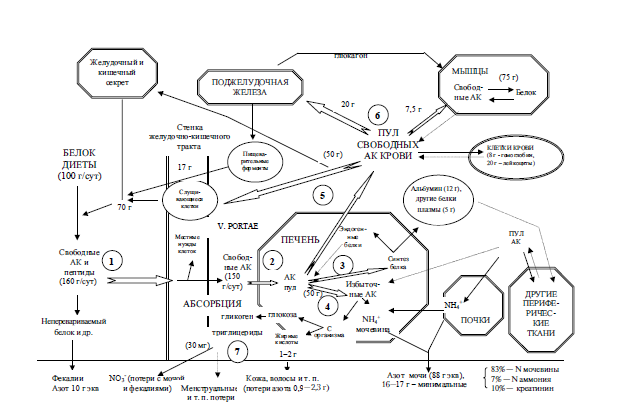
Рис. 3.1. Метаболизм белка (по: Н. N. Munro, М. Munro, 1988, с изменениями).
Примечания: АК – аминокислоты. У человека с массой тела 62,5 кг содержание общее белка – 10,9 кг(17,5 %), 240 г белка ежедневно синтезируется и распадается. 1 – абсорбция свободных аминокислот и пептидов после переваривания; 2 – поступление аминокислот в печень; 3 – синтез белков печени и плазмы, в том числе альбумина; 4 – катаболизм излишних аминокислот; 5 – распределение аминокислот в состоянии покоя; 6 – поступление в мышцы, поджелудочную железу, эпителиальные клетки; 7 – экскреция азота в различных формах.
Многие свободные аминокислоты подвергаются трансформации в печени. Часть свободного пула инкорпорируется в белки организма и при их катаболизме вновь поступает в кровоток. Другие непосредственно подвергаются катаболическим реакциям. Некоторые свободные аминокислоты используются для синтеза новых азотсодержащих соединений (пурина, креатинина, адреналина) и в дальнейшем деградируют, не возвращаясь в свободный пул, в специфичные продукты распада.
Печень обеспечивает постоянство содержания различных аминокислот в крови. Она утилизирует примерно 1/ всех аминокислот, поступающих в организм, что позволяет предотвратить скачки в их концентрации в зависимости от питания. Первостепенная роль печени в азотном и других видах обмена обеспечивается ее анатомическим расположением – продукты переваривания попадают по воротной вене непосредственно в этот орган. Кроме того, печень непосредственно связана с экскреторной системой – билиарным трактом, что позволяет выводить некоторые соединения в составе желчи. Гепатоциты – единственные клетки, обладающие полным набором ферментов, участвующих в аминокислотном обмене. Здесь выполняются все основные процессы азотного метаболизма: распад аминокислот для выработки энергии и обеспечения глюконеогенеза, образование заменимых аминокислот и нуклеиновых кислот, обезвреживание аммиака и других конечных продуктов. Печень является основным местом деградации большинства незаменимых аминокислот (за исключением аминокислот с ветвящимися цепями).
Синтез азотсодержащих соединений (белка и нуклеиновых кислот) в печени весьма чувствителен к поступлению их предшественников из пищи. После каждого приема пищи наступает период повышенного внутрипеченочного синтеза белков, в том числе альбумина. Аналогичное усиление синтетических процессов происходит и в мышцах. Эти реакции связаны, прежде всего, с действием инсулина, который секретируется в ответ на введение аминокислот и/или глюкозы. Некоторые аминокислоты (аргинин и аминокислоты с ветвящимися цепями) усиливают продукцию инсулина в большей степени, чем остальные. Другие (аспаргин, глицин, серии, цистеин) стимулируют секрецию глюкагона, который усиливает утилизацию аминокислот печенью и воздействует на ферменты глюконеогенеза и аминокислотного катаболизма. Благодаря этим механизмам происходит снижение уровня аминокислот в крови после поступления их с пищей. Действие инсулина наиболее выражено для аминокислот, содержащихся в кровотоке в свободном виде (аминокислоты с ветвящимися цепями), и малозначимо для тех, которые транспортируются в связанном (триптофан). Обратное инсулину влияние на белковый метаболизм оказывают глюкокортикостероиды.
