Страница:
состав сообщества и биоценоза по видам и численности особей в них определяется тем фактором среды, который находится в своем минимуме (наиболее неблагоприятен) для данного сообщества или биоценоза.В 1939 г. он сформулировал известные биоценотические принципы:
принцип (правило) разнообразия условий биотопа:
чем разнообразнее условия жизни в рамках биотопа, | тем больше число видов в заселяющем его биоценозе;принцип отклонения условий существования от нормы:
чем больше отклонение условий существования от оптимума (нормы) в пределах биотопа, тем беднее видами становится заселяющий его биоценоз и тем относительно больше особей имеет каждый присутствующий вид.Число особей внутри вида и число видов в биоценозе обратно пропорциональны. В этом можно легко убедиться, сравнивая биоценозы тундры и тропических лесов. Кроме того, в результате многочисленных исследований была определена биомасса ряда водных биоценозов, выраженная в килограмме сухой массы на 1 га морского дна:
Средиземное море…………….100
Балтийское море……………..330
Пролив Ла-Манш…………….400
Баренцево море ……………. 1 000
Берингово море…………….1 650
Северное море…………….. 3 460
Антарктида……………… 13 470
Из приведенных данных следует, что чем холоднее море, тем богаче жизнь.
В неблагоприятных условиях или условиях экологической монотонности число видов мало и межвидовая конкуренция снижается. Р. Крогерус (1932) констатировал эмпирическое правило приспособления (специализации) к экстремальным условиям:
в биотопах с экстремальными условиями складываются биоценозы из строго специализированных видов с относительно большим числом особей.Наиболее общей закономерностью распространения сообществ признано правило А. Уоллеса (1859):
видовое разнообразие увеличивается по мере продвижения с севера на юг.Причины этого многообразны, в частности, северные биоценозы исторически моложе и находятся в условиях меньшего поступления энергии от Солнца. Очевидно, что имеется в виду Северное полушарие Земли, а в Южном наблюдается обратная картина.
5.3. Пространственная структура биоценозов
Популяции различных видов биоценоза, подчиняясь соответствующим природным закономерностям, располагаются в пределах пространственных границ биотопа как по площади, так и по высоте.
Пространственная структура наземного биоценоза определяется закономерностью распределения надземных и подземных органов растительности по ярусам (расчленением растительных сообществ по высоте). Ярусное строение растительности (фитоценоза) позволяет максимально использовать лучистую энергию Солнца и зависит от теневыносливости растений. Ярусность хорошо выражена в лесах умеренного пояса. Так, например, в широколиственном лесу выделяются 5–6 ярусов: деревья первой, второй величины, подлесок, кустарник, высокие травы, низкие (приземные) травы. Существуют межъярусные растения – лишайники на стволах и ветках, лианы и др. Ярусность существует и в травянистых сообществах лугов, степей, саванн.
Ярусное строение подземных органов определяется разной глубиной проникновения корневых систем. В каждом ярусе растительности преимущественно обитают свои животные из состава биоценоза. Также существует разделение птиц на экологические группы по месту их питания (воздух, листва, ствол, земля). Дальнейшее подразделение этих групп в зависимости от основного типа пищи показано на рис. 5.14.
Несмотря на то что каждый вид стремится обзавестись собственной нишей, некоторая межвидовая конкуренция между ними за доступные ресурсы неизбежна.
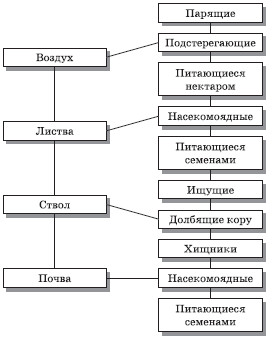 Рис. 5.14. Классификация сообществ птиц в Калифорнии с учетом пространственной структуры биоценоза
Рис. 5.14. Классификация сообществ птиц в Калифорнии с учетом пространственной структуры биоценоза
Ярусное строение наземных биоценозов тесно связано с их функциональной активностью. Так, пастбищные пищевые цепи преобладают в надземной части биоценозов, а детритные – в подземной. В водных экосистемах крупномасштабная вертикальная структура задается в первую очередь абиотическими условиями. Определяющими факторами являются градиенты освещенности, температуры, концентрации биогенов и т. п. На значительных глубинах усиливается влияние гидростатического давления. В донных биоценозах важны состав грунтов, гидродинамика придонных течений. Особенности вертикальной структуры выражаются в видовом составе, смене доминирующих видов, показателях биомассы и продуктивности. Фотосинтезирующие водоросли преобладают в верхних, хорошо освещенных горизонтах, что формирует вертикальные потоки вещества и энергии в направлении глубоководных биоценозов, жизнь которых основывается на привнесенной органике.
