Страница:
3.5. Организмы – индикаторы качества среды
Чувствительность организмов к изменениям условий среды и особенно к наличию конкретных химических примесей положена в основу биологической индикации и биотестирования, которые используют наряду с методами оценки загрязнения природной среды с помощью приборов. Редкие, как правило, стенобионтные (требующие строго определенных условий существования) виды часто являются лучшими индикаторами (показателями) состояния среды. Их исчезновение служит доказательством неблагоприятных воздействий на среду обитания в конкретных местах.
Наибольшее распространение получил метод л и х е н о-индикации (от лат. lichen – лишайник), основанный на учете количества лишайников в городских насаждениях, районах крупных предприятий. Наличие лишайников на стволах деревьев взаимосвязано с химическим составом (загрязненностью) воздуха. В Англии для определения средней концентрации SO2 (мг/м3) в атмосферном воздухе используют эмпирическое уравнение:
CSO2 = 18,72 – 3,94Q – 0,15h – 2,38p,
где Q – освещенность на высоте 1,5 м, лк; h – средняя высота мха на стволе, м; p – степень покрытия древесной растительности лишайниками, %.
В лесных массивах удобными индикаторами качества служат жуки-короеды. Сильно ослабленные и отмирающие деревья заселяются короедами. Однако, если причиной их гибели послужили химические вещества, не характерные нормальному составу воздуха, короеды не получают широкого распространения. Отмирание насаждений при отсутствии заселения деревьев короедами – надежное доказательство антропогенного загрязнения воздуха.
Поскольку в настоящее время установлены допустимые нормативы присутствия в воде не на все возможные вещества и далеко не каждое из них можно определить непосредственно химическим экспериментом, то биотестирование можно считать обязательным для получения интегральных оценок загрязненности водоемов к с е н о б и о т и к а м и (любое чужеродное для данного организма или группы организмов разного вида вещество, вызывающее нарушение биотических процессов, включая гибель организмов). В ряде случаев, и особенно в условиях аварийной ситуации, бывает необходим экспресс-анализ самого факта загрязнения воды. Хотя биотестирование и не позволяет установить спектр чужеродных веществ в воде (если они неизвестны из других источников), но общий вывод можно получить быстро и надежно.
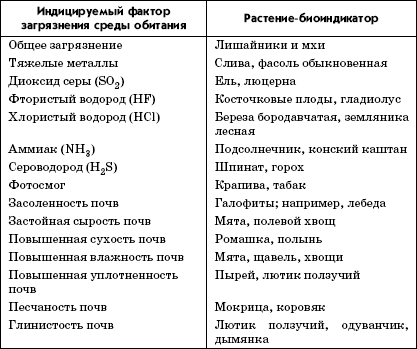
В качестве организмов-индикаторов (биоиндикаторов) используют бактерии, водоросли, беспозвоночные (инфузории, ракообразные, моллюски). По дикорастущим растениям (табл. 3.3) можно судить о характере и состоянии почвы, ибо среда обитания растений определяется такими свойствами почв, как влагоемкость, структура, плотность, температура, содержание кислорода, питательных веществ, тяжелых металлов и солей.
Таблица 3.3
Примеры растений-биоиндикаторов
 Показателем условий существования служит и общее число видов, обитающих в данном месте. Его уменьшение может указывать на изменение (загрязнение) раньше, чем выявится снижение общего количества особей или плодородия.
Показателем условий существования служит и общее число видов, обитающих в данном месте. Его уменьшение может указывать на изменение (загрязнение) раньше, чем выявится снижение общего количества особей или плодородия.
Наибольшее распространение получил метод л и х е н о-индикации (от лат. lichen – лишайник), основанный на учете количества лишайников в городских насаждениях, районах крупных предприятий. Наличие лишайников на стволах деревьев взаимосвязано с химическим составом (загрязненностью) воздуха. В Англии для определения средней концентрации SO2 (мг/м3) в атмосферном воздухе используют эмпирическое уравнение:
CSO2 = 18,72 – 3,94Q – 0,15h – 2,38p,
где Q – освещенность на высоте 1,5 м, лк; h – средняя высота мха на стволе, м; p – степень покрытия древесной растительности лишайниками, %.
В лесных массивах удобными индикаторами качества служат жуки-короеды. Сильно ослабленные и отмирающие деревья заселяются короедами. Однако, если причиной их гибели послужили химические вещества, не характерные нормальному составу воздуха, короеды не получают широкого распространения. Отмирание насаждений при отсутствии заселения деревьев короедами – надежное доказательство антропогенного загрязнения воздуха.
Поскольку в настоящее время установлены допустимые нормативы присутствия в воде не на все возможные вещества и далеко не каждое из них можно определить непосредственно химическим экспериментом, то биотестирование можно считать обязательным для получения интегральных оценок загрязненности водоемов к с е н о б и о т и к а м и (любое чужеродное для данного организма или группы организмов разного вида вещество, вызывающее нарушение биотических процессов, включая гибель организмов). В ряде случаев, и особенно в условиях аварийной ситуации, бывает необходим экспресс-анализ самого факта загрязнения воды. Хотя биотестирование и не позволяет установить спектр чужеродных веществ в воде (если они неизвестны из других источников), но общий вывод можно получить быстро и надежно.