Печень обладает повышенной скоростью синтеза и распада белков, по сравнению с другими тканями организма (кроме поджелудочной железы). Это позволяет ей синтезировать «на экспорт», а также быстро обеспечивать лабильный резерв аминокислот в период недостаточного питания за счет распада собственных белков. Особенность внутрипеченочного белкового синтеза заключается в том, что он усиливается под действием гормонов, которые в других тканях производят катаболический эффект. Так, при голодании белки мышц, для обеспечения организма энергией, подвергаются распаду, а в печени одновременно усиливается синтез белков, являющихся ферментами глюконеогенеза и мочеви-нообразования.
Прием пищи, содержащей избыток белка, приводит к интенсификации синтеза в печени и в мышцах, образованию избыточных количеств альбумина и деградации излишка аминокислот до предшественников глюкозы и липидов. Глюкоза и триглицериды утилизируются как горючее или депонируются, а альбумин становится временным хранилищем аминокислот и средством их транспортировки в периферические ткани.
При голодании уровень альбумина прогрессивно снижается, а при последующей нормализации поступления белка медленно восстанавливается. Поэтому, хотя альбумин и является показателем белковой недостаточности, он низкочувствителен и не реагирует оперативно на изменения в питании (см. главу «Оценка состояния питания»).
7 из 10 эссенциальных аминокислот деградируют в печени – либо образуя мочевину, либо впоследствии используясь в глюконеогенезе. Мочевина преимущественно выделяется с мочой, но часть ее поступает в просвет кишечника, где подвергается уреазному воздействию микрофлоры. Аминокислоты с ветвящимися цепями катаболизируются в основном в почках, мышцах и головном мозге.
Мышцы синтезируют ежедневно 75 г белка. У среднего человека они содержат 40 % от всего белка организма. Хотя белковый метаболизм происходит здесь несколько медленнее, чем в других тканях, мышечный белок представляет собой самый большой эндогенный аминокислотный резерв, который при голодании может использоваться для глюконеогенеза.
В отсутствие пищи синтез альбумина и мышечного белка замедляется, но продолжается деградация аминокислот. Поэтому на начальном этапе голодания мышцы теряют аминокислоты, которые идут на энергетические нужды. В дальнейшем организм адаптируется к отсутствию новых поступлений аминокислот (снижается потребность в зависящем от белка глюконеогенезе за счет использования энергетического потенциала кетоновых тел) и потеря белка мускулатуры уменьшается.
Мышцы являются основной мишенью воздействия инсулина: здесь под его влиянием усиливается поступление аминокислот, увеличивается синтез мышечного белка и снижается распад.
В процессе превращений в мышцах образуются аланин и глутамин, их условно можно считать транспортными формами азота. Аланин непосредственно из мышц попадает в печень, а глутамин вначале поступает в кишечник, где частично превращается в аланин. Поскольку в печени из аланина происходит синтез глюкозы, частично обеспечивающий мышцу энергией, получающийся кругооборот получил название глюкозо-аланинового цикла.
К азотсодержащим веществам мышц также относятся высокоэнергетичный креатин-фосфат и продукт его деградации креатинин. Экскреция креатинина обычно расматривается как мера мышечной массы. Однако это соединение может поступать в организм с высокобелковой пищей и влиять на результаты исследования содержания его в моче. Продукт распада миофибриллярных белков – 3-метилгистидин экскретируется с мочой в течение короткого времени и является достаточно точным показателем скорости распада в мышцах – при мышечном истощении скорость его выхода пропорционально снижается.
Почки не только выводят конечные продукты азотного распада (мочевину, креатинин и др.), но и являются дополнительным местом ресинтеза глюкозы из аминокислот, а также регулируют образование аммиака, компенсируя избыток ионов водорода в крови. Глюконеогенез и функционирование кислотно-щелочной регуляции тесно скоординированы, поскольку субстраты этих процессов появляются при дезаминировании аминокислот: углерод для синтеза глюкозы и азот – для аммиака. Существует даже мнение, что именно производство глюкозы является основной реакцией почек на ацидоз, а образование аммиака происходит вторично.
Для нервной ткани характерны более высокие концентрации аминокислот, чем в плазме. Это позволяет обеспечить мозг достаточным количеством ароматических аминокислот, являющихся предшественниками нейромедиаторов. Некоторые заменимые аминокислоты, такие как глутамат (из которого при участии пиридоксина образуется ГАМ К) и аспартат, также обладают влиянием на возбудимость нервной ткани. Их концентрация здесь высока, при этом заменимые аминокислоты способны синтезироваться и на месте.
