Страница:
В. В. Фролов, А. А. Волков, В. В. Анников, О. В. Бейдик
Стоматология собак
ВВЕДЕНИЕ
Если спросить у ветеринарного специалиста, что значит здоровая собака, он всегда будет перечислять все признаки здорового животного – это хорошая подвижность четвероногого питомца, блестящая и лоснящаяся шерсть, чистые и ясные глаза, слегка влажный и холодный нос. Здоровая собака реагирует на зов хозяина, охотно выполняет команды. У нее хороший аппетит, кишечник опорожняется регулярно, мочеиспускание нормальное. Слизистые оболочки бледно-розового цвета. Температура, пульс и дыхание в норме. Однако, перечисляя все признаки здорового состояния животного, зачастую ветеринарный врач упускает из виду характеристику состояния зубов. Обращает внимание на это только в том случае, когда клинические признаки конкретно указывают на патологию в полости рта.
Из всех заболеваний органов и систем болезни зубов являются той группой заболеваний, где чаще всего признаки наблюдаются лишь тогда, когда болезнь зашла слишком далеко, когда затрагиваются не только зубы, но и органы, окружающие их. Поэтому хорошие клинические признаки здоровья собаки не всегда являются достоверными по отношению к состоянию зубов.
Развитие ветеринарной медицины привело к пониманию важности профилактики и лечения ротовой полости у животных. В течение многих столетий стоматологическое исследование как человека, так и животных заключалось только в лечении больных зубов. В медицине человека стоматология выделилась в самостоятельную специальность уже в 1796 г., а концепция профилактики в этой области появилась в конце 1800-х гг. Ветеринарная стоматология начала свое развитие только в течение последних лет. Такое отставание – результат отсутствия внимания к стоматологическим программам в большинстве ветеринарных школ и, как следствие, – слабое понимание важности оральной гигиены домашних животных.
Ветеринарная стоматология тесно связана с другими науками, на базе которых она развивается. Анатомия и физиология – первоначальные слагаемые большого комплекса, без учета которого невозможно изучение патологии органов зубочелюстной системы. Патологическая анатомия и патологическая физиология дают возможность изучать особенности течения воспалительных и других процессов в ротовой полости. Нельзя познать инфекционный процесс в зубной системе и ее защитных приспособлениях без данных по микробиологии, вирусологии и микологии.
Стоматология тесно связана с фармакологией, изучающей фармакодинамику лекарственных средств и наиболее оптимальные условия их применения. Назначение лекарственных средств для лечения зубов и полости рта в целом без учета этиологии и патогенеза болезни может принести больше вреда, чем пользы.
Значение терапии и клинической диагностики необходимо при анализе механизма развития процесса и его влияния на весь организм. Изучение стоматологии позволяет овладеть оперативной техникой лечения при ряде болезней, требующих хирургического вмешательства. Задача состоит в том, чтобы выяснить закономерности течения болезни, найти причину и следствие. Только в этом случае возможны назначение квалифицированного лечения и организация мер профилактики. Включение стоматологии в образовательные программы всех ветеринарных школ способствует расширению знаний и пониманию важности здоровья ротовой полости у домашних животных. Ветеринарные врачи должны включать стоматологию в общую профилактическую программу заботы о здоровье, начиная с первого визита щенка и обеспечивая полное зубоврачебное обслуживание. Это в свою очередь влечет за собой оснащение инструментарием для проведения зубоврачебных процедур, адекватное обучение персонала и многое другое.
Из всех заболеваний органов и систем болезни зубов являются той группой заболеваний, где чаще всего признаки наблюдаются лишь тогда, когда болезнь зашла слишком далеко, когда затрагиваются не только зубы, но и органы, окружающие их. Поэтому хорошие клинические признаки здоровья собаки не всегда являются достоверными по отношению к состоянию зубов.
Развитие ветеринарной медицины привело к пониманию важности профилактики и лечения ротовой полости у животных. В течение многих столетий стоматологическое исследование как человека, так и животных заключалось только в лечении больных зубов. В медицине человека стоматология выделилась в самостоятельную специальность уже в 1796 г., а концепция профилактики в этой области появилась в конце 1800-х гг. Ветеринарная стоматология начала свое развитие только в течение последних лет. Такое отставание – результат отсутствия внимания к стоматологическим программам в большинстве ветеринарных школ и, как следствие, – слабое понимание важности оральной гигиены домашних животных.
Ветеринарная стоматология тесно связана с другими науками, на базе которых она развивается. Анатомия и физиология – первоначальные слагаемые большого комплекса, без учета которого невозможно изучение патологии органов зубочелюстной системы. Патологическая анатомия и патологическая физиология дают возможность изучать особенности течения воспалительных и других процессов в ротовой полости. Нельзя познать инфекционный процесс в зубной системе и ее защитных приспособлениях без данных по микробиологии, вирусологии и микологии.
Стоматология тесно связана с фармакологией, изучающей фармакодинамику лекарственных средств и наиболее оптимальные условия их применения. Назначение лекарственных средств для лечения зубов и полости рта в целом без учета этиологии и патогенеза болезни может принести больше вреда, чем пользы.
Значение терапии и клинической диагностики необходимо при анализе механизма развития процесса и его влияния на весь организм. Изучение стоматологии позволяет овладеть оперативной техникой лечения при ряде болезней, требующих хирургического вмешательства. Задача состоит в том, чтобы выяснить закономерности течения болезни, найти причину и следствие. Только в этом случае возможны назначение квалифицированного лечения и организация мер профилактики. Включение стоматологии в образовательные программы всех ветеринарных школ способствует расширению знаний и пониманию важности здоровья ротовой полости у домашних животных. Ветеринарные врачи должны включать стоматологию в общую профилактическую программу заботы о здоровье, начиная с первого визита щенка и обеспечивая полное зубоврачебное обслуживание. Это в свою очередь влечет за собой оснащение инструментарием для проведения зубоврачебных процедур, адекватное обучение персонала и многое другое.