Пространственная структура биоценозов по горизонтали проявляется в их мозаичности и реализуется в виде неравномерного распределения популяций по площади из-за неоднородности почвенно-грунтовых условий, микроклимата, рельефа и т. п. Основой горизонтальной структуры могут служить особи одного вида, обладающего средообразующими свойствами, например, сосна со всеми связанными с ней микроорганизмами, грибами, лишайниками, насекомыми, птицами и т. д.
Пространственная структура наземного биоценоза определяется закономерностью распределения надземных и подземных органов растительности по ярусам (расчленением растительных сообществ по высоте). Ярусное строение растительности (фитоценоза) позволяет максимально использовать лучистую энергию Солнца и зависит от теневыносливости растений. Ярусность хорошо выражена в лесах умеренного пояса. Так, например, в широколиственном лесу выделяются 5–6 ярусов: деревья первой, второй величины, подлесок, кустарник, высокие травы, низкие (приземные) травы. Существуют межъярусные растения – лишайники на стволах и ветках, лианы и др. Ярусность существует и в травянистых сообществах лугов, степей, саванн.
Ярусное строение подземных органов определяется разной глубиной проникновения корневых систем. В каждом ярусе растительности преимущественно обитают свои животные из состава биоценоза. Также существует разделение птиц на экологические группы по месту их питания (воздух, листва, ствол, земля). Дальнейшее подразделение этих групп в зависимости от основного типа пищи показано на рис. 5.14.
Несмотря на то что каждый вид стремится обзавестись собственной нишей, некоторая межвидовая конкуренция между ними за доступные ресурсы неизбежна.
Ярусное строение наземных биоценозов тесно связано с их функциональной активностью. Так, пастбищные пищевые цепи преобладают в надземной части биоценозов, а детритные – в подземной. В водных экосистемах крупномасштабная вертикальная структура задается в первую очередь абиотическими условиями. Определяющими факторами являются градиенты освещенности, температуры, концентрации биогенов и т. п. На значительных глубинах усиливается влияние гидростатического давления. В донных биоценозах важны состав грунтов, гидродинамика придонных течений. Особенности вертикальной структуры выражаются в видовом составе, смене доминирующих видов, показателях биомассы и продуктивности. Фотосинтезирующие водоросли преобладают в верхних, хорошо освещенных горизонтах, что формирует вертикальные потоки вещества и энергии в направлении глубоководных биоценозов, жизнь которых основывается на привнесенной органике.
Пространственная структура биоценозов по горизонтали проявляется в их мозаичности и реализуется в виде неравномерного распределения популяций по площади из-за неоднородности почвенно-грунтовых условий, микроклимата, рельефа и т. п. Основой горизонтальной структуры могут служить особи одного вида, обладающего средообразующими свойствами, например, сосна со всеми связанными с ней микроорганизмами, грибами, лишайниками, насекомыми, птицами и т. д.
5.4. Экологические ниши видов в сообществах
Концепция ниши проникла во все разделы экологии, хотя первоначально она возникла как попытка дать всестороннее описание роли вида в сообществе, определяя все связи между популяциями, сообществом и всем биоценозом в данной среде обитания. В таком понимании ниша описывает, каким образом экологические объекты приспосабливаются друг к другу, образуя устойчиво функционирующее целое и позволяя понять, как даже весьма различные по своей организации сообщества могут быть похожи друг на друга.
Термин «ниша» введен Дж. Гриннеллом с целью обозначения самой мелкой единицы распространения вида. Уточненное формальное и потенциально количественное определение ниши дано Г. Хатчинсоном (1958), считавшим, что нишу следует определять с учетом всего диапазона физических, химических и биотических переменных среды, к которым должен быть адаптирован данный вид и под действием которых видовая популяция живет и возобновляется бесконечно долгое время. Теоретически такую переменную можно рассматривать как некий градиент, на котором у каждого вида имеется свой диапазон активности или устойчивости. Примером служит освещенность в лесу, которая убывает логарифмически от вершины деревьев к почве.
Далее Г. Хатчинсон подразделил видовую нишу на:
• фундаментальную – охватывает все множество оптимальных условий, в которых данный вид может обитать в отсутствие врагов;
• реализованную – фактический комплекс условий, в которых вид обычно существует.