В качестве организмов-индикаторов (биоиндикаторов) используют бактерии, водоросли, беспозвоночные (инфузории, ракообразные, моллюски). По дикорастущим растениям (табл. 3.3) можно судить о характере и состоянии почвы, ибо среда обитания растений определяется такими свойствами почв, как влагоемкость, структура, плотность, температура, содержание кислорода, питательных веществ, тяжелых металлов и солей.
Таблица 3.3
Примеры растений-биоиндикаторов
Контрольные вопросы и задания
3.1. Что такое среда обитания и какие среды заселены организмами?
3.2. Какие факторы среды относят к абиотическим и биотическим?
3.3. Как называют совокупность влияний жизнедеятельности одних организмов на жизнедеятельность других?
3.4. Что такое ресурсы живых существ, как они классифицируются и в чем их экологическое значение?
3.5. Как формулируется закон минимума? Какие существуют к нему уточнения?
3.6. Сформулируйте закон толерантности. Кто установил эту закономерность?
3.7. Приведите примеры использования законов минимума и толерантности в практической деятельности.
3.8. Какие механизмы позволят живым организмам компенсировать действие экологических факторов?
3.9. В чем различие между местообитанием и экологической нишей?
3.10. Какие факторы следует учитывать в первую очередь при создании проектов управления экосистемами. Почему?
3.2. Какие факторы среды относят к абиотическим и биотическим?
3.3. Как называют совокупность влияний жизнедеятельности одних организмов на жизнедеятельность других?
3.4. Что такое ресурсы живых существ, как они классифицируются и в чем их экологическое значение?
3.5. Как формулируется закон минимума? Какие существуют к нему уточнения?
3.6. Сформулируйте закон толерантности. Кто установил эту закономерность?
3.7. Приведите примеры использования законов минимума и толерантности в практической деятельности.
3.8. Какие механизмы позволят живым организмам компенсировать действие экологических факторов?
3.9. В чем различие между местообитанием и экологической нишей?
3.10. Какие факторы следует учитывать в первую очередь при создании проектов управления экосистемами. Почему?
ГЛАВА 4
ЭКОЛОГИЯ ПОПУЛЯЦИЙ[20] (ДЕМЭКОЛОГИЯ)
С. С. Четвериковым (1903) сформулировано правило объединения в популяции:
Популяция – это минимальная самовоспроизводящаяся группа особей одного вида, на протяжении эволюционно длительного времени населяющая определенное пространство, образующая генетическую систему и формирующая собственную экологическую нишу.[22]
У растений совокупность индивидуумов одного вида среди особей других видов называют ценопопуляцией.
Наиболее близким по значению к популяции является известное из курса истории понятие «племя».
Постоянное обновление популяции сопровождается сложными процессами обмена генетическим материалом, происходящим в результате случайного подбора родительских пар. В некотором смысле она напоминает котел, в котором происходит постоянное перемешивание материала, идущего на создание свойств новых, нарождающихся организмов. Одновременно происходит отбор и закрепление таких видовых признаков и свойств, которые оказываются полезными для поддержания жизни популяции в сложившихся условиях.
Необходимость сужения понятия «вид» до понятия «популяция» в биологии (в том числе и в экологии) вызвано следующим. Существование ареала распространения вида не означает реальной возможности всем особям свободно перемещаться в его границах. Степень подвижности особей выражается расстоянием, на которое может перемещаться животное, т. е. радиусом активности. Для растений этот радиус определяется расстоянием, на которое распространяется пыльца, семена или вегетативные части, способные дать начало новому растению. Для улитки он составляет несколько десятков метров, для ондатры – несколько сотен метров, для северного оленя – более ста километров.
Вследствие ограниченности радиуса активности лесные полевки, обитающие в одном лесу, имеют мало шансов встретиться в период размножения с полевками соседнего леса, хотя их встречи не исключаются полностью. Это приводит к обособлению отдельных групп – популяций. Так, например, популяцию образуют все особи окуня в небольшом озере или все деревья одного вида в лесу.
Таким образом, популяция представляет собой форму существования вида, обеспечивающую приспособленность его к конкретным условиям среды. Все живые существа входят в популяции.
Численность популяций может резко меняться по сезонам и годам. Известно массовое размножение в некоторые годы леммингов (мелких грызунов), саранчи, болезнетворных микробов, божьих коровок. У видов животных и растений с большой продолжительностью жизни и малой плодовитостью численность популяции более устойчивая. У насекомых и мелких
растений на открытых пространствах она нередко составляет сотни тысяч и миллионы особей.
Популяция обладает не только биологическими свойствами составляющих ее организмов, но и собственными, присущими только группе особей в целом. Рождаемость, смертность, возрастная структура, плотность населения и другие свойства могут иметь смысл только на групповом уровне. Основными экологическими характеристиками популяции считаются:
• величина по занимаемому пространству и по численности особей;
• структура возрастная, половая, пространственная, экологическая и др.;
• динамика.