ГЛАВА I
РАЗВИТИЕ ГОЛОВЫ И ОРГАНОВ РОТОВОЙ ПОЛОСТИ СОБАКИ
В основе ветеринарной стоматологии должны лежать знания развития, строения и физиологии головы, и ротовой полости в частности. Это позволяет ветеринарному врачу представлять картину не только анатомо-топографического строения пасти собаки, физиологических процессов, происходящих в ней, но и выявлять все отклонения органов полости рта, возникшие при внутриутробном развитии или в постнатальный период. Кроме того, знание строения ротовой полости и ее взаимосвязи с соприкасающимися или близлежащими органами дает возможность локализовать патологический процесс в пасти, не дать ему распространиться, применяя более грамотное и необходимое местное или общее лечение. Знание строения и расположения органов полости рта необходимо при оценке рабочих качеств собак. Изменение в строении пасти у щенят, как, например, нарушение прикуса у некоторых пород, приводит к тому, что их выбраковывают из разведения, так как это является наследственным признаком.
Филогенез головы, черепа и ротовой полости
Голова собаки, как и другого животного, является универсальным вместилищем большого ряда органов. Помимо головного мозга как центра регулирования всего организма в целом, в области головы расположены основные органы чувств, такие как зрение, слух, обоняние, вкус. Также в области головы, в ротовой полости, начинаются системы органов пищеварения и дыхания, которые обеспечивают поступление в организм питательных веществ и кислорода, необходимых для его жизнедеятельности.
Историческое развитие (филогенез) головы происходило по тем же принципам, что и любая другая часть тела животного, включая в себя одновременно филогенез различных органов и систем.
Одними из основных принципов филогенетического формообразования головы собаки являются дифференциация (лат. differentia– различие) и интеграция (лат. integratio – возобновление). Принципу дифференциации подвержены все части тела, органы и системы, так как именно этим путем в эволюции животного мира возникает все новое.
Гомогенные части организма с весьма схожими функциями распадаются на органы, выполняющие определенные, более частные функции, и весь организм приобретает более сложное строение. С развитием дифференциации идет разделение органов головы по жизненно важным функциям с одновременным формированием различных систем (пищеварительная, дыхательная).
Обособляясь в отношении одного или нескольких специальных функциональных направлений, части головы становятся в полную зависимость от всех остальных частей тела, несущих иные жизненно важные свойства. Вместе с разделением частей головы по функциям одновременно идет и противоположный процесс – увеличение взаимозависимости участков головы, восстановление гармонического целого, или процесс интеграции.
Сильное изменение в процессе филогенеза головы претерпел череп. У предков позвоночных он появился в связи с дифференциацией переднего края нервной трубки в головной мозг и развитием органов чувств, вызвавших формирование осевого черепа, или нейрокраниума. Дифференциация начальной части кишечной трубки, в свою очередь, вызвала образование, а затем и различные видоизменения висцерального черепа, или спланхнокраниума. Вначале эти два отдела черепа развивались самостоятельно, и лишь в более позднее время они объединились.
Основной опорой головы, как и туловища, у предков позвоночных была хорда. Над ней располагался начальный головной мозг archen-cephalon, заключенный лишь в соединительную капсулу – перепончатый скелет осевого черепа. Под хордой находился начальный отдел системы органов пищеварения и дыхания – головная кишка.
Развитие головного мозга вызвало ряд преобразований осевого черепа, прошедшего три стадии: соединительнотканную, хрящевую и костную. Соединительнотканный (перепончатый) осевой череп, располагающийся над хордой и впереди нее, имеет ряд отверстий для выхода черепномозговых нервов и вен и для входа в черепную полость артерий. В дальнейшем он сменяется хрящевым, а хрящевой – костным черепом. Хрящевой череп имеет вид неравномерно развитой коробки с углублениями, обозначающими границы отдельных областей черепа: затылочной, слуховой, зрительной и обонятельной – образующихся в соответствии с дифференциацией мозга и развитием органов чувств.
В костном черепе формируется большое количество костей. В дальнейшем многие из них сливаются воедино. У млекопитающих их оказывается значительно меньше. Так, непарная затылочная кость образовалась путем соединения четырех затылочных костей, имеющихся у рептилий, а в состав клиновидной кости входит еще большее количество костей. Во взрослом состоянии у млекопитающих различные кости черепа, в свою очередь, срастаются вместе, формируя крышу мозгового отдела черепа – кальварий (лат. calva – лысый).
Висцеральный череп, как и осевой, прошел те же три стадии. Исходным остовом висцерального черепа, располагавшегося под хордой, явились соединительнотканные жаберные дуги. Ископаемые щитковые имели свыше десяти жаберных дуг и сплошной наружный панцирь, состоящий из многих кожных костей, из которых позднее развивались многие покровные кости черепа. В дальнейшем число жаберных дуг уменьшилось до восьми, сократилось и количество покровных костей. Задние жаберные дуги расчленились на четыре подвижных элемента. Передние четыре дуги утратили участие в дыхательной функции и превратились в челюстной аппарат. При этом первые две дуги образовали губные хрящи, третья – челюстную дугу с зубами, состоящую из двух элементов верхнего нёбно-квадратного хряща и нижнего челюстного хряща. Позднее в области челюстной дуги развилось несколько заменяющих ее покровных костей, на некоторых из них появились зубы (рис. 1).