Специфическую роль играет триптофан, являющийся предшественником серотонина. Именно с повышением концентрации триптофана (а, следовательно, и серотонина) связана сонливость после еды. Такой эффект особенно выражен при приеме больших количеств триптофана совместно с углеводной пищей. Повышенная секреция инсулина снижает уровень в крови аминокислот с ветвящимися цепями, которые при преодолении барьера кровь – мозг обладают конкурентными взаимоотношениями с ароматическими, но в то же время не оказывает влияния на концентрацию связанного с альбумином триптофана. Благодаря подобным эффектам препараты триптофана могут использоваться в психиатрической практике.
Ограничение ароматических аминокислот в рационе, в связи с их влиянием на центральную нервную систему, имеет профилактическое значение при ведении пациентов с печеночной энцефалопатией. Элементные аминокислотные диеты с преимущественным содержанием лейцина, изолейцина, валина и аргинина помогают избежать развития белковой недостаточности у гепатологических больных, и в то же время не приводят к возникновению печеночной комы.
Основные пластические функции протеиногенных аминокислот перечислены в табл. 3.2.
Таблица 3.2
Основные функции аминокислот

Аминокислоты делятся на эссенциальные и неэссенциальные в зависимости от того, возможно ли их образование в организме из предшественников. К незаменимым аминокислотам относятся гистидин, лейцин, изолейцин, лизин, метионин, фенилаланин, триптофан и валин, а также цистеин и тирозин, синтезируемые соответственно из метионина и фенилаланина. 9 заменимых аминокислот (аланин, аргинин, аспаргиновая и глутаминовая кислоты, глутамин, глицин, пролин и серии) могут отсутствовать в рационе, так как способны образовываться из других веществ. В организме также существуют аминокислоты, которые продуцируются путем модификации боковых цепей вышеперечисленных (например, компонент коллагена – гидроксипролин – и сократительных белков мышц – 3-метилгистидин).
Большинство аминокислот имеют изомеры (D– и L-формы), из которых только L-формы входят в состав белков человеческого организма. D-формы могут участвовать в метаболизме, превращаясь в L-формы, однако утилизируются гораздо менее эффективно.
По химическому строению аминокислоты делятся на двухосновные, двухкислотные и нейтральные с алифатическими и ароматическими боковыми цепями, что имеет важное значение для их транспорта, поскольку каждый класс аминокислот обладает специфическими переносчиками. Аминокислоты с аналогичным строением обычно вступают в сложные, часто конкурентные взаимоотношения.
Так, ароматические аминокислоты (фенилаланин, тирозин и триптофан) близкородственны между собой. Хотя фенилаланин является незаменимой, а тирозин синтезируемой из него заменимой аминокислотой, наличие тирозина в рационе как будто бы «сберегает» фенилаланин. Если фенилаланина недостаточно, или его метаболизм нарушен (например, при дефиците витамина С) – тирозин становится незаменимой аминокислотой. Подобные взаимоотношения характерны и для серосодержащих аминокислот: незаменимой – метионина, и образующегося из него цистеина.
Приведем другой пример. Триптофан в ходе превращений, для которых необходим витамин В6 (пиридоксин), включается в структуру НАД и НАДФ, то есть дублирует роль ниацина. Приблизительно половина обычной потребности в ниацине удовлетворяется за счет триптофана: 1 мг ниацина пищи эквивалентен 60 мг триптофана. Поэтому состояние пеллагры может развиваться не только при недостатке витамина РР в рационе, но и при нехватке триптофана или нарушении его обмена, в том числе вследствие дефицита пиридоксина.
Аминокислоты также делятся на глюкогенные и кетогенные, в зависимости от того, могут ли они при определенных условиях становиться предшественниками глюкозы или кетоновых тел (табл. 3.1).
Таблица 3.1
Классификация аминокислот
Примечания: Г – глюкогенные, К – кетогенные аминокислоты; * – гистидин незаменим у детей до года; ** – «условно-незаменимые» аминокислоты (могут синтезироваться из фенилаланина и метионина).