Имея дело с растениями, особенно культурными, целесообразно, а иногда и проще учитывать биомассу,[23] а не число особей.
Минимальный размер для самовоспроизводства на протяжении разных поколений подразумевает численность, достаточную для выживания популяции при резких ее изменениях.
Под численностью популяции понимают эффективную величину численности, т. е. число размножающихся животных, которое всегда меньше общего числа особей, составляющих популяцию.
Группы многочисленных видов одомашненных или селекционно выведенных домашних животных и растений можно считать популяциями лишь с серьезными оговорками, ибо они всецело зависят от человека. Тем не менее при одичании домашние животные в природных условиях могут образовывать популяции.
индивиды любого вида живого всегда представлены в природной среде не изолированными отдельностями, а только их определенным образом организованными совокупностями.Особи любого вида живых организмов распределены в пределах ареала неравномерно. Наблюдения показывают, что существует «островная» форма распределения групп особей и форма «сгущений». Участки с относительно высокой встречаемостью особей данного вида чередуются с участками с низкой плотностью. «Центры плотности» населения каждого вида и являются, как правило, популяциями. Популяция – первая надорганизменная структурная единица вида.[21]
Популяция – это минимальная самовоспроизводящаяся группа особей одного вида, на протяжении эволюционно длительного времени населяющая определенное пространство, образующая генетическую систему и формирующая собственную экологическую нишу.[22]
У растений совокупность индивидуумов одного вида среди особей других видов называют ценопопуляцией.
Наиболее близким по значению к популяции является известное из курса истории понятие «племя».
Постоянное обновление популяции сопровождается сложными процессами обмена генетическим материалом, происходящим в результате случайного подбора родительских пар. В некотором смысле она напоминает котел, в котором происходит постоянное перемешивание материала, идущего на создание свойств новых, нарождающихся организмов. Одновременно происходит отбор и закрепление таких видовых признаков и свойств, которые оказываются полезными для поддержания жизни популяции в сложившихся условиях.
Необходимость сужения понятия «вид» до понятия «популяция» в биологии (в том числе и в экологии) вызвано следующим. Существование ареала распространения вида не означает реальной возможности всем особям свободно перемещаться в его границах. Степень подвижности особей выражается расстоянием, на которое может перемещаться животное, т. е. радиусом активности. Для растений этот радиус определяется расстоянием, на которое распространяется пыльца, семена или вегетативные части, способные дать начало новому растению. Для улитки он составляет несколько десятков метров, для ондатры – несколько сотен метров, для северного оленя – более ста километров.
Вследствие ограниченности радиуса активности лесные полевки, обитающие в одном лесу, имеют мало шансов встретиться в период размножения с полевками соседнего леса, хотя их встречи не исключаются полностью. Это приводит к обособлению отдельных групп – популяций. Так, например, популяцию образуют все особи окуня в небольшом озере или все деревья одного вида в лесу.
Таким образом, популяция представляет собой форму существования вида, обеспечивающую приспособленность его к конкретным условиям среды. Все живые существа входят в популяции.
Численность популяций может резко меняться по сезонам и годам. Известно массовое размножение в некоторые годы леммингов (мелких грызунов), саранчи, болезнетворных микробов, божьих коровок. У видов животных и растений с большой продолжительностью жизни и малой плодовитостью численность популяции более устойчивая. У насекомых и мелких
растений на открытых пространствах она нередко составляет сотни тысяч и миллионы особей.
Популяция обладает не только биологическими свойствами составляющих ее организмов, но и собственными, присущими только группе особей в целом. Рождаемость, смертность, возрастная структура, плотность населения и другие свойства могут иметь смысл только на групповом уровне. Основными экологическими характеристиками популяции считаются:
• величина по занимаемому пространству и по численности особей;
• структура возрастная, половая, пространственная, экологическая и др.;
• динамика.
Имея дело с растениями, особенно культурными, целесообразно, а иногда и проще учитывать биомассу,[23] а не число особей.
Минимальный размер для самовоспроизводства на протяжении разных поколений подразумевает численность, достаточную для выживания популяции при резких ее изменениях.
Под численностью популяции понимают эффективную величину численности, т. е. число размножающихся животных, которое всегда меньше общего числа особей, составляющих популяцию.
Группы многочисленных видов одомашненных или селекционно выведенных домашних животных и растений можно считать популяциями лишь с серьезными оговорками, ибо они всецело зависят от человека. Тем не менее при одичании домашние животные в природных условиях могут образовывать популяции.
4.1. Размер популяции (популяционные законы)
Численность популяции не произвольна даже при постоянной средней продолжительности жизни, а изменяется в пределах определенного диапазона. В соответствии с правилом максимума размера колебаний плотности популяционного населения Ю. Одума
С другой стороны, размеры популяции уменьшаются в результате эмиграции и (или) смертности. Таким образом,
И3МЕНЕНИЕ ЧИСЛЕННОСТИ ПОПУЛЯЦИИ = (РОЖДАЕМОСТЬ + ИММИГРАЦИЯ) – (СМЕРТНОСТЬ + ЭМИГРАЦИЯ).