 Рис. 1. Схема филогенетического изменения висцерального скелета головы: а) селяхии, б) собаки; 1 – подъязычно-челюстной хрящ, 1′– квадратная кость, 1′′– наковальня, 2 – нёбно-квадратный хрящ, 3, 2′– челюстной хрящ, 2′′– молоточек, 3′– стремечко, 4 – подъязычный хрящ, 4′– элементы подъязычной кости;I–V – хрящевые жаберные дуги, I′—тело подъязычной кости, II′—III′– щитовидный хрящ, IV′– надгортанных хрящ, V′– черпаловидный и перстневидный хрящи
Рис. 1. Схема филогенетического изменения висцерального скелета головы: а) селяхии, б) собаки; 1 – подъязычно-челюстной хрящ, 1′– квадратная кость, 1′′– наковальня, 2 – нёбно-квадратный хрящ, 3, 2′– челюстной хрящ, 2′′– молоточек, 3′– стремечко, 4 – подъязычный хрящ, 4′– элементы подъязычной кости;I–V – хрящевые жаберные дуги, I′—тело подъязычной кости, II′—III′– щитовидный хрящ, IV′– надгортанных хрящ, V′– черпаловидный и перстневидный хрящи
С переходом животных к существованию на суше количество костей, образующих висцеральный череп, значительно уменьшилось. Часть из них осталась в области лицевого отдела черепа, сохранив связь с системой органов пищеварения и дыхания, а другая часть оттеснилась развившимися покровными костями аборальнее, вошла в состав костей осевого черепа и приобрела иное значение.
Осевой и висцеральный черепа млекопитающих в процессе своего развития становятся в такие тесные взаимоотношения между собой, что кости одного из них входят в состав другого. Поэтому череп взрослых животных делят на мозговой отдел, в котором располагается головной мозг, и на лицевой отдел, составляющий костный остов морды животного, остов носовой и ротовой полостей (рис. 2).
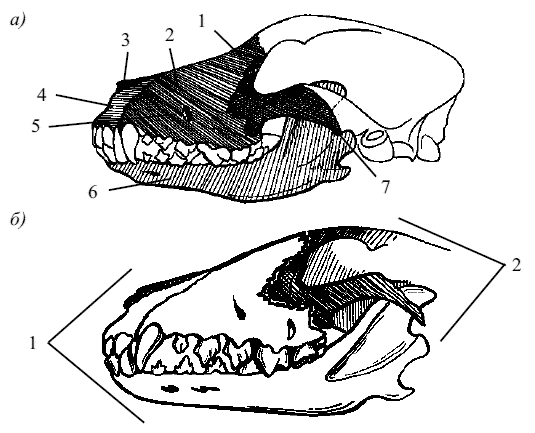 Рис. 2. Отделы черепа собаки: а) лицевой отдел: 1 – слезная кость(os lacrimale), 2 – верхняя челюсть(maxila), 3 – носовая кость(os nasale), 4 – резцовая кость(osincisivum), 5 – подглазное отверстие(foramen infraorbitale), 6 – нижняя челюсть(mandibula), 7 – скуловая кость(os zygomaticum); б) главные отделы: 1 – лицевой отдел, 2 – мозговой отдел
Рис. 2. Отделы черепа собаки: а) лицевой отдел: 1 – слезная кость(os lacrimale), 2 – верхняя челюсть(maxila), 3 – носовая кость(os nasale), 4 – резцовая кость(osincisivum), 5 – подглазное отверстие(foramen infraorbitale), 6 – нижняя челюсть(mandibula), 7 – скуловая кость(os zygomaticum); б) главные отделы: 1 – лицевой отдел, 2 – мозговой отдел
В процессе филогенеза головы произошло формирование ротовой полости с последующим развитием в полости рта ряда органов специального значения, таких как зубы, язык и ротовые железы.
Ротовое отверстие у позвоночных располагается на передней части головы, оно ограничено кожными складками, или губами, которые обыкновенно неподвижны. Только у млекопитающих в губах развивается значительная мускулатура, что позволяет губам быть мясистыми и подвижными. У млекопитающих развивается заметная полость между губами и зубами – преддверие рта. Развитие преддверия ротовой полости позволило животным повысить работоспособность рта для удержания корма, пережевывания, защиты и нападения на жертву. Кроме того, у ряда млекопитающих, таких как обезьяны и грызуны, по бокам преддверия рта образовались защечные мешки.
Сильное развитие получила крыша ротовой полости – нёбо, с первоначальным формированием твердого нёба, его удлинением и образованием мягкого нёба. Если у рыб и у амфибий нёбо образуется за счет основания самого черепа, у ящериц появляются горизонтальные складки, более четко разделяющие носоглоточный ход от собственно ротовой полости, то у млекопитающих нёбо непосредственно разделяет носовую полость от ротовой. Твердое нёбо удлиняется в каудальном направлении и в спадающем виде вместе со слизистой оболочкой и мускулатурой формируют мягкое нёбо. Она отграничивает ротовую полость от глотки. Мягкое нёбо опускается по бокам в виде небноглоточных дуг. По середине между нёбно-глоточными дугами свешивается выступ – язычок. На твердом нёбе животных имеются обычно плотные поперечные пластинки – нёбные валики. Они образовались с целью наилучшего продвижения корма по ротовой полости в пищевод.
В процессе филогенеза зубы позвоночных развиваются путем окостенения сосочков кожи и слизистой оболочки, т. е. являются производными кожи. Процесс закладки зубной системы можно проследить на примере развития зародыша акулы, у которого строение и развитие зубов сходны с плакоидными чешуями, покрывающими кожу этих рыб. Вначале в ротовой полости появляются выросты, соответствующие будущим зубам, с одновременной закладкой кожной чешуи. Закладка зубов и кожной чешуи морфологически ничем не отличается. Затем чешуя, располагающаяся в ротовой полости, увеличивается в размере и приобретает строение зубов с направлением верхушки зуба в полость рта. Характерной особенностью зубов у акул является постоянная их смена в течение всей жизни (плевро-донтная система). Все зубы гомодонтные, или однотипные. У пресмыкающихся более отчетливо формируются нижне– и верхнечелюстные аркады гомодонтных зубов. Наблюдается закрепление нижней части зуба в костной альвеоле с одновременным развитием корня зуба (текодонтная система). Однако у некоторых ящериц зубы укрепляются своими внешними краями к внутреннему краю челюсти, за что получили название плевродонтные зубы. Характерной особенностью зубов у пресмыкающихся является то, что при изнашивании они меняются в течение жизни (полифеодонтизм) и имеют большое количество – 100–200 штук.