Поступление азотсодержащих веществ с пищей происходит в основном за счет белка и, в менее значимых количествах, свободных аминокислот и других соединений. В животной пище основное количество азота содержится в виде белка. В продуктах растительного происхождения большая часть азота представлена небелковыми соединениями, а также содержится множество аминокислот, которые не встречаются в организме человека и зачастую не могут метаболизироваться им.
Человек не нуждается в поступлении с пищей нуклеиновых кислот. Пуриновые и пиримидиновые основания синтезируются в печени из аминокислот, а избыток этих оснований, поступивший с пищей, выводится в виде мочевой кислоты. В синтезе пиримидиновых колец принимает участие витамин В12, для образования пуриновых структур необходима фолиевая кислота. Именно поэтому дефицит этих нутриентов отражается, прежде всего, на органе с высоким уровнем пролиферации, где идет наиболее интенсивный синтез нуклеиновых кислот – кроветворной ткани.
Обычный (но не оптимальный) ежедневный прием белка у среднестатистического человека составляет приблизительно 100 г. К ним присоединяется примерно 70 г белка, секретируемого в полость желудочно-кишечного тракта. Из этого количества абсорбируется около 160 г. Самим организмом в сутки синтезируется в среднем 240–250 г белка. Такая разница между поступлением и эндогенным преобразованием свидетельствует об активности процессов ресинтеза (рис. 3.1).
Для здорового человека характерно состояние азотного равновесия, когда потери белка (с мочой, калом, эпидермисом и т. п.) соответствуют его количеству, поступившему с пищей. При преобладании катаболических процессов возникает отрицательный азотный баланс, который характерен для низкого потребления азотсодержащих веществ (низкобелковые рационы, голодание, нарушение абсорбции белка) и многих патологических процессов, вызывающих интенсификацию распада (опухоли, ожоговая болезнь и т. п.). При доминировании синтетических процессов количество вводимого азота преобладает над его выведением и возникает положительный азотный баланс, характерный для детей, беременных женщин и реконвалесцентов после тяжелых заболеваний.
После прохождения энтерального барьера белки поступают в кровь в виде свободных аминокислот. Следует отметить, что клетки слизистой оболочки желудочно-кишечного тракта могут метаболизировать некоторые аминокислоты (в том числе глутаминовую кислоту и аспаргиновую кислоту в аланин). Способность энтероцитов видоизменять эти аминокислоты, возможно, позволяет избежать токсического эффекта при их избыточном введении.
Аминокислоты, как поступившие в кровь при переваривании белка, так и синтезированные в клетках, в крови образуют постоянно обновляющийся свободный пул аминокислот, который составляет около 100 г.
75 % аминокислот, находящихся в системной циркуляции, представлены аминокислотами с ветвящимися цепями (лейцином, изолейцином и валином). Из мышечной ткани в кровоток выделяются аланин, который является основным предшественником синтеза глюкозы, и глутамин.
Рис. 3.1. Метаболизм белка (по: Н. N. Munro, М. Munro, 1988, с изменениями).
Примечания: АК – аминокислоты. У человека с массой тела 62,5 кг содержание общее белка – 10,9 кг(17,5 %), 240 г белка ежедневно синтезируется и распадается. 1 – абсорбция свободных аминокислот и пептидов после переваривания; 2 – поступление аминокислот в печень; 3 – синтез белков печени и плазмы, в том числе альбумина; 4 – катаболизм излишних аминокислот; 5 – распределение аминокислот в состоянии покоя; 6 – поступление в мышцы, поджелудочную железу, эпителиальные клетки; 7 – экскреция азота в различных формах.
Многие свободные аминокислоты подвергаются трансформации в печени. Часть свободного пула инкорпорируется в белки организма и при их катаболизме вновь поступает в кровоток. Другие непосредственно подвергаются катаболическим реакциям. Некоторые свободные аминокислоты используются для синтеза новых азотсодержащих соединений (пурина, креатинина, адреналина) и в дальнейшем деградируют, не возвращаясь в свободный пул, в специфичные продукты распада.