Для стабильных популяций справедливо утверждение, что в них «рождаемость уравновешена смертностью». Один из основных факторов, влияющих на размеры популяции, – это процент особей, погибающих до достижения половозрелого возраста.
существуют определенные верхние и нижние пределы для размеров плотности (численности) популяции, которые соблюдаются в природе или которые теоретически могли бы существовать в течение сколь угодно длительного отрезка времени в условиях стабильности среды обитания.К. Фридерихсом (1927) была сформулирована теория биоценотической регуляции численности популяции:
регулирование численности популяции есть результат комплекса воздействий абиотической и биотической среды в местообитании вида.Размеры популяции возрастают в результате иммиграции из соседних популяций и (или) за счет размножения особей. Общая численность и плотность населения популяций регулируется правилом максимальной рождаемости (воспроизводства):
в популяции имеется тенденция к образованию теоретически максимально возможного количества новых особей.Максимальную рождаемость достигают в идеальных условиях, когда отсутствуют лимитирующие экологические факторы, и размножение ограничено лишь физиологическими особенностями вида. Обычно существует экологическая или реализуемая рождаемость, возникающая в обычных или специфических условиях среды.
С другой стороны, размеры популяции уменьшаются в результате эмиграции и (или) смертности. Таким образом,
И3МЕНЕНИЕ ЧИСЛЕННОСТИ ПОПУЛЯЦИИ = (РОЖДАЕМОСТЬ + ИММИГРАЦИЯ) – (СМЕРТНОСТЬ + ЭМИГРАЦИЯ).
Для стабильных популяций справедливо утверждение, что в них «рождаемость уравновешена смертностью». Один из основных факторов, влияющих на размеры популяции, – это процент особей, погибающих до достижения половозрелого возраста.
4.2. Возрастная и половая структуры популяций
Средняя продолжительность жизни организмов и соотношение численности (или биомассы) особей различного возраста характеризуются возрастной, а соотношение особей разного пола – половой структурами популяции (рис. 4.1). Формирование возрастной структуры происходит в результате совместного действия процессов размножения и смертности.
Со временем даже в пределах одной и той же популяции могут происходить значительные изменения возрастной структуры. В таких случаях «включаются механизмы», автоматически возвращающие популяцию к некоторому нормальному для данной популяции возрастному распределению. Причем популяции, включающие в себя много возрастных групп, в меньшей степени подвержены воздействию факторов, влияющих на размножение в конкретном году.
Даже крайне неблагоприятные условия, вызывающие полную гибель приплода в тот или иной год, не становятся катастрофическими для популяции сложной структуры, так как родительские пары участвуют в воспроизводстве многократно. Тем не менее следы изменений условий жизни сохраняются в облике сложных популяций гораздо дольше, чем у простых.
Для роста численности популяций большое значение имеет соотношение особей по полу. Генетический механизм обеспечивает примерно равное соотношение особей разного пола при рождении. Однако исходное соотношение вскоре нарушается в результате различий физиологических, поведенческих и экологических реакций самцов и самок, вызывающих неравномерную смертность.
Анализ возрастной и половой структуры популяции позволяет прогнозировать ее численность на ряд ближайших поколений и лет. Этим пользуются для оценки возможности промысла рыбы, в охотничьих хозяйствах и прочих подобных случаях.
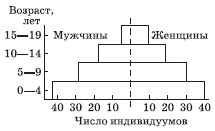 Рис. 4.1. Гистограмма распределения индивидуумов по возрастам в соответствии с полом для гипотетической популяции, имеющей высокую рождаемость и высокую смертность в ранних возрастных группах
Рис. 4.1. Гистограмма распределения индивидуумов по возрастам в соответствии с полом для гипотетической популяции, имеющей высокую рождаемость и высокую смертность в ранних возрастных группах
Со временем даже в пределах одной и той же популяции могут происходить значительные изменения возрастной структуры. В таких случаях «включаются механизмы», автоматически возвращающие популяцию к некоторому нормальному для данной популяции возрастному распределению. Причем популяции, включающие в себя много возрастных групп, в меньшей степени подвержены воздействию факторов, влияющих на размножение в конкретном году.
Даже крайне неблагоприятные условия, вызывающие полную гибель приплода в тот или иной год, не становятся катастрофическими для популяции сложной структуры, так как родительские пары участвуют в воспроизводстве многократно. Тем не менее следы изменений условий жизни сохраняются в облике сложных популяций гораздо дольше, чем у простых.
Для роста численности популяций большое значение имеет соотношение особей по полу. Генетический механизм обеспечивает примерно равное соотношение особей разного пола при рождении. Однако исходное соотношение вскоре нарушается в результате различий физиологических, поведенческих и экологических реакций самцов и самок, вызывающих неравномерную смертность.