У млекопитающих происходит исчезновение гомодонтизма зубов, а на смену приходит гетеродонтизм, т. е. дифференцировка зубов (резцы, клыки, коренные) с характерной особенностью каждого вида зубов. Дифференцировка зубов у млекопитающих сопровождается увеличением их размеров и более прочным прикреплением в отдельных ячейках челюсти. Увеличение размеров этих органов приводит к сокращению их числа и уменьшению смен зубов (рис. 3).
 Рис. 3. Внешний вид зубов собак: 1 – клыки, 2 – резцы, 3 – премоляры, 4 – моляры, 5 – нижняя челюсть
Рис. 3. Внешний вид зубов собак: 1 – клыки, 2 – резцы, 3 – премоляры, 4 – моляры, 5 – нижняя челюсть
Присущее высокоорганизованным позвоночным животным дифиодонтное прорезывание зубов сопровождается процессом дифференциации и специализации отдельных видов зубов, т. е. возникновением гетеродонтного прикуса соответственно степени совершенства позвоночного животного. Дифиодонтная и гетеродонтная системы представляют, таким образом, явления, не только сопутствующие друг другу, но и сопряженные. Примитивные гомодонтные зубы рыб многократно меняются в течение жизни; более совершенные зубы рептилий прорезываются один раз; дифференцированные зубы собак и большинства млекопитающих прорезываются дважды в течение жизни – молочная смена и постоянная.
Получил сильное развитие у млекопитающих корень зуба. Если в зоологическом ряду до высших позвоночных был один корень, то у млекопитающих появляются два и более корней. Кроме того, возникают ложные корни, которые являются непосредственным продолжением коронки в челюсти. Эти ложные корни не отделены от коронки шейкой зуба и обладают перманентным ростом, как, например, клыки у слона. Для всех млекопитающих характерно укрепление зуба по типу вколоченного соединения костей (гомфозис), или зубоальвеолярный состав. За счет разнообразия корма у млекопитающих стала изменяться и форма зубов с характерной для них функцией. У хищных животных сильно развиты клыки, а у травоядных – коренные зубы. Основная форма коренных зубов развивалась из простой конической формы (гаплоидной формы). В дальнейшем к конической форме прибавляются добавочные конусы, формируя трехзубчатую (трикодонтную) форму.
В процессе филогенеза у млекопитающих зубной аппарат приобрел резкое изменение и превратился в жевательный аппарат. Это определялось за счет отложения большого количества минеральных солей и придало органическим частям зуба твердость и неподатливость. Постепенная потеря чувствительности эпителиального покрова привела к тому, что коронка легко выдерживает давление во время жевания и ограждает нижележащие ткани от определенных внешних воздействий. Характер расположения и взаимоотношений составных элементов зубов и особенности сочленения последних с челюстными костями придали зубам высших животных устойчивость, необходимую для выполнения механической функции режущего и размалывающего аппарата. На дне ротовой полости у позвоночных имеется непарный выступ – язык. В наиболее простом виде он впервые появляется у рыб. Язык у этих представителей животного мира представляет собой складку слизистой оболочки, которая поддерживается передним непарным элементом висцерального скелета. У рыб в языке отсутствует собственная мускулатура. Он движется лишь вместе со всем висцеральным аппаратом.
У наземных позвоночных язык приобрел значение подвижного органа, служащего не только для удерживания корма в полости рта, но и для его перемещения в ротовой полости. Вначале это наблюдалось у амфибий за счет появления в языке собственной мускулатуры, являющейся продуктом обособления и дифференцировки подъязычной мускулатуры. В процессе развития мускулатуры языка совершенствовалась и иннервация этого органа. Она осуществляется главным образом за счет подъязычного нерва, отвечающего за подвижность, и языко-глоточного нерва, отвечающего за вкус.
В иннервации языка у рептилий принимает участие, кроме двух ранее названных нервов, еще тройничный нерв. Форма и степень подвижности языка у рептилий в высшей степени различны. У черепах и крокодилов он мало подвижен, а у большинства ящериц и змей он имеет удлиненную форму, на конце обыкновенно глубоко раздвоен и отличается чрезвычайной подвижностью и чувствительностью.
У птиц обычно тонкий и малоподвижный язык лишен внутренней мускулатуры, заострен впереди и отличается сильным развитием рогового слоя, который в задней части языка снабжен роговыми сосочками.
Язык млекопитающих развивается в общем сходно с языком рептилий. Собственная мускулатура достигает наибольшего развития, так что язык оказывается необычайно подвижным и несет разнообразные функции, в особенности выполняет важную роль при акте глотания, а также в качестве вкусового органа. Язык млекопитающих чрезвычайно богат железами и снабжен сосочками различной формы и значения – частью чувствительными, частью ороговевшими. Вообще ороговение языка иногда бывает довольно значительным, как, например у хищников. Ротовые железы, как и другие органы полости рта, имели свое историческое развитие. У рыб и водных амфибий ротовые железы имеют самое примитивное строение по сравнению с другими вышестоящими в зоологическом ряду животными.
У наземных амфибий имеется обыкновенно непарная межносовая железа трубчатого строения, выделяющая слизистый секрет, и нёбные железы. У рептилий, кроме этих желез, имеются еще особые подъязычные, губные, нижние и зубные железы. У ядовитых змей пара задних верхнезубных желез преобразована в сложную трубчатую ядоотделительную железу. Выводные протоки этих желез находятся в связи с ядовитыми зубами. Собственной мускулатурой ядоотделительные железы не обладают, но они прикрыты частью жевательной мышцы, при сокращении которой содержимое железы выдавливается и изливается по протоку к борозде или каналу ядовитого зуба.