Печень обеспечивает постоянство содержания различных аминокислот в крови. Она утилизирует примерно 1/ всех аминокислот, поступающих в организм, что позволяет предотвратить скачки в их концентрации в зависимости от питания. Первостепенная роль печени в азотном и других видах обмена обеспечивается ее анатомическим расположением – продукты переваривания попадают по воротной вене непосредственно в этот орган. Кроме того, печень непосредственно связана с экскреторной системой – билиарным трактом, что позволяет выводить некоторые соединения в составе желчи. Гепатоциты – единственные клетки, обладающие полным набором ферментов, участвующих в аминокислотном обмене. Здесь выполняются все основные процессы азотного метаболизма: распад аминокислот для выработки энергии и обеспечения глюконеогенеза, образование заменимых аминокислот и нуклеиновых кислот, обезвреживание аммиака и других конечных продуктов. Печень является основным местом деградации большинства незаменимых аминокислот (за исключением аминокислот с ветвящимися цепями).
Синтез азотсодержащих соединений (белка и нуклеиновых кислот) в печени весьма чувствителен к поступлению их предшественников из пищи. После каждого приема пищи наступает период повышенного внутрипеченочного синтеза белков, в том числе альбумина. Аналогичное усиление синтетических процессов происходит и в мышцах. Эти реакции связаны, прежде всего, с действием инсулина, который секретируется в ответ на введение аминокислот и/или глюкозы. Некоторые аминокислоты (аргинин и аминокислоты с ветвящимися цепями) усиливают продукцию инсулина в большей степени, чем остальные. Другие (аспаргин, глицин, серии, цистеин) стимулируют секрецию глюкагона, который усиливает утилизацию аминокислот печенью и воздействует на ферменты глюконеогенеза и аминокислотного катаболизма. Благодаря этим механизмам происходит снижение уровня аминокислот в крови после поступления их с пищей. Действие инсулина наиболее выражено для аминокислот, содержащихся в кровотоке в свободном виде (аминокислоты с ветвящимися цепями), и малозначимо для тех, которые транспортируются в связанном (триптофан). Обратное инсулину влияние на белковый метаболизм оказывают глюкокортикостероиды.
Печень обладает повышенной скоростью синтеза и распада белков, по сравнению с другими тканями организма (кроме поджелудочной железы). Это позволяет ей синтезировать «на экспорт», а также быстро обеспечивать лабильный резерв аминокислот в период недостаточного питания за счет распада собственных белков. Особенность внутрипеченочного белкового синтеза заключается в том, что он усиливается под действием гормонов, которые в других тканях производят катаболический эффект. Так, при голодании белки мышц, для обеспечения организма энергией, подвергаются распаду, а в печени одновременно усиливается синтез белков, являющихся ферментами глюконеогенеза и мочеви-нообразования.
Прием пищи, содержащей избыток белка, приводит к интенсификации синтеза в печени и в мышцах, образованию избыточных количеств альбумина и деградации излишка аминокислот до предшественников глюкозы и липидов. Глюкоза и триглицериды утилизируются как горючее или депонируются, а альбумин становится временным хранилищем аминокислот и средством их транспортировки в периферические ткани.
При голодании уровень альбумина прогрессивно снижается, а при последующей нормализации поступления белка медленно восстанавливается. Поэтому, хотя альбумин и является показателем белковой недостаточности, он низкочувствителен и не реагирует оперативно на изменения в питании (см. главу «Оценка состояния питания»).
7 из 10 эссенциальных аминокислот деградируют в печени – либо образуя мочевину, либо впоследствии используясь в глюконеогенезе. Мочевина преимущественно выделяется с мочой, но часть ее поступает в просвет кишечника, где подвергается уреазному воздействию микрофлоры. Аминокислоты с ветвящимися цепями катаболизируются в основном в почках, мышцах и головном мозге.
Мышцы синтезируют ежедневно 75 г белка. У среднего человека они содержат 40 % от всего белка организма. Хотя белковый метаболизм происходит здесь несколько медленнее, чем в других тканях, мышечный белок представляет собой самый большой эндогенный аминокислотный резерв, который при голодании может использоваться для глюконеогенеза.
В отсутствие пищи синтез альбумина и мышечного белка замедляется, но продолжается деградация аминокислот. Поэтому на начальном этапе голодания мышцы теряют аминокислоты, которые идут на энергетические нужды. В дальнейшем организм адаптируется к отсутствию новых поступлений аминокислот (снижается потребность в зависящем от белка глюконеогенезе за счет использования энергетического потенциала кетоновых тел) и потеря белка мускулатуры уменьшается.
Мышцы являются основной мишенью воздействия инсулина: здесь под его влиянием усиливается поступление аминокислот, увеличивается синтез мышечного белка и снижается распад.