Анализ возрастной и половой структуры популяции позволяет прогнозировать ее численность на ряд ближайших поколений и лет. Этим пользуются для оценки возможности промысла рыбы, в охотничьих хозяйствах и прочих подобных случаях.
4.3. Пространственная и этологическая структуры популяций
Пространственная структура. Каждая популяция занимает пространство, обеспечивающее условия жизни для ограниченного числа особей. При изучении пространственной структуры различают случайное, равномерное и неравномерное (групповое) распределения особей на территории (в пространстве).
Случайное распределение (рис. 4.2, а) в природе встречается редко; оно наблюдается в случаях, когда среда очень однородна, а организмы не стремятся объединиться в группы. Равномерное распределение (рис. 4.2, б) бывает там, где между особями очень сильна конкуренция или существует антагонизм. Наиболее часто наблюдается неравномерное (групповое) распределение (рис. 4.2, в) – образование различных скоплений.
Активность особей, пар и семейных групп у позвоночных и высших беспозвоночных обычно ограничена определенной зоной, называемой индивидуальным (или семейным) участком территории. У высших животных внутрипопуляционное распределение регулируется системой инстинктов. Им свойственно особое территориальное поведение – реакция на местонахождение других членов популяции.
В зависимости от характера использования пространства подвижных животных подразделяют на о с е д л ы х и к о ч е в ы х. Оседлые животные в течение всей или большей части жизни используют довольно ограниченный участок среды. Им присущи инстинкты привязанности к своему участку, регулярное возвращение к месту размножения после длительных и дальних миграций.
Кочевые животные совершают постоянные передвижения в пространстве, так как они зависят от запаса корма на данной территории. Кочевой образ жизни характерен преимуществен
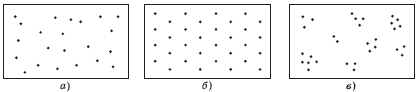 Рис. 4.2. Основные типы распределения особей популяции по территории по Ю. Одуму
Рис. 4.2. Основные типы распределения особей популяции по территории по Ю. Одуму
но для стад и стай (кочующие группы многих рыб во время нагульных миграций, стада слонов, зебр, антилоп, северных оленей и т. п.). Перемещение кочующих видов по площади обычно происходит за сроки, достаточные для восстановления кормовых ресурсов на пастбищных участках. Масштабы и длительность таких миграций определяют обилие пищи и численность стада.
Этологическая (поведенческая) структура. Она отражает разнообразные формы совместного существования особей в популяциях. О д и н о ч н ы й о б р а з жизни следует выделить в первую очередь, хотя полностью одиночного существования организмов в природе нет, так как в этом случае было бы невозможно размножение. С е м е й н ы й о б р а з жизни – усиливаются связи между родителями и потомством, начинает заметно проявляться территориальное поведение животных. Путем различных сигналов, маркировки, угроз и тому подобного обеспечивается владение участком, достаточным для выкармливания потомства.
Стая – временное объединение животных, проявляющих биологически полезную организованность действий (для защиты от врагов, добычи пищи, миграции и т. п.). Наиболее широко стайность распространена среди рыб и птиц, хотя встречается и у млекопитающих (например, у собак).
Стадо – длительное или постоянное объединение животных, в котором осуществляются все основные функции жизни вида: добывание корма, защита от хищников, миграция, размножение, воспитание молодняка.
Основу группового поведения в стадах составляют взаимоотношения доминирования. Характерно наличие временного или относительно постоянного лидера, которому подражают особи стада.
Активное руководство стадом (специальные сигналы или угрозы) осуществляют вожаки. Иерархически организованному стаду свойственен закономерный порядок перемещения, определенные позиции при защите от врагов, расположение на местах отдыха и др. (рис. 4.3).
Колония – это групповое поселение оседлых животных на длительное время или на период размножения. По сложности взаимоотношений между особями колонии очень разнообразны, наиболее сложные отношения складываются в поселениях для общественных насекомых (термитов, муравьев, пчел, ос и др.), возникающие на основе сильно разросшейся семьи. Члены колоний постоянно обмениваются информацией друг с другом.
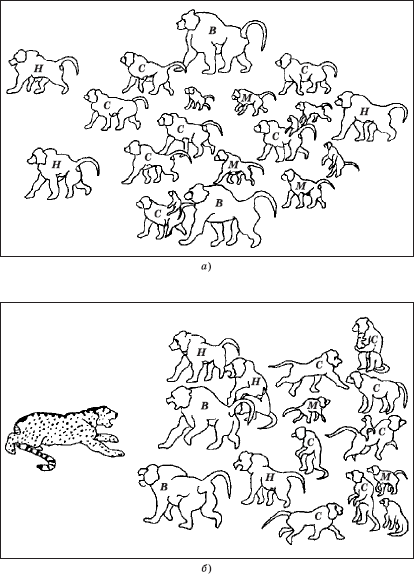 Рис. 4.3. Походный порядок стада павианов: а – в пути; б – при нападении хищника (по Ф. П. Фуэнтэ); В – вожаки; Н – самцы низкого ранга; С – самки; М – молодняк
Рис. 4.3. Походный порядок стада павианов: а – в пути; б – при нападении хищника (по Ф. П. Фуэнтэ); В – вожаки; Н – самцы низкого ранга; С – самки; М – молодняк
Случайное распределение (рис. 4.2, а) в природе встречается редко; оно наблюдается в случаях, когда среда очень однородна, а организмы не стремятся объединиться в группы. Равномерное распределение (рис. 4.2, б) бывает там, где между особями очень сильна конкуренция или существует антагонизм. Наиболее часто наблюдается неравномерное (групповое) распределение (рис. 4.2, в) – образование различных скоплений.