Птицы обладают железами на нёбе и подъязычными железами, которые особенно развиты у зерноядных птиц.
У млекопитающих железы полости рта достигают наивысшего развития. Данное развитие касается не только самих желез, но и высокого качества выделяемого ими секрета. Если у нижестоящих животных по сравнению с млекопитающими секрет желез служит первоначально для смачивания слизистой оболочки ротовой полости и в особенности для смачивания корма, то у млекопитающих выделяемая ими слюна содержит не только слизь, но и серозную жидкость. Серозная жидкость содержит пищеварительный фермент, расщепляющий углеводы: птиалин, превращающий крахмал в мальтозу, а мальтоза потом преобразуется в глюкозу. У млекопитающих в полости рта имеется большое количество мелких слизистых желез (губные, нёбные, язычные), которые вполне сравнимы с одноименными железами амфибий и рептилий.
В процессе филогенеза у млекопитающих развились и особенно крупные слюнные железы с одним или несколькими выводными протоками: подъязычная, заднеязычная, подчелюстная и околоушная.
Из этих крупных желез первые три представляют собой, по-видимому, результат дифференцировки подъязычной железы рептилий, а околоушная железа – новое приобретение млекопитающих, развившееся из щечных желез. Кроме того, у различных представителей млекопитающих встречаются и некоторые другие железы полости рта, такие как глазничные железы собак.
Историческое развитие (филогенез) головы происходило по тем же принципам, что и любая другая часть тела животного, включая в себя одновременно филогенез различных органов и систем.
Одними из основных принципов филогенетического формообразования головы собаки являются дифференциация (лат. differentia– различие) и интеграция (лат. integratio – возобновление). Принципу дифференциации подвержены все части тела, органы и системы, так как именно этим путем в эволюции животного мира возникает все новое.
Гомогенные части организма с весьма схожими функциями распадаются на органы, выполняющие определенные, более частные функции, и весь организм приобретает более сложное строение. С развитием дифференциации идет разделение органов головы по жизненно важным функциям с одновременным формированием различных систем (пищеварительная, дыхательная).
Обособляясь в отношении одного или нескольких специальных функциональных направлений, части головы становятся в полную зависимость от всех остальных частей тела, несущих иные жизненно важные свойства. Вместе с разделением частей головы по функциям одновременно идет и противоположный процесс – увеличение взаимозависимости участков головы, восстановление гармонического целого, или процесс интеграции.
Сильное изменение в процессе филогенеза головы претерпел череп. У предков позвоночных он появился в связи с дифференциацией переднего края нервной трубки в головной мозг и развитием органов чувств, вызвавших формирование осевого черепа, или нейрокраниума. Дифференциация начальной части кишечной трубки, в свою очередь, вызвала образование, а затем и различные видоизменения висцерального черепа, или спланхнокраниума. Вначале эти два отдела черепа развивались самостоятельно, и лишь в более позднее время они объединились.
Основной опорой головы, как и туловища, у предков позвоночных была хорда. Над ней располагался начальный головной мозг archen-cephalon, заключенный лишь в соединительную капсулу – перепончатый скелет осевого черепа. Под хордой находился начальный отдел системы органов пищеварения и дыхания – головная кишка.
Развитие головного мозга вызвало ряд преобразований осевого черепа, прошедшего три стадии: соединительнотканную, хрящевую и костную. Соединительнотканный (перепончатый) осевой череп, располагающийся над хордой и впереди нее, имеет ряд отверстий для выхода черепномозговых нервов и вен и для входа в черепную полость артерий. В дальнейшем он сменяется хрящевым, а хрящевой – костным черепом. Хрящевой череп имеет вид неравномерно развитой коробки с углублениями, обозначающими границы отдельных областей черепа: затылочной, слуховой, зрительной и обонятельной – образующихся в соответствии с дифференциацией мозга и развитием органов чувств.
В костном черепе формируется большое количество костей. В дальнейшем многие из них сливаются воедино. У млекопитающих их оказывается значительно меньше. Так, непарная затылочная кость образовалась путем соединения четырех затылочных костей, имеющихся у рептилий, а в состав клиновидной кости входит еще большее количество костей. Во взрослом состоянии у млекопитающих различные кости черепа, в свою очередь, срастаются вместе, формируя крышу мозгового отдела черепа – кальварий (лат. calva – лысый).
Висцеральный череп, как и осевой, прошел те же три стадии. Исходным остовом висцерального черепа, располагавшегося под хордой, явились соединительнотканные жаберные дуги. Ископаемые щитковые имели свыше десяти жаберных дуг и сплошной наружный панцирь, состоящий из многих кожных костей, из которых позднее развивались многие покровные кости черепа. В дальнейшем число жаберных дуг уменьшилось до восьми, сократилось и количество покровных костей. Задние жаберные дуги расчленились на четыре подвижных элемента. Передние четыре дуги утратили участие в дыхательной функции и превратились в челюстной аппарат. При этом первые две дуги образовали губные хрящи, третья – челюстную дугу с зубами, состоящую из двух элементов верхнего нёбно-квадратного хряща и нижнего челюстного хряща. Позднее в области челюстной дуги развилось несколько заменяющих ее покровных костей, на некоторых из них появились зубы (рис. 1).
С переходом животных к существованию на суше количество костей, образующих висцеральный череп, значительно уменьшилось. Часть из них осталась в области лицевого отдела черепа, сохранив связь с системой органов пищеварения и дыхания, а другая часть оттеснилась развившимися покровными костями аборальнее, вошла в состав костей осевого черепа и приобрела иное значение.
Осевой и висцеральный черепа млекопитающих в процессе своего развития становятся в такие тесные взаимоотношения между собой, что кости одного из них входят в состав другого. Поэтому череп взрослых животных делят на мозговой отдел, в котором располагается головной мозг, и на лицевой отдел, составляющий костный остов морды животного, остов носовой и ротовой полостей (рис. 2).