В процессе превращений в мышцах образуются аланин и глутамин, их условно можно считать транспортными формами азота. Аланин непосредственно из мышц попадает в печень, а глутамин вначале поступает в кишечник, где частично превращается в аланин. Поскольку в печени из аланина происходит синтез глюкозы, частично обеспечивающий мышцу энергией, получающийся кругооборот получил название глюкозо-аланинового цикла.
К азотсодержащим веществам мышц также относятся высокоэнергетичный креатин-фосфат и продукт его деградации креатинин. Экскреция креатинина обычно расматривается как мера мышечной массы. Однако это соединение может поступать в организм с высокобелковой пищей и влиять на результаты исследования содержания его в моче. Продукт распада миофибриллярных белков – 3-метилгистидин экскретируется с мочой в течение короткого времени и является достаточно точным показателем скорости распада в мышцах – при мышечном истощении скорость его выхода пропорционально снижается.
Почки не только выводят конечные продукты азотного распада (мочевину, креатинин и др.), но и являются дополнительным местом ресинтеза глюкозы из аминокислот, а также регулируют образование аммиака, компенсируя избыток ионов водорода в крови. Глюконеогенез и функционирование кислотно-щелочной регуляции тесно скоординированы, поскольку субстраты этих процессов появляются при дезаминировании аминокислот: углерод для синтеза глюкозы и азот – для аммиака. Существует даже мнение, что именно производство глюкозы является основной реакцией почек на ацидоз, а образование аммиака происходит вторично.
Для нервной ткани характерны более высокие концентрации аминокислот, чем в плазме. Это позволяет обеспечить мозг достаточным количеством ароматических аминокислот, являющихся предшественниками нейромедиаторов. Некоторые заменимые аминокислоты, такие как глутамат (из которого при участии пиридоксина образуется ГАМ К) и аспартат, также обладают влиянием на возбудимость нервной ткани. Их концентрация здесь высока, при этом заменимые аминокислоты способны синтезироваться и на месте.
Специфическую роль играет триптофан, являющийся предшественником серотонина. Именно с повышением концентрации триптофана (а, следовательно, и серотонина) связана сонливость после еды. Такой эффект особенно выражен при приеме больших количеств триптофана совместно с углеводной пищей. Повышенная секреция инсулина снижает уровень в крови аминокислот с ветвящимися цепями, которые при преодолении барьера кровь – мозг обладают конкурентными взаимоотношениями с ароматическими, но в то же время не оказывает влияния на концентрацию связанного с альбумином триптофана. Благодаря подобным эффектам препараты триптофана могут использоваться в психиатрической практике.
Ограничение ароматических аминокислот в рационе, в связи с их влиянием на центральную нервную систему, имеет профилактическое значение при ведении пациентов с печеночной энцефалопатией. Элементные аминокислотные диеты с преимущественным содержанием лейцина, изолейцина, валина и аргинина помогают избежать развития белковой недостаточности у гепатологических больных, и в то же время не приводят к возникновению печеночной комы.
Основные пластические функции протеиногенных аминокислот перечислены в табл. 3.2.
Таблица 3.2
Основные функции аминокислот
Углеводы
Углеводы – это полиатомные альдегидо– или кетоспирты, которые подразделяются в зависимости от количества мономеров на моно-, олиго– и полисахариды.
Таблица 3.3
Основные представители углеводов

Моносахариды (глюкоза, фруктоза, галактоза и др.), олигосахариды (сахароза, мальтоза, лактоза) и перевариваемые полисахариды (крахмал, гликоген) являются основными источниками энергии, а также выполняют пластическую функцию (табл. 3.3).
Неперевариваемые полисахариды, или пищевые волокна, играют в питании важнейшую роль, участвуя в формировании каловых масс, регулируя моторную функцию кишечника, выступая в качестве сорбентов (табл. 3.4). Разделяют нерастворимые полисахариды: целлюлоза, гемицеллюлоза; растворимые: каррагинаны, альгинаты, арабиногалактан и др. Именно растворимые полисахариды являются пробиотиками. К пищевым волокнам относят и не являющийся углеводом лигнин.
Таблица 3.4
Роль неперевариваемых полисахаридов (пищевых волокон) в питании

Перевариваемые углеводы в тонком кишечнике расщепляются до дисахаридов, а далее – до моносахаридов (рис. 3.2).