Большинство организмов не обитает в своей потенциальной фундаментальной нише, а вследствие взаимодействий с другими организмами занимает меньшую по размерам реализованную нишу. Главными взаимодействиями обычно считают хищничество и конкуренцию. Последняя связана с теорией ниши через концепцию перекрывания ниш. Дело в том, что виды склонны частично разделять между собой свои фундаментальные ниши и в результате на один и тот же ресурс одновременно претендуют две или более популяции. По терминологии Г. Хатчинсона, «гиперпространства» ниш одних видов включают в себя также части «гиперпространств» других видов, т. е. перекрываются с ними.
Если перекрывание незначительно или ресурсы сверхобильны, то виды с перекрывающимися нишами могут сосуществовать в практически раздельных и почти фундаментальных нишах. В перекрывающихся частях любых двух ниш может произойти конкурентное исключение.
Итак, чем обильнее ресурс, тем менее вероятно, что его совместное использование приведет к конкуренции. Не следует ожидать конкуренции из-за неограниченного ресурса, например из-за наличия кислорода в большинстве наземных мест обитания.
Анализ перекрывания ниш привел к другому аспекту теории ниши – диффузной конкуренции. Обычно ниша данного вида перекрывается только с ограниченным количеством соседних ниш вдоль градиента одного ресурса, однако по мере изучения все большего числа факторов среды перечень потенциальных соседей возрастает. Поэтому, хотя попарное перекрывание ниш может быть невелико, суммарный эффект такой диффузной конкуренции может сильно сократить величину реализованной ниши – иногда даже до столь малых размеров, что она становится непригодна для поддержания жизнеспособной популяции (рис. 5.15). В результате вид может быть «выдавлен» группой других видов.
П. Джиллет (1984) сделал вывод, что сообщество представляет собой обширное я-мерное гиперпространство, в пределах которого каждая видовая популяция эволюционирует в таком направлении, чтобы соответствовать своей собственной части этого пространства. Ниша данного вида определяется его положением и реакцией на факторы гиперпространства данного сообщества. В результате каждый вид занимает неясно очерченное диффузное пространство, которое отличается от пространств, занимаемых другими видами сообщества, но может перекрываться с ними. Размеры ниш и их местоположение изменяются как в экологическом, так и в эволюционном (см. разд. 7.4) масштабах времени.
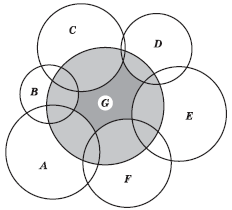 Рис. 5.15. Схема сокращения фундаментальной ниши вида G (темно-серая и светло-серые области) до его реализованной ниши (темно-серая область) в результате диффузной конкуренции с видами A, B, С, D, E и F (по П. Джиллету)
Рис. 5.15. Схема сокращения фундаментальной ниши вида G (темно-серая и светло-серые области) до его реализованной ниши (темно-серая область) в результате диффузной конкуренции с видами A, B, С, D, E и F (по П. Джиллету)
Термин «ниша» введен Дж. Гриннеллом с целью обозначения самой мелкой единицы распространения вида. Уточненное формальное и потенциально количественное определение ниши дано Г. Хатчинсоном (1958), считавшим, что нишу следует определять с учетом всего диапазона физических, химических и биотических переменных среды, к которым должен быть адаптирован данный вид и под действием которых видовая популяция живет и возобновляется бесконечно долгое время. Теоретически такую переменную можно рассматривать как некий градиент, на котором у каждого вида имеется свой диапазон активности или устойчивости. Примером служит освещенность в лесу, которая убывает логарифмически от вершины деревьев к почве.
Далее Г. Хатчинсон подразделил видовую нишу на:
• фундаментальную – охватывает все множество оптимальных условий, в которых данный вид может обитать в отсутствие врагов;
• реализованную – фактический комплекс условий, в которых вид обычно существует.
Большинство организмов не обитает в своей потенциальной фундаментальной нише, а вследствие взаимодействий с другими организмами занимает меньшую по размерам реализованную нишу. Главными взаимодействиями обычно считают хищничество и конкуренцию. Последняя связана с теорией ниши через концепцию перекрывания ниш. Дело в том, что виды склонны частично разделять между собой свои фундаментальные ниши и в результате на один и тот же ресурс одновременно претендуют две или более популяции. По терминологии Г. Хатчинсона, «гиперпространства» ниш одних видов включают в себя также части «гиперпространств» других видов, т. е. перекрываются с ними.