В процессе филогенеза головы произошло формирование ротовой полости с последующим развитием в полости рта ряда органов специального значения, таких как зубы, язык и ротовые железы.
Ротовое отверстие у позвоночных располагается на передней части головы, оно ограничено кожными складками, или губами, которые обыкновенно неподвижны. Только у млекопитающих в губах развивается значительная мускулатура, что позволяет губам быть мясистыми и подвижными. У млекопитающих развивается заметная полость между губами и зубами – преддверие рта. Развитие преддверия ротовой полости позволило животным повысить работоспособность рта для удержания корма, пережевывания, защиты и нападения на жертву. Кроме того, у ряда млекопитающих, таких как обезьяны и грызуны, по бокам преддверия рта образовались защечные мешки.
Сильное развитие получила крыша ротовой полости – нёбо, с первоначальным формированием твердого нёба, его удлинением и образованием мягкого нёба. Если у рыб и у амфибий нёбо образуется за счет основания самого черепа, у ящериц появляются горизонтальные складки, более четко разделяющие носоглоточный ход от собственно ротовой полости, то у млекопитающих нёбо непосредственно разделяет носовую полость от ротовой. Твердое нёбо удлиняется в каудальном направлении и в спадающем виде вместе со слизистой оболочкой и мускулатурой формируют мягкое нёбо. Она отграничивает ротовую полость от глотки. Мягкое нёбо опускается по бокам в виде небноглоточных дуг. По середине между нёбно-глоточными дугами свешивается выступ – язычок. На твердом нёбе животных имеются обычно плотные поперечные пластинки – нёбные валики. Они образовались с целью наилучшего продвижения корма по ротовой полости в пищевод.
В процессе филогенеза зубы позвоночных развиваются путем окостенения сосочков кожи и слизистой оболочки, т. е. являются производными кожи. Процесс закладки зубной системы можно проследить на примере развития зародыша акулы, у которого строение и развитие зубов сходны с плакоидными чешуями, покрывающими кожу этих рыб. Вначале в ротовой полости появляются выросты, соответствующие будущим зубам, с одновременной закладкой кожной чешуи. Закладка зубов и кожной чешуи морфологически ничем не отличается. Затем чешуя, располагающаяся в ротовой полости, увеличивается в размере и приобретает строение зубов с направлением верхушки зуба в полость рта. Характерной особенностью зубов у акул является постоянная их смена в течение всей жизни (плевро-донтная система). Все зубы гомодонтные, или однотипные. У пресмыкающихся более отчетливо формируются нижне– и верхнечелюстные аркады гомодонтных зубов. Наблюдается закрепление нижней части зуба в костной альвеоле с одновременным развитием корня зуба (текодонтная система). Однако у некоторых ящериц зубы укрепляются своими внешними краями к внутреннему краю челюсти, за что получили название плевродонтные зубы. Характерной особенностью зубов у пресмыкающихся является то, что при изнашивании они меняются в течение жизни (полифеодонтизм) и имеют большое количество – 100–200 штук.
У млекопитающих происходит исчезновение гомодонтизма зубов, а на смену приходит гетеродонтизм, т. е. дифференцировка зубов (резцы, клыки, коренные) с характерной особенностью каждого вида зубов. Дифференцировка зубов у млекопитающих сопровождается увеличением их размеров и более прочным прикреплением в отдельных ячейках челюсти. Увеличение размеров этих органов приводит к сокращению их числа и уменьшению смен зубов (рис. 3).
Присущее высокоорганизованным позвоночным животным дифиодонтное прорезывание зубов сопровождается процессом дифференциации и специализации отдельных видов зубов, т. е. возникновением гетеродонтного прикуса соответственно степени совершенства позвоночного животного. Дифиодонтная и гетеродонтная системы представляют, таким образом, явления, не только сопутствующие друг другу, но и сопряженные. Примитивные гомодонтные зубы рыб многократно меняются в течение жизни; более совершенные зубы рептилий прорезываются один раз; дифференцированные зубы собак и большинства млекопитающих прорезываются дважды в течение жизни – молочная смена и постоянная.
Получил сильное развитие у млекопитающих корень зуба. Если в зоологическом ряду до высших позвоночных был один корень, то у млекопитающих появляются два и более корней. Кроме того, возникают ложные корни, которые являются непосредственным продолжением коронки в челюсти. Эти ложные корни не отделены от коронки шейкой зуба и обладают перманентным ростом, как, например, клыки у слона. Для всех млекопитающих характерно укрепление зуба по типу вколоченного соединения костей (гомфозис), или зубоальвеолярный состав. За счет разнообразия корма у млекопитающих стала изменяться и форма зубов с характерной для них функцией. У хищных животных сильно развиты клыки, а у травоядных – коренные зубы. Основная форма коренных зубов развивалась из простой конической формы (гаплоидной формы). В дальнейшем к конической форме прибавляются добавочные конусы, формируя трехзубчатую (трикодонтную) форму.
В процессе филогенеза у млекопитающих зубной аппарат приобрел резкое изменение и превратился в жевательный аппарат. Это определялось за счет отложения большого количества минеральных солей и придало органическим частям зуба твердость и неподатливость. Постепенная потеря чувствительности эпителиального покрова привела к тому, что коронка легко выдерживает давление во время жевания и ограждает нижележащие ткани от определенных внешних воздействий. Характер расположения и взаимоотношений составных элементов зубов и особенности сочленения последних с челюстными костями придали зубам высших животных устойчивость, необходимую для выполнения механической функции режущего и размалывающего аппарата. На дне ротовой полости у позвоночных имеется непарный выступ – язык. В наиболее простом виде он впервые появляется у рыб. Язык у этих представителей животного мира представляет собой складку слизистой оболочки, которая поддерживается передним непарным элементом висцерального скелета. У рыб в языке отсутствует собственная мускулатура. Он движется лишь вместе со всем висцеральным аппаратом.
