Страница:
7. Правило ограничивающих факторов. Возможности существования организмов в первую очередь ограничивают те факторы среды, которые наиболее удаляются от оптимума. Если хотя бы один из экологических факторов приближается или выходит за пределы критических величин, то, несмотря на оптимальное сочетание остальных условий, особям грозит гибель. Любые сильно уклоняющиеся от оптимума факторы приобретают первостепенное значение в жизни вида или отдельных его представителей в конкретные отрезки времени.
Ограничивающие факторы среды определяют географический ареал вида. Природа этих факторов может быть различной (рис. 6). Так, продвижение вида на север может лимитироваться недостатком тепла, в аридные районы – недостатком влаги или слишком высокими температурами. Ограничивающим распространение фактором могут служить и биотические отношения, например занятость территории более сильным конкурентом или недостаток опылителей для растений. Так, опыление инжира всецело зависит от единственного вида насекомых – осы Blastophaga psenes. Родина этого дерева – Средиземноморье. Завезенный в Калифорнию инжир не плодоносил до тех пор, пока туда не завезли ос-опылителей. Распространение бобовых в Арктике ограничивается распределением опыляющих их шмелей. На острове Диксон, где нет шмелей, не встречаются и бобовые, хотя по температурным условиям существование там этих растений еще допустимо.

Рис. 6. Глубокий снежный покров – лимитирующий фактор в распространении оленей (по Г. А. Новикову, 1981)
Чтобы определить, сможет ли вид существовать в данном географическом районе, нужно в первую очередь выяснить, не выходят ли какие-либо факторы среды за пределы его экологической валентности, особенно в наиболее уязвимый период развития.
Выявление ограничивающих факторов очень важно в практике сельского хозяйства, так как, направив основные усилия на их устранение, можно быстро и эффективно повысить урожайность растений или производительность животных. Так, на сильно кислых почвах урожай пшеницы можно несколько увеличить, применяя разные агрономические воздействия, но наилучший эффект будет получен только в результате известкования, которое снимет ограничивающие действия кислотности. Знание ограничивающих факторов, таким образом, ключ к управлению жизнедеятельностью организмов. В разные периоды жизни особей в качестве ограничивающих выступают различные факторы среды, поэтому требуется умелое и постоянное регулирование условий жизни выращиваемых растений и животных.
2.4. Принципы экологической классификации организмов
2.5. Активная и скрытая жизнь
Глава 3. ВАЖНЕЙШИЕ АБИОТИЧЕСКИЕ ФАКТОРЫ И АДАПТАЦИИ К НИМ ОРГАНИЗМОВ
3.1. Температура
3.1.1. Температурные границы существования видов
Ограничивающие факторы среды определяют географический ареал вида. Природа этих факторов может быть различной (рис. 6). Так, продвижение вида на север может лимитироваться недостатком тепла, в аридные районы – недостатком влаги или слишком высокими температурами. Ограничивающим распространение фактором могут служить и биотические отношения, например занятость территории более сильным конкурентом или недостаток опылителей для растений. Так, опыление инжира всецело зависит от единственного вида насекомых – осы Blastophaga psenes. Родина этого дерева – Средиземноморье. Завезенный в Калифорнию инжир не плодоносил до тех пор, пока туда не завезли ос-опылителей. Распространение бобовых в Арктике ограничивается распределением опыляющих их шмелей. На острове Диксон, где нет шмелей, не встречаются и бобовые, хотя по температурным условиям существование там этих растений еще допустимо.
Рис. 6. Глубокий снежный покров – лимитирующий фактор в распространении оленей (по Г. А. Новикову, 1981)
Чтобы определить, сможет ли вид существовать в данном географическом районе, нужно в первую очередь выяснить, не выходят ли какие-либо факторы среды за пределы его экологической валентности, особенно в наиболее уязвимый период развития.