Всасывание моносахаров происходит путем облегченной диффузии и активного транспорта, что обеспечивает высокую их абсорбцию даже при низкой концентрации в кишечнике. Основным углеводным мономером является глюкоза, которая по системе воротной вене доставляется в печень, а далее или метаболизируется в ней, или поступает в общий кровоток и доставляется в органы и ткани.
Метаболизм глюкозы в тканях начинается с образования глюкозо-6-фосфата, который, в отличие от свободной глюкозы, не способен покидать клетку. Дальнейшие превращения этого соединения идут в следующих направлениях:
– расщепление вновь до глюкозы в печени, почках и эпителии кишечника, что позволяет поддерживать постоянный уровень сахара в крови;
– синтез депонируемой формы глюкозы – гликогена – в печени, мышцах и почках;
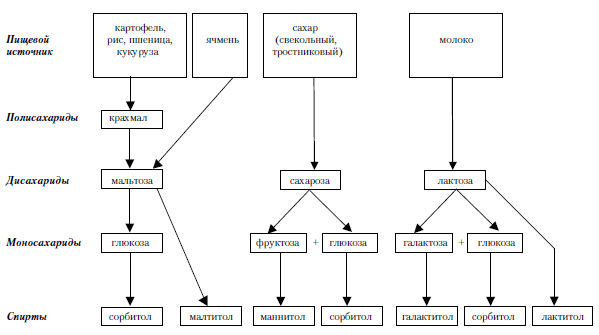
Рис. 3.2. Обмен пищевых растворимых углеводов (схема).
– окисление по основному (аэробному) пути катаболизма;
– окисление по пути гликолиза (анаэробного катаболизма), обеспечивающего энергией интенсивно работающие (мышечная ткань) или лишенные митохондрий (эритроциты) ткани и клетки;
– по пентозофосфатному пути превращений, происходящему под действием коферментной формы витамина в ходе которого генерируются продукты, используемые в синтезе биологически значимых молекул (НАДФ · Н2, нуклеиновых кислот).
Таким образом, метаболизм глюкозы может происходить по различным направлениям, использующим ее энергетический потенциал, пластические возможности или способность депонироваться.
Обеспечение тканей глюкозой, как энергетическим материалом, происходит за счет экзогенных сахаров, использования запасов гликогена и синтеза глюкозы из неуглеводных предшественников. В базальном (доабсорбционном) состоянии печень вырабатывает глюкозу со скоростью, равной ее утилизации во всем организме. Примерно 30 % производства глюкозы печенью происходит за счет гликогенолиза, а 70 % – как результат глюконеогенеза. Общее содержание гликогена в организме составляет примерно 500 г. Если нет экзогенного поступления глюкозы, его запасы истощаются через 12–18 часов. При отсутствии резервного гликогена в результате голодания резко усиливаются процессы окисления другого энергетического субстрата – жирных кислот. Одновременно увеличивается скорость глюконеогенеза, направленного в первую очередь на обеспечение глюкозой головного мозга, для которого она является основным источником энергии.
Синтез глюкозы происходит из аминокислот, лактата, пирувата, глицерина и жирных кислот с нечетной углеродной цепью. Большинство аминокислот способны быть предшественниками глюкозы, однако основную роль при этом, как сказано выше, играет аланин. Из аминокислотных источников происходит синтез примерно 6 % эндогенной глюкозы, из глицерина, пирувата и лактата соответственно 2 %, 1 % и 16 %. Вклад жирных кислот в глюконеогенез малозначим, поскольку лишь небольшой процент их имеет нечетное углеродное число.
В постабсорбционном состоянии печень из органа, вырабатывающего глюкозу, превращается в запасающий. При повышении концентрации глюкозы скорость ее утилизации периферическими тканями почти не изменяется, поэтому основным механизмом элиминации ее из кровотока является депонирование. Только небольшая часть избыточной глюкозы непосредственно участвует в липогенезе, который происходит в печени и в жировой ткани. Эти особенности углеводного метаболизма становятся значимыми при парентеральном введении высококонцентрированных растворов глюкозы.