Если перекрывание незначительно или ресурсы сверхобильны, то виды с перекрывающимися нишами могут сосуществовать в практически раздельных и почти фундаментальных нишах. В перекрывающихся частях любых двух ниш может произойти конкурентное исключение.
Итак, чем обильнее ресурс, тем менее вероятно, что его совместное использование приведет к конкуренции. Не следует ожидать конкуренции из-за неограниченного ресурса, например из-за наличия кислорода в большинстве наземных мест обитания.
Анализ перекрывания ниш привел к другому аспекту теории ниши – диффузной конкуренции. Обычно ниша данного вида перекрывается только с ограниченным количеством соседних ниш вдоль градиента одного ресурса, однако по мере изучения все большего числа факторов среды перечень потенциальных соседей возрастает. Поэтому, хотя попарное перекрывание ниш может быть невелико, суммарный эффект такой диффузной конкуренции может сильно сократить величину реализованной ниши – иногда даже до столь малых размеров, что она становится непригодна для поддержания жизнеспособной популяции (рис. 5.15). В результате вид может быть «выдавлен» группой других видов.
П. Джиллет (1984) сделал вывод, что сообщество представляет собой обширное я-мерное гиперпространство, в пределах которого каждая видовая популяция эволюционирует в таком направлении, чтобы соответствовать своей собственной части этого пространства. Ниша данного вида определяется его положением и реакцией на факторы гиперпространства данного сообщества. В результате каждый вид занимает неясно очерченное диффузное пространство, которое отличается от пространств, занимаемых другими видами сообщества, но может перекрываться с ними. Размеры ниш и их местоположение изменяются как в экологическом, так и в эволюционном (см. разд. 7.4) масштабах времени.
5.5. Закономерности саморегуляции биоценозов, экологическое дублирование
Известный русский лесовед Г.Ф.Морозов (1912), опираясь на учение В. В. Докучаева и работы К. Мебиуса, сформулировал правило:
Если же измельчание особей ведет к производству относительно меньшего количества биомассы, то ее удельный выход с единицы площади (из-за более полного заселения пространства) увеличивается. Данный эмпирический факт Н. Ф. Реймерс назвал законом удельной продуктивности:
Несколько позже правила смены видов в биоценозах Н. Ф. Реймерс сформулировал в более популярной форме:
• «свято место пусто не бывает»;
• крупные организмы исчезают раньше, и их сменяют мелкие;
• как правило, более эволюционно высокоорганизованные виды бывают вытеснены менее высокоорганизованными, быстрее размножающимися существами;
• всегда побеждают те, кто легче и быстрее изменяется, в том числе генетически.
Проанализировав описанные теоретические основы закономерностей дублирования в биоценозах, Н. Ф. Реймерс (1973) выдвинул гипотетическую теорию механизма возникновения новых инфекционных заболеваний человека путем замены видов на неблагоприятные, опубликованную из-за существовавших ограничений только в 1983 г.
Он предположил, что в одних случаях возникает совершенно новая экологическая ниша, а в других случаях борьба с существующими заболеваниями и уничтожение их возбудителей освобождает такую нишу в человеческих популяциях.
Так, за 13 лет до открытия вируса иммунодефицита человека (ВИЧ) – возбудителя синдрома приобретенного иммунного дефицита (СПИД) – была предсказана вероятность появления «гриппоподобного заболевания с высокой летальностью». Организм человека – это место жизни многих, в том числе болезнетворных организмов. Когда многие болезни были побеждены и уничтожены их возбудители, появилась свободная экологическая ниша. Ее и заполнил вирус СПИДа, соответствующий приведенным свойствам: мелок, примитивен, относительно быстро размножается и очень изменчив.
Установлено, что в биоценозах существует столько видов, сколько необходимо для максимальной утилизации приходящей энергии и обеспечения круговорота веществ в пределах энергетического потока. В связи с этим к правилу Уоллеса добавляется принцип эколого-географического максимума (стабильности числа) видов:
Балансовый подход был уточнен Г. Реммертом (1978), сформулировавшим принцип продукционной оптимизации:
Именно как результат экологического дублирования, сдвига в подвижном равновесии и снижения «рентабельности» биоценоза возникают явления массового размножения нежелательных для хозяйства организмов. Монокультура в сельском хозяйстве, однопородные и одновозрастные леса «с позиций природы» чрезвычайно мало рентабельны, неравновесны и потому «требуют исправления» массовыми организмами.