У наземных позвоночных язык приобрел значение подвижного органа, служащего не только для удерживания корма в полости рта, но и для его перемещения в ротовой полости. Вначале это наблюдалось у амфибий за счет появления в языке собственной мускулатуры, являющейся продуктом обособления и дифференцировки подъязычной мускулатуры. В процессе развития мускулатуры языка совершенствовалась и иннервация этого органа. Она осуществляется главным образом за счет подъязычного нерва, отвечающего за подвижность, и языко-глоточного нерва, отвечающего за вкус.
В иннервации языка у рептилий принимает участие, кроме двух ранее названных нервов, еще тройничный нерв. Форма и степень подвижности языка у рептилий в высшей степени различны. У черепах и крокодилов он мало подвижен, а у большинства ящериц и змей он имеет удлиненную форму, на конце обыкновенно глубоко раздвоен и отличается чрезвычайной подвижностью и чувствительностью.
У птиц обычно тонкий и малоподвижный язык лишен внутренней мускулатуры, заострен впереди и отличается сильным развитием рогового слоя, который в задней части языка снабжен роговыми сосочками.
Язык млекопитающих развивается в общем сходно с языком рептилий. Собственная мускулатура достигает наибольшего развития, так что язык оказывается необычайно подвижным и несет разнообразные функции, в особенности выполняет важную роль при акте глотания, а также в качестве вкусового органа. Язык млекопитающих чрезвычайно богат железами и снабжен сосочками различной формы и значения – частью чувствительными, частью ороговевшими. Вообще ороговение языка иногда бывает довольно значительным, как, например у хищников. Ротовые железы, как и другие органы полости рта, имели свое историческое развитие. У рыб и водных амфибий ротовые железы имеют самое примитивное строение по сравнению с другими вышестоящими в зоологическом ряду животными.
У наземных амфибий имеется обыкновенно непарная межносовая железа трубчатого строения, выделяющая слизистый секрет, и нёбные железы. У рептилий, кроме этих желез, имеются еще особые подъязычные, губные, нижние и зубные железы. У ядовитых змей пара задних верхнезубных желез преобразована в сложную трубчатую ядоотделительную железу. Выводные протоки этих желез находятся в связи с ядовитыми зубами. Собственной мускулатурой ядоотделительные железы не обладают, но они прикрыты частью жевательной мышцы, при сокращении которой содержимое железы выдавливается и изливается по протоку к борозде или каналу ядовитого зуба.
Птицы обладают железами на нёбе и подъязычными железами, которые особенно развиты у зерноядных птиц.
У млекопитающих железы полости рта достигают наивысшего развития. Данное развитие касается не только самих желез, но и высокого качества выделяемого ими секрета. Если у нижестоящих животных по сравнению с млекопитающими секрет желез служит первоначально для смачивания слизистой оболочки ротовой полости и в особенности для смачивания корма, то у млекопитающих выделяемая ими слюна содержит не только слизь, но и серозную жидкость. Серозная жидкость содержит пищеварительный фермент, расщепляющий углеводы: птиалин, превращающий крахмал в мальтозу, а мальтоза потом преобразуется в глюкозу. У млекопитающих в полости рта имеется большое количество мелких слизистых желез (губные, нёбные, язычные), которые вполне сравнимы с одноименными железами амфибий и рептилий.
В процессе филогенеза у млекопитающих развились и особенно крупные слюнные железы с одним или несколькими выводными протоками: подъязычная, заднеязычная, подчелюстная и околоушная.
Из этих крупных желез первые три представляют собой, по-видимому, результат дифференцировки подъязычной железы рептилий, а околоушная железа – новое приобретение млекопитающих, развившееся из щечных желез. Кроме того, у различных представителей млекопитающих встречаются и некоторые другие железы полости рта, такие как глазничные железы собак.
Онтогенез полости рта
Полость рта является начальным отделом пищеварительного тракта и в своем развитии связана со взаимодействием многих структур и процессов. Самый первый этап становления этой полости связан с образованием на головном конце зародыша впячивания кожной эктодермы, или ротовой ямки, дно которой растет навстречу слепому концу передней кишки. Эта ротовая ямка и представляет собой зачаток первичной ротовой полости, а также будущей полости носа. В ходе дальнейшего развития дно ротовой ямки входит в соприкосновение с эндодермой первичной кишки, образуя глоточную, или ротовую, перепонку. На 7-10-й день эмбрионального развития глоточная перепонка прорывается, и с этого момента ротовая ямка вступает в сообщение с полостью первичной кишки. После прорыва глоточной перепонки самая минимальная часть передней кишки присоединяется к ротовой ямке и вместе с ней принимает участие в образовании полости рта (рис. 4).