Активность особей, пар и семейных групп у позвоночных и высших беспозвоночных обычно ограничена определенной зоной, называемой индивидуальным (или семейным) участком территории. У высших животных внутрипопуляционное распределение регулируется системой инстинктов. Им свойственно особое территориальное поведение – реакция на местонахождение других членов популяции.
В зависимости от характера использования пространства подвижных животных подразделяют на о с е д л ы х и к о ч е в ы х. Оседлые животные в течение всей или большей части жизни используют довольно ограниченный участок среды. Им присущи инстинкты привязанности к своему участку, регулярное возвращение к месту размножения после длительных и дальних миграций.
Кочевые животные совершают постоянные передвижения в пространстве, так как они зависят от запаса корма на данной территории. Кочевой образ жизни характерен преимуществен
но для стад и стай (кочующие группы многих рыб во время нагульных миграций, стада слонов, зебр, антилоп, северных оленей и т. п.). Перемещение кочующих видов по площади обычно происходит за сроки, достаточные для восстановления кормовых ресурсов на пастбищных участках. Масштабы и длительность таких миграций определяют обилие пищи и численность стада.
Этологическая (поведенческая) структура. Она отражает разнообразные формы совместного существования особей в популяциях. О д и н о ч н ы й о б р а з жизни следует выделить в первую очередь, хотя полностью одиночного существования организмов в природе нет, так как в этом случае было бы невозможно размножение. С е м е й н ы й о б р а з жизни – усиливаются связи между родителями и потомством, начинает заметно проявляться территориальное поведение животных. Путем различных сигналов, маркировки, угроз и тому подобного обеспечивается владение участком, достаточным для выкармливания потомства.
Стая – временное объединение животных, проявляющих биологически полезную организованность действий (для защиты от врагов, добычи пищи, миграции и т. п.). Наиболее широко стайность распространена среди рыб и птиц, хотя встречается и у млекопитающих (например, у собак).
Стадо – длительное или постоянное объединение животных, в котором осуществляются все основные функции жизни вида: добывание корма, защита от хищников, миграция, размножение, воспитание молодняка.
Основу группового поведения в стадах составляют взаимоотношения доминирования. Характерно наличие временного или относительно постоянного лидера, которому подражают особи стада.
Активное руководство стадом (специальные сигналы или угрозы) осуществляют вожаки. Иерархически организованному стаду свойственен закономерный порядок перемещения, определенные позиции при защите от врагов, расположение на местах отдыха и др. (рис. 4.3).
Колония – это групповое поселение оседлых животных на длительное время или на период размножения. По сложности взаимоотношений между особями колонии очень разнообразны, наиболее сложные отношения складываются в поселениях для общественных насекомых (термитов, муравьев, пчел, ос и др.), возникающие на основе сильно разросшейся семьи. Члены колоний постоянно обмениваются информацией друг с другом.
4.4. Динамика популяций
Динамика популяций – это процессы изменения ее основных биологических показателей (численности, биомассы, структуры) во времени в зависимости от экологических факторов. Жизнь популяции проявляется в ее динамике – одном из наиболее значимых биологических и экологических явлений.
4.4.1. Кривые выживания
Кривая выживания представляет собой график зависимости от времени числа выживших на данный момент особей для некоторой начальной группы новорожденных. Каждому биологическому виду свойственна своя характерная кривая выживания. При построении графика по оси абсцисс откладывают относительный возраст, а по оси ординат – абсолютное число выживших особей или их процент от исходного числа по формуле:
 Типичные примеры представлены на рис. 4.4.
Типичные примеры представлены на рис. 4.4.
Все животные и растения подвержены старению, которое проявляется в снижении жизненной активности с возрастом после периода зрелости. Непосредственные причины смерти могут быть разными, но в основе их лежит уменьшение сопротивляемости организма к неблагоприятным факторам, например болезням.
Кривая 1 на рис. 4.4 близка к идеальной кривой выживания для популяции, в которой старение – главный фактор, влияющий на смертность. Так, в современной развитой стране с высоким уровнем медицинского обслуживания и рациональным питанием большинство людей доживает до старости, но среднюю ожидаемую продолжительность жизни почти невозможно увеличить более чем до 75 лет. Поскольку даже в наиболее высокоразвитых странах детская смертность выше средней, она изображается начальным участком а, а наличие долгожителей – конечным участком б (кривая 1). Кроме того, на кривую выживания влияет фактор «случайной гибели», причины которой с возрастом могут меняться. Например, в Англии наибольшая смертность в результате автомобильных аварий приходится на возраст 20–25 лет.