Для хозяйственной деятельности особенно велико значение правила монокультуры Ю. Одума:
в природе не существует полезных и вредных птиц, полезных и вредных насекомых, там все служит друг другу и взаимоприспособлено.Современная формулировка правила Мебиуса—Морозова – правила взаимоприспособленности организмов в биоценозе – гласит:
виды в биоценозе приспособлены друг к другу настолько, что их сообщество составляет внутреннее противоречивое, но единое и взаимно увязанное системное целое.Ю. Одум установил правило «метаболизм и размеры особей», которое в идеальных условиях[30] имеет всеобщее значение:
при неизменном энергетическом потоке в пищевой сети или цепи более мелкие наземные организмы с высоким удельным метаболизмом создают относительно меньшую биомассу, чем крупные. Значительная часть энергии уходит на поддержание обмена веществ.Правило Ю. Одума особенно важно в связи с тем, что из-за антропогенного нарушения природной среды происходит измельчение «средней» особи живого на суше – крупные звери и птицы истреблены и вообще все крупные представители растительного и животного мира становятся все более и более раритетными. Поэтому неминуемо следует ожидать общее снижение относительной продуктивности организмов суши и термодинамический разлад в сообществах и биоценозах.
Если же измельчание особей ведет к производству относительно меньшего количества биомассы, то ее удельный выход с единицы площади (из-за более полного заселения пространства) увеличивается. Данный эмпирический факт Н. Ф. Реймерс назвал законом удельной продуктивности:
никогда слоны не дадут той биомассы и продукции с единицы площади, которую способна дать саранча и тем более совсем мелкие беспозвоночные.Исчезновение видов, представленных крупными особями, меняет вещественно-энергетическую структуру биоценозов. Так как энергетический поток, проходящий через биоценоз в целом, практически не меняется (иначе бы произошла смена типа биоценоза), включаются механизмы биоценотического или экологического дублирования: организмы одной трофической группы или уровня экологической пирамиды закономерно замещают друг друга. Правило (принцип) экологического дублирования Н. Ф. Реймерс сформулировал следующим образом:
исчезающий или уничтожаемый вид живого в рамках одного уровня экологической пирамиды заменяет другой функционально-биоценотически аналогичный. Замена происходит по схеме: мелкий сменяет крупного, эволюционно нижеорганизованный – более высокоорганизованного, более генетически лабильный[31] и мутабельный[32] – менее генетически изменчивого.Поскольку экологическая ниша в биоценозе не может пустовать, экологическое дублирование происходит обязательно. Действительно, копытных в степи сменяют грызуны, а в ряде случаев растительноядные насекомые. При отсутствии хищников на водоразделах южного Сахалина в зарослях бамбука их роль выполняет серая крыса.
Несколько позже правила смены видов в биоценозах Н. Ф. Реймерс сформулировал в более популярной форме:
• «свято место пусто не бывает»;
• крупные организмы исчезают раньше, и их сменяют мелкие;
• как правило, более эволюционно высокоорганизованные виды бывают вытеснены менее высокоорганизованными, быстрее размножающимися существами;
• всегда побеждают те, кто легче и быстрее изменяется, в том числе генетически.
Проанализировав описанные теоретические основы закономерностей дублирования в биоценозах, Н. Ф. Реймерс (1973) выдвинул гипотетическую теорию механизма возникновения новых инфекционных заболеваний человека путем замены видов на неблагоприятные, опубликованную из-за существовавших ограничений только в 1983 г.
Он предположил, что в одних случаях возникает совершенно новая экологическая ниша, а в других случаях борьба с существующими заболеваниями и уничтожение их возбудителей освобождает такую нишу в человеческих популяциях.
Так, за 13 лет до открытия вируса иммунодефицита человека (ВИЧ) – возбудителя синдрома приобретенного иммунного дефицита (СПИД) – была предсказана вероятность появления «гриппоподобного заболевания с высокой летальностью». Организм человека – это место жизни многих, в том числе болезнетворных организмов. Когда многие болезни были побеждены и уничтожены их возбудители, появилась свободная экологическая ниша. Ее и заполнил вирус СПИДа, соответствующий приведенным свойствам: мелок, примитивен, относительно быстро размножается и очень изменчив.