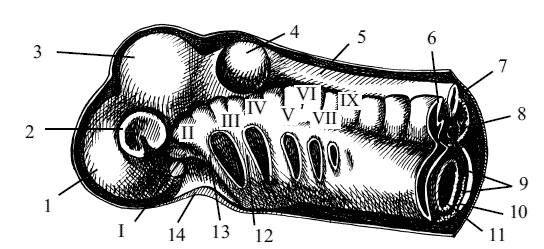 Рис. 4. Стереограмма сегментации мезодермы в области головы плода: 1 – передний мозг, 2 – глазной бокал, 3 – средний мозг, 4 – слуховой пузырь, 5 – нервная трубка, 6 – миотом, 7 – хорда, 8 – склеротом, 9 – боковая планка, или спланхнотом, 10 – полость тела плода, 11 – кишка, 12 – подъязычная перегородка (зачаток дуги), 13 – челюстная перегородка, 14 – ротовая ямка, I–IX – головные сомиты
Рис. 4. Стереограмма сегментации мезодермы в области головы плода: 1 – передний мозг, 2 – глазной бокал, 3 – средний мозг, 4 – слуховой пузырь, 5 – нервная трубка, 6 – миотом, 7 – хорда, 8 – склеротом, 9 – боковая планка, или спланхнотом, 10 – полость тела плода, 11 – кишка, 12 – подъязычная перегородка (зачаток дуги), 13 – челюстная перегородка, 14 – ротовая ямка, I–IX – головные сомиты
Последующее развитие полости рта тесно связано с образованием у зародыша собаки жаберного аппарата, состоящего из набора жаберных дуг, карманов и щелей. Вначале появляются жаберные карманы, представляющие собой выпячивание эндодермы в области боковых стенок глоточного отдела первичной кишки. Навстречу этим выпячиваниям эндодермы растут выпячивания соответствующих отделов эктодермы шейной области зародыша, которые получили название жаберных щелей. Там, где дно жаберных щелей соприкасается с дном жаберных карманов, образуются жаберные перепонки, покрытые снаружи кожным, а изнутри эндодермальным эпителием. Участки мезенхимы, заключенные между соседними жаберными карманами и щелями, разрастаются и образуют на передней поверхности шеи зародыша валикообразные возвышения, называемые жаберными дугами. В мезенхимную основу каждой жаберной дуги врастают кровеносные сосуды. Из жаберных дуг образуются: подъязычная кость, щитовидный хрящ, верхняя и нижняя челюсти. Жаберные карманы формируют нёбные миндалины, полость среднего уха, зачатки околощитовидной и вилочковой желез. Жаберные щели идут на постройку наружных слуховых проходов.
Зачаток ротовой полости снаружи ограничен пятью бугорками: непарным лобным, в основе которого находится развивающийся головной мозг, и двумя парными – верхнечелюстными и нижнечелюстными. Между лобным и верхнечелюстными бугорками проходят слезно-носовые борозды – будущие слезно-носовые каналы. Борозды, отделяющие верхнечелюстные бугорки от нижнечелюстных, в дальнейшем превращаются в углы рта животного (рис. 5).
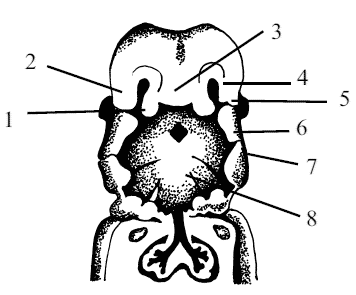
Рис. 5. Онтогенез полости рта: 1 – медиальный носовой отросток, 2 – латеральный носовой отросток, 3 – лобный бугорок, 4 – обонятельные ямки, 5 – слезно-носовая борозда, 6 – верхнечелюстной бугорок, 7 – нижнечелюстной бугорок, 8 – ротовая бухта
На лобном бугорке видны парные углубления – обонятельные ямки. От каждой из них к свободному краю бугорка следует борозда, разделяющая этот край на латеральный и медиальный носовые отростки. В дальнейшем лобный отросток срастается с верхнечелюстным, формируя верхнюю челюсть и губу. Границы этих зачатков впоследствии проходят по швам между верхнечелюстными и резцовыми костями. Нижнечелюстные бугорки срастаются в нижнюю челюсть и губу. Обонятельные ямки со временем углубляются и прорываются в ротовую полость, превращаясь в каналы, наружные отверстия которых называются первичными ноздрями, а внутренние – первичными хоанами. Полость рта на этой стадии развития становится первичной ротовой полостью.
Последующее развитие полости рта тесно связано с образованием у зародыша собаки жаберного аппарата, состоящего из набора жаберных дуг, карманов и щелей. Вначале появляются жаберные карманы, представляющие собой выпячивание эндодермы в области боковых стенок глоточного отдела первичной кишки. Навстречу этим выпячиваниям эндодермы растут выпячивания соответствующих отделов эктодермы шейной области зародыша, которые получили название жаберных щелей. Там, где дно жаберных щелей соприкасается с дном жаберных карманов, образуются жаберные перепонки, покрытые снаружи кожным, а изнутри эндодермальным эпителием. Участки мезенхимы, заключенные между соседними жаберными карманами и щелями, разрастаются и образуют на передней поверхности шеи зародыша валикообразные возвышения, называемые жаберными дугами. В мезенхимную основу каждой жаберной дуги врастают кровеносные сосуды. Из жаберных дуг образуются: подъязычная кость, щитовидный хрящ, верхняя и нижняя челюсти. Жаберные карманы формируют нёбные миндалины, полость среднего уха, зачатки околощитовидной и вилочковой желез. Жаберные щели идут на постройку наружных слуховых проходов.
Зачаток ротовой полости снаружи ограничен пятью бугорками: непарным лобным, в основе которого находится развивающийся головной мозг, и двумя парными – верхнечелюстными и нижнечелюстными. Между лобным и верхнечелюстными бугорками проходят слезно-носовые борозды – будущие слезно-носовые каналы. Борозды, отделяющие верхнечелюстные бугорки от нижнечелюстных, в дальнейшем превращаются в углы рта животного (рис. 5).
Рис. 5. Онтогенез полости рта: 1 – медиальный носовой отросток, 2 – латеральный носовой отросток, 3 – лобный бугорок, 4 – обонятельные ямки, 5 – слезно-носовая борозда, 6 – верхнечелюстной бугорок, 7 – нижнечелюстной бугорок, 8 – ротовая бухта
На лобном бугорке видны парные углубления – обонятельные ямки. От каждой из них к свободному краю бугорка следует борозда, разделяющая этот край на латеральный и медиальный носовые отростки. В дальнейшем лобный отросток срастается с верхнечелюстным, формируя верхнюю челюсть и губу. Границы этих зачатков впоследствии проходят по швам между верхнечелюстными и резцовыми костями. Нижнечелюстные бугорки срастаются в нижнюю челюсть и губу. Обонятельные ямки со временем углубляются и прорываются в ротовую полость, превращаясь в каналы, наружные отверстия которых называются первичными ноздрями, а внутренние – первичными хоанами. Полость рта на этой стадии развития становится первичной ротовой полостью.