Закономерность, аналогичная кривой 1, свойственна и однолетним растениям, например злаковым, ибо на поле все растения одного вида стареют одновременно.
Кривая 3 иллюстрирует изменения в популяции, у которой смертность относительно постоянна в течение всей жизни организмов (например, 50 % за определенное время). Причиной смерти преимущественно служит случай, и особи гибнут до начала заметного старения. Кривая, подобная этой, была получена, например, для «популяции» стеклянных (легко бьющихся) стаканов в кафетерии. В природе к таким видам относятся, например, устрицы, дающие огромное потомство, а также растения, размножающиеся благодаря рассеиванию большого числа семян. Большая или меньшая смертность среди молодых особей приводит к изменению крутизны опускания начальной части кривой 3.
К промежуточному типу относят кривые выживания таких видов, для которых смертность мало меняется с возрастом. В природе существует много видов птиц, ящериц, мелких млекопитающих и других организмов, имеющих кривые выживания 2, которые отличаются от прямолинейной диагонали некоторой выпуклостью (вогнутостью) или волнообразностью.
Форма кривой выживания часто меняется при изменении плотности популяции. С увеличением плотности кривая становится более вогнутой, т. е. при увеличении численности организмов их смертность возрастает.
Существуют также внутривидовые различия в кривых выживания. Причины могут быть разные и нередко связаны с полом. Так, женщины живут дольше мужчин, хотя причины этого точно не известны.
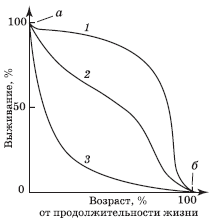 Рис. 4.4. Кривые выживания: 1 – человек или крупные млекопитающие; 2 – гидра или чернохвостый олень; 3 – устрица или рыба
Рис. 4.4. Кривые выживания: 1 – человек или крупные млекопитающие; 2 – гидра или чернохвостый олень; 3 – устрица или рыба
Все животные и растения подвержены старению, которое проявляется в снижении жизненной активности с возрастом после периода зрелости. Непосредственные причины смерти могут быть разными, но в основе их лежит уменьшение сопротивляемости организма к неблагоприятным факторам, например болезням.
Кривая 1 на рис. 4.4 близка к идеальной кривой выживания для популяции, в которой старение – главный фактор, влияющий на смертность. Так, в современной развитой стране с высоким уровнем медицинского обслуживания и рациональным питанием большинство людей доживает до старости, но среднюю ожидаемую продолжительность жизни почти невозможно увеличить более чем до 75 лет. Поскольку даже в наиболее высокоразвитых странах детская смертность выше средней, она изображается начальным участком а, а наличие долгожителей – конечным участком б (кривая 1). Кроме того, на кривую выживания влияет фактор «случайной гибели», причины которой с возрастом могут меняться. Например, в Англии наибольшая смертность в результате автомобильных аварий приходится на возраст 20–25 лет.
Закономерность, аналогичная кривой 1, свойственна и однолетним растениям, например злаковым, ибо на поле все растения одного вида стареют одновременно.
Кривая 3 иллюстрирует изменения в популяции, у которой смертность относительно постоянна в течение всей жизни организмов (например, 50 % за определенное время). Причиной смерти преимущественно служит случай, и особи гибнут до начала заметного старения. Кривая, подобная этой, была получена, например, для «популяции» стеклянных (легко бьющихся) стаканов в кафетерии. В природе к таким видам относятся, например, устрицы, дающие огромное потомство, а также растения, размножающиеся благодаря рассеиванию большого числа семян. Большая или меньшая смертность среди молодых особей приводит к изменению крутизны опускания начальной части кривой 3.
К промежуточному типу относят кривые выживания таких видов, для которых смертность мало меняется с возрастом. В природе существует много видов птиц, ящериц, мелких млекопитающих и других организмов, имеющих кривые выживания 2, которые отличаются от прямолинейной диагонали некоторой выпуклостью (вогнутостью) или волнообразностью.
Форма кривой выживания часто меняется при изменении плотности популяции. С увеличением плотности кривая становится более вогнутой, т. е. при увеличении численности организмов их смертность возрастает.
Существуют также внутривидовые различия в кривых выживания. Причины могут быть разные и нередко связаны с полом. Так, женщины живут дольше мужчин, хотя причины этого точно не известны.
4.4.2. Рост популяции и кривые роста
Если при незначительной эмиграции и иммиграции рождаемость превышает смертность, то популяция будет расти. Рост популяции является непрерывным процессом, если в ней существуют все возрастные группы. Скорость роста популяции при отсутствии каких-либо экологических ограничений описывает дифференциальное уравнение:
dN/dτ = rN, (4.1)
где N – численность особей в популяции; τ – время; r – константа скорости естественного прироста.