Установлено, что в биоценозах существует столько видов, сколько необходимо для максимальной утилизации приходящей энергии и обеспечения круговорота веществ в пределах энергетического потока. В связи с этим к правилу Уоллеса добавляется принцип эколого-географического максимума (стабильности числа) видов:
число видов в составе географических зон и их биоценозов – относительно постоянно и регулируется вещественно-энергетическими процессами; это число всегда стремится к необходимому и достаточному максимуму.Дублирование – один из природных механизмов поддержания надежности биоценозов. Это наиболее мобильный способ их адаптации. При этом возможны и генетические изменения в популяциях типа усиления хищнических наклонностей у крысы в приведенном ранее для Сахалина примере. Также возможно межвидовое и внутривидовое дублирование, а в сельском хозяйстве даже межсортовое. Общий «смысл» дублирования остается тем же:
максимально полно провести и использовать поток энергии, стабилизировать биоценоз в меняющихся условиях существования.Это свойство было подмечено А. А. Еленкиным (1921), определившим принцип подвижного равновесия:
биотическое сообщество сохраняется как единое целое вопреки регулярным колебаниям среды его существования, но при воздействии необычных факторов структурно изменяется с переносом «точки опоры» на другие растительные компоненты (группы растений).Если необычные, нерегулярные факторы оказывают многолетнее воздействие, то сообщество формирует иную структуру. Однако, как правило, в биоценозе сохраняются элементы дублирования в виде очень большого числа видов, которые могут быть мобилизованы в случае новых резких изменений среды.
Балансовый подход был уточнен Г. Реммертом (1978), сформулировавшим принцип продукционной оптимизации:
отношение между первичной и вторичной продукцией (между продуцентами и консументами) соответствует принципу оптимизации – «рентабельности» биопродукции.Как правило, растения и другие продуценты дают биомассу достаточную, но не излишнюю для потребления всем биотическим сообществом (с эволюционно определенным запасом, который обеспечивает надежность системы и обычно в 100 раз превышает потребление в биоценозах суши). При относительном «перепроизводстве» органического вещества биоценоз становится «нерентабельным» и возникают предпосылки для массового размножения отдельных видов. После периода автоколебаний соотношение «популяция—потребление» уравновешивается, биоценоз стабилизируется, балансируются отношения между трофическими уровнями.
Именно как результат экологического дублирования, сдвига в подвижном равновесии и снижения «рентабельности» биоценоза возникают явления массового размножения нежелательных для хозяйства организмов. Монокультура в сельском хозяйстве, однопородные и одновозрастные леса «с позиций природы» чрезвычайно мало рентабельны, неравновесны и потому «требуют исправления» массовыми организмами.
Для хозяйственной деятельности особенно велико значение правила монокультуры Ю. Одума:
эксплуатируемые для нужд человека экосистемы (биоценозы), представленные одним видом, равно как и системы монокультур (например, сельскохозяйственные монокультуры), неустойчивы по своей природе.Пагубность монокультур «учтена» природой. Более того, ею выработана стратегия сохранения гомеостаза на основе поддержания разнообразия и взаимозависимости членов биоценоза. Один из путей реализации такой стратегии отражает принцип П. Эрлиха и П. Равена или теории сопряженной эволюции, называемой также принципом коэволюции:
случайное функциональное изменение жертв (потребляемого растения) ведет к закономерному изменению свойств хищников (потребителей), что в свою очередь стимулирует разнообразие как первых, так и вторых.Все перечисленные закономерности саморегуляции биоценозов обобщены Н. Ф. Реймерсом в виде принципа стабильности:
любая относительно замкнутая биосистема с проходящим через нее потоком энергии в ходе саморегуляции развивается в сторону устойчивого состояния.Им же сформулировано и обобщающее правило биоценотической надежности:
надежность биоценоза зависит от его энергетической эффективности в данных условиях среды и возможностей структурно-функциональной перестройки в ответ на изменение внешних воздействий (материала для дублирования, межвидового и внутривидового, поддержания продукционной «рентабельности» и т. п.).
5.6. Биоразнообразие
Многообразие биологических видов соответствует разнообразию условий обитания на Земле. Одна из главных особенностей живого состоит в заселении не только зон наиболее благоприятного климата, но и всех уголков планеты: высокогорных, глубоководных, подземных ареалов.
Многочисленность и разнообразие обитателей планеты соответствует разнообразию экологических ниш в биогеоценозах. Миллионы биологических видов – основной ресурс и базис устойчивости (гомеостаза) биосферы. Техногенные воздействия на природные экосистемы приводят к вымиранию многих видов, этот процесс катастрофически ускорился в XX в., он ведет к потере устойчивости отдельных экосистем и биосферы в целом.