Таблица 3.3
Основные представители углеводов
Моносахариды (глюкоза, фруктоза, галактоза и др.), олигосахариды (сахароза, мальтоза, лактоза) и перевариваемые полисахариды (крахмал, гликоген) являются основными источниками энергии, а также выполняют пластическую функцию (табл. 3.3).
Неперевариваемые полисахариды, или пищевые волокна, играют в питании важнейшую роль, участвуя в формировании каловых масс, регулируя моторную функцию кишечника, выступая в качестве сорбентов (табл. 3.4). Разделяют нерастворимые полисахариды: целлюлоза, гемицеллюлоза; растворимые: каррагинаны, альгинаты, арабиногалактан и др. Именно растворимые полисахариды являются пробиотиками. К пищевым волокнам относят и не являющийся углеводом лигнин.
Таблица 3.4
Роль неперевариваемых полисахаридов (пищевых волокон) в питании
Перевариваемые углеводы в тонком кишечнике расщепляются до дисахаридов, а далее – до моносахаридов (рис. 3.2).
Всасывание моносахаров происходит путем облегченной диффузии и активного транспорта, что обеспечивает высокую их абсорбцию даже при низкой концентрации в кишечнике. Основным углеводным мономером является глюкоза, которая по системе воротной вене доставляется в печень, а далее или метаболизируется в ней, или поступает в общий кровоток и доставляется в органы и ткани.
Метаболизм глюкозы в тканях начинается с образования глюкозо-6-фосфата, который, в отличие от свободной глюкозы, не способен покидать клетку. Дальнейшие превращения этого соединения идут в следующих направлениях:
– расщепление вновь до глюкозы в печени, почках и эпителии кишечника, что позволяет поддерживать постоянный уровень сахара в крови;
– синтез депонируемой формы глюкозы – гликогена – в печени, мышцах и почках;
Рис. 3.2. Обмен пищевых растворимых углеводов (схема).
– окисление по основному (аэробному) пути катаболизма;
– окисление по пути гликолиза (анаэробного катаболизма), обеспечивающего энергией интенсивно работающие (мышечная ткань) или лишенные митохондрий (эритроциты) ткани и клетки;
– по пентозофосфатному пути превращений, происходящему под действием коферментной формы витамина в ходе которого генерируются продукты, используемые в синтезе биологически значимых молекул (НАДФ · Н2, нуклеиновых кислот).
Таким образом, метаболизм глюкозы может происходить по различным направлениям, использующим ее энергетический потенциал, пластические возможности или способность депонироваться.
Обеспечение тканей глюкозой, как энергетическим материалом, происходит за счет экзогенных сахаров, использования запасов гликогена и синтеза глюкозы из неуглеводных предшественников. В базальном (доабсорбционном) состоянии печень вырабатывает глюкозу со скоростью, равной ее утилизации во всем организме. Примерно 30 % производства глюкозы печенью происходит за счет гликогенолиза, а 70 % – как результат глюконеогенеза. Общее содержание гликогена в организме составляет примерно 500 г. Если нет экзогенного поступления глюкозы, его запасы истощаются через 12–18 часов. При отсутствии резервного гликогена в результате голодания резко усиливаются процессы окисления другого энергетического субстрата – жирных кислот. Одновременно увеличивается скорость глюконеогенеза, направленного в первую очередь на обеспечение глюкозой головного мозга, для которого она является основным источником энергии.
Синтез глюкозы происходит из аминокислот, лактата, пирувата, глицерина и жирных кислот с нечетной углеродной цепью. Большинство аминокислот способны быть предшественниками глюкозы, однако основную роль при этом, как сказано выше, играет аланин. Из аминокислотных источников происходит синтез примерно 6 % эндогенной глюкозы, из глицерина, пирувата и лактата соответственно 2 %, 1 % и 16 %. Вклад жирных кислот в глюконеогенез малозначим, поскольку лишь небольшой процент их имеет нечетное углеродное число.
В постабсорбционном состоянии печень из органа, вырабатывающего глюкозу, превращается в запасающий. При повышении концентрации глюкозы скорость ее утилизации периферическими тканями почти не изменяется, поэтому основным механизмом элиминации ее из кровотока является депонирование. Только небольшая часть избыточной глюкозы непосредственно участвует в липогенезе, который происходит в печени и в жировой ткани. Эти особенности углеводного метаболизма становятся значимыми при парентеральном введении высококонцентрированных растворов глюкозы.