J-образная модель роста популяции. Если r > 0, то со временем численность популяции становится больше. Рост происходит сначала медленно, а затем стремительно увеличивается по экспоненциальному закону, т. е. кривая роста популяции принимает J-образный вид (рис. 4.5, а). Такая модель основывается на допущении, что рост популяции не зависит от ее плотности. Считают, что почти любой вид теоретически способен увеличить свою численность до заселения всей Земли при достатке пищи, воды, пространства, постоянстве условий среды и отсутствии хищников. Эта идея была выдвинута еще на рубеже XVIII и XIX вв. английским экономистом Томасом Р. Мальтусом, основоположником теории мальтузианства.
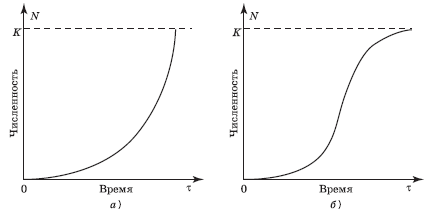 Рис. 4.5. Типы кривых роста численности популяции (модели роста популяции): а – J-образная; б – S-образная; K – поддерживающая емкость среды
Рис. 4.5. Типы кривых роста численности популяции (модели роста популяции): а – J-образная; б – S-образная; K – поддерживающая емкость среды
«S-образная модель роста популяции. Иное развитие получает ситуация при ограниченности пищевых ресурсов либо при скоплении токсичных продуктов (отходов) метаболизма. Первоначальный экспоненциальный рост в исходных благоприятных условиях со временем продолжаться не может и постепенно замедляется. Плотность популяции регулирует истощение пищевых ресурсов, накопление токсикантов и поэтому влияет на рост численности. С увеличением плотности скорость роста популяции постепенно снижается до нуля, и кривая выходит на некоторый стабильный уровень (график образует плато). Кривая такого роста (рис. 4.5, б) имеет S-образную форму, и поэтому соответствующая модель развития событий называется S-образной. Она характерна, например, для дрожжей, фактором, ограничивающим их рост, является накопление спирта, а также для водорослей, самозатеняющих друг друга. В обоих случаях численность популяции не достигает уровня, на котором начинает сказываться нехватка элементов питания (биогенов).
На рост численности, в которой значительную (возможно, даже главную) роль играет пространство, также влияет перенаселенность. Лабораторные опыты с крысами показали, что по достижении определенной плотности популяции плодовитость животных резко снижается даже при избытке пищи. Возникают гормональные сдвиги, влияющие на половое поведение; чаще встречается бесплодие, поедание детенышей родителями и т. п. Резко ослабевает родительская забота о потомстве, детеныши раньше покидают гнездо, в результате чего снижается вероятность их выживания. Усиливается агрессивность животных. Подобные явления встречаются также в популяциях ряда млекопитающих, причем не только в лабораторных, но и природных условиях.
dN/dτ = rN, (4.1)
где N – численность особей в популяции; τ – время; r – константа скорости естественного прироста.
J-образная модель роста популяции. Если r > 0, то со временем численность популяции становится больше. Рост происходит сначала медленно, а затем стремительно увеличивается по экспоненциальному закону, т. е. кривая роста популяции принимает J-образный вид (рис. 4.5, а). Такая модель основывается на допущении, что рост популяции не зависит от ее плотности. Считают, что почти любой вид теоретически способен увеличить свою численность до заселения всей Земли при достатке пищи, воды, пространства, постоянстве условий среды и отсутствии хищников. Эта идея была выдвинута еще на рубеже XVIII и XIX вв. английским экономистом Томасом Р. Мальтусом, основоположником теории мальтузианства.
«S-образная модель роста популяции. Иное развитие получает ситуация при ограниченности пищевых ресурсов либо при скоплении токсичных продуктов (отходов) метаболизма. Первоначальный экспоненциальный рост в исходных благоприятных условиях со временем продолжаться не может и постепенно замедляется. Плотность популяции регулирует истощение пищевых ресурсов, накопление токсикантов и поэтому влияет на рост численности. С увеличением плотности скорость роста популяции постепенно снижается до нуля, и кривая выходит на некоторый стабильный уровень (график образует плато). Кривая такого роста (рис. 4.5, б) имеет S-образную форму, и поэтому соответствующая модель развития событий называется S-образной. Она характерна, например, для дрожжей, фактором, ограничивающим их рост, является накопление спирта, а также для водорослей, самозатеняющих друг друга. В обоих случаях численность популяции не достигает уровня, на котором начинает сказываться нехватка элементов питания (биогенов).
На рост численности, в которой значительную (возможно, даже главную) роль играет пространство, также влияет перенаселенность. Лабораторные опыты с крысами показали, что по достижении определенной плотности популяции плодовитость животных резко снижается даже при избытке пищи. Возникают гормональные сдвиги, влияющие на половое поведение; чаще встречается бесплодие, поедание детенышей родителями и т. п. Резко ослабевает родительская забота о потомстве, детеныши раньше покидают гнездо, в результате чего снижается вероятность их выживания. Усиливается агрессивность животных. Подобные явления встречаются также в популяциях ряда млекопитающих, причем не только в лабораторных, но и природных условиях.