Исследуя закономерности, определяющие разнообразие видов в живой природе и устойчивость биоценозов, Н. Ф. Реймерс предложил очень наглядную аналоговую модель, имеющую форму волчка (рис. 5.16). Диаметр колес (цилиндров) волчка пропорционален числу видов, а толщина (высота цилиндров) – биомассе соответствующего трофического уровня. При этом автором взято минимальное называемое число видов консументов, хотя в литературе оно приводится многократно большим (не менее 5 млн видов), что делает модель только нагляднее. Волчок вращает энергия Солнца, что на рис. 5.16 показано стрелками сверху. Модель справедлива для любого конкретного биоценоза.
Из анализа модели следует, что система становится устойчивее с наращиванием подводимой к ней мощности (количества энергии в единицу времени) и с увеличением диаметра среднего колеса волчка, т. е. количества консументов. Таким образом, консументы служат управляющим (балансирующим) звеном в системе биоценоза. Именно они порождают спектр разнообразия в биоценозе (сообществе) и препятствуют монополии доминантов. Массовое размножение «вредителей» на полях монокультуры и в моновидных омоложенных лесных насаждениях иллюстрирует сделанный вывод.
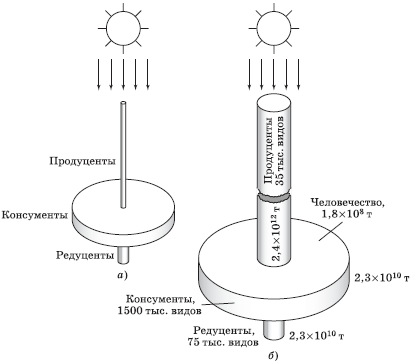 Рис. 5.16. «Волчок жизни» (по Н. Реймерсу): а – для упрощенной модельной экосистемы Ю. Одума: люцерна – телятина – мальчик (с включением других консументов и редуцентов); б – для биоценоза Земли в целом
Рис. 5.16. «Волчок жизни» (по Н. Реймерсу): а – для упрощенной модельной экосистемы Ю. Одума: люцерна – телятина – мальчик (с включением других консументов и редуцентов); б – для биоценоза Земли в целом
Поскольку эволюционные переходы в природе занимают сравнительно небольшое время, считается, что предельная совокупность видов могла быть образована в эволюции жизни всего за 70 млн лет. Дальнейшее нарастание количества видов практически исключено из-за ограниченности числа экологических ниш и принципа конкурентного исключения. Так как за последние 500 млн лет число видов оставалось практически одинаковым, Н. Ф. Реймерс сформулировал правило константности видов в ходе стационарной эволюции биосферы:[33]
Наиболее естественным природным таксоном является вид – классификационная единица низшего ранга. Современная систематика насчитывает в живой природе пять высших таксонов, представители которых различаются по типу обменных процессов и роли в природе: это бактерии, простейшие, грибы, растения и животные. В каждой из этих крупных групп организмов можно найти более примитивных и более морфологически и физиологически сложных представителей, причем все они в высокой степени адаптированы к среде своего обитания.
Бактерии. Бактерии (от греч. bakterion – палочка) – микроорганизмы с этопрокариотным типом строения клетки. Их клетки не имеют отделенного от цитоплазмы ядра. Однако генетическая программа, как и у всех живых организмов, закодирована в виде последовательности нуклеотидов в ДНК и несет информацию о структуре белков. Бактериальные клетки не содержат таких органелл, как хлоропласты (специализированных для фотосинтеза) и митохондрии (специализированных для клеточного дыхания и синтеза АТФ). Эти биохимические процессы происходят у бактерий в цитоплазме.
Роли бактерий в природе очень разнообразны, что связано с различными источниками энергии, используемыми разными группами бактерий. Многие гетеротрофные аэробные бактерии являются редуцентами в экосистемах. В почве они участвуют в образовании плодородного слоя, преобразуя лесную подстилку и гниющие остатки животных в гумус. Бактерии почвы также разлагают органические соединения до минеральных веществ. Установлено, что до 90 % СО2 попадает в атмосферу за счет деятельности бактерий и грибов. Бактерии участвуют в биогеохимических циклах азота, серы, фосфора. Самоочищение воды в природных водоемах, а также очистка сточных вод производится аэробными и анаэробными гетеротофными бактериями.
Бактерии-симбионты населяют кишечник травоядных животных; бактериальная микрофлора кишечника человека участвует в процессах переваривания целлюлозы (растительной клетчатки). Эти бактерии также синтезируют некоторые витамины. Нитрифицирующие бактерии – симбионты бобовых растений – обогащают почву азотом.
Многочисленность и разнообразие обитателей планеты соответствует разнообразию экологических ниш в биогеоценозах. Миллионы биологических видов – основной ресурс и базис устойчивости (гомеостаза) биосферы. Техногенные воздействия на природные экосистемы приводят к вымиранию многих видов, этот процесс катастрофически ускорился в XX в., он ведет к потере устойчивости отдельных экосистем и биосферы в целом.
Исследуя закономерности, определяющие разнообразие видов в живой природе и устойчивость биоценозов, Н. Ф. Реймерс предложил очень наглядную аналоговую модель, имеющую форму волчка (рис. 5.16). Диаметр колес (цилиндров) волчка пропорционален числу видов, а толщина (высота цилиндров) – биомассе соответствующего трофического уровня. При этом автором взято минимальное называемое число видов консументов, хотя в литературе оно приводится многократно большим (не менее 5 млн видов), что делает модель только нагляднее. Волчок вращает энергия Солнца, что на рис. 5.16 показано стрелками сверху. Модель справедлива для любого конкретного биоценоза.
Из анализа модели следует, что система становится устойчивее с наращиванием подводимой к ней мощности (количества энергии в единицу времени) и с увеличением диаметра среднего колеса волчка, т. е. количества консументов. Таким образом, консументы служат управляющим (балансирующим) звеном в системе биоценоза. Именно они порождают спектр разнообразия в биоценозе (сообществе) и препятствуют монополии доминантов. Массовое размножение «вредителей» на полях монокультуры и в моновидных омоложенных лесных насаждениях иллюстрирует сделанный вывод.
Поскольку эволюционные переходы в природе занимают сравнительно небольшое время, считается, что предельная совокупность видов могла быть образована в эволюции жизни всего за 70 млн лет. Дальнейшее нарастание количества видов практически исключено из-за ограниченности числа экологических ниш и принципа конкурентного исключения. Так как за последние 500 млн лет число видов оставалось практически одинаковым, Н. Ф. Реймерс сформулировал правило константности видов в ходе стационарной эволюции биосферы:[33]
число нарождающихся видов в среднем равно числу вымерших и общее видовое разнообразие в биосфере есть константа.Биологическая наука – систематика – подразделяет все живое на таксоны – группы организмов, имеющие общие черты морфологической организации и физиологических процессов и в то же время достаточно обособленные в природе, т. е. не имеющие гибридов с представителями других таксонов.
Наиболее естественным природным таксоном является вид – классификационная единица низшего ранга. Современная систематика насчитывает в живой природе пять высших таксонов, представители которых различаются по типу обменных процессов и роли в природе: это бактерии, простейшие, грибы, растения и животные. В каждой из этих крупных групп организмов можно найти более примитивных и более морфологически и физиологически сложных представителей, причем все они в высокой степени адаптированы к среде своего обитания.
Бактерии. Бактерии (от греч. bakterion – палочка) – микроорганизмы с этопрокариотным типом строения клетки. Их клетки не имеют отделенного от цитоплазмы ядра. Однако генетическая программа, как и у всех живых организмов, закодирована в виде последовательности нуклеотидов в ДНК и несет информацию о структуре белков. Бактериальные клетки не содержат таких органелл, как хлоропласты (специализированных для фотосинтеза) и митохондрии (специализированных для клеточного дыхания и синтеза АТФ). Эти биохимические процессы происходят у бактерий в цитоплазме.
Роли бактерий в природе очень разнообразны, что связано с различными источниками энергии, используемыми разными группами бактерий. Многие гетеротрофные аэробные бактерии являются редуцентами в экосистемах. В почве они участвуют в образовании плодородного слоя, преобразуя лесную подстилку и гниющие остатки животных в гумус. Бактерии почвы также разлагают органические соединения до минеральных веществ. Установлено, что до 90 % СО2 попадает в атмосферу за счет деятельности бактерий и грибов. Бактерии участвуют в биогеохимических циклах азота, серы, фосфора. Самоочищение воды в природных водоемах, а также очистка сточных вод производится аэробными и анаэробными гетеротофными бактериями.
Бактерии-симбионты населяют кишечник травоядных животных; бактериальная микрофлора кишечника человека участвует в процессах переваривания целлюлозы (растительной клетчатки). Эти бактерии также синтезируют некоторые витамины. Нитрифицирующие бактерии – симбионты бобовых растений – обогащают почву азотом.