Выявление ограничивающих факторов очень важно в практике сельского хозяйства, так как, направив основные усилия на их устранение, можно быстро и эффективно повысить урожайность растений или производительность животных. Так, на сильно кислых почвах урожай пшеницы можно несколько увеличить, применяя разные агрономические воздействия, но наилучший эффект будет получен только в результате известкования, которое снимет ограничивающие действия кислотности. Знание ограничивающих факторов, таким образом, ключ к управлению жизнедеятельностью организмов. В разные периоды жизни особей в качестве ограничивающих выступают различные факторы среды, поэтому требуется умелое и постоянное регулирование условий жизни выращиваемых растений и животных.
2.4. Принципы экологической классификации организмов
Современная систематика растений и животных построена на основе единственного главного критерия – степени родства организмов. При этом внешние особенности видов, относимых к одной группе, часто могут сильно различаться. Так, например, паразит крабов саккулина, напоминающая бесформенный, набитый половыми продуктами мешок с сильно разветвленной в теле хозяина сетью тяжей, внешне совершенно не похожа на сидячих, обладающих раковинами морских желудей и морских уточек, хотя все они относятся к одному отряду усоногих раков. О родстве этих видов говорит глубокое внутреннее сходство, прослеживаемое па первых этапах развития особей.
В экологии разнообразие и разноплановость способов и путей адаптации к среде создают необходимость множественных классификаций. Используя какой-либо единственный критерий, нельзя отразить все стороны приспособленности организмов к среде. Экологические классификации отражают сходство, возникающее у представителей самых разных групп, если они используют сходные пути адаптации. Например, если мы классифицируем животных по способам движения, то в экологическую группу видов, передвигающихся в воде реактивным путем, попадут такие разные по систематическому положению животные, как медузы, головоногие моллюски, некоторые инфузории и жгутиковые, личинки ряда стрекоз и др. (рис. 7). В основу экологических классификаций могут быть положены самые разнообразные критерии: способы питания, передвижения, отношение к температуре, влажности, солености среды, давлению и т. п. Разделение всех организмов на эврибионтных и стенобионтных по широте диапазона приспособлений к среде представляет пример простейшей экологической классификации.
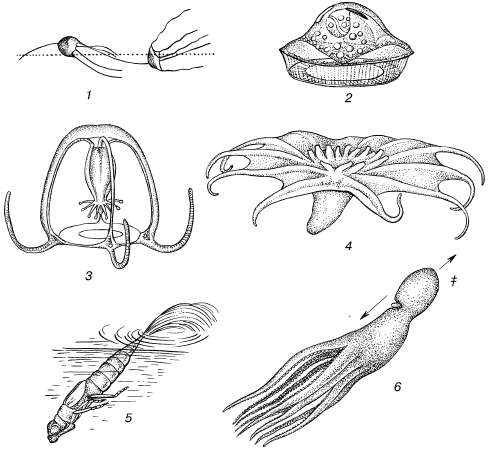
Рис. 7. Представители экологической группы организмов, передвигающихся в воде реактивным способом (по С. A. Зернову, 1949):
1– жгутиковое Medusochloris phiale;
2– инфузория Craspedotella pileosus;
3– медуза Cytaeis vulgaris;
4– пелагическая голотурия Pelagothuria;
5– личинка стрекозы-коромысла;
6– плывущий осьминог Octopus vulgaris:
а– направление струи воды;
б– направление движения животного
Другой пример – разделение организмов на группы по характеру питания. Автотрофы – это организмы, использующие в качестве источника для построения своего тела неорганические соединения. Гетеротрофы – все живые существа, нуждающиеся в пище органического происхождения. В свою очередь, автотрофы делятся на фототрофов и хемотрофов. Первые для синтеза органических молекул используют энергию солнечного света, вторые – энергию химических связей. Гетеротрофов делят на сапрофитов, использующих растворы простых органических соединений, и голозоев. Голозои обладают сложным комплексом пищеварительных ферментов и могут употреблять в пищу сложные органические соединения, разлагая их на более простые составные компоненты. Голозои делятся на сапрофагов (питаются мертвыми растительными остатками), фитофагов (потребителей живых растений), зоофагов (нуждающихся в живой пище) и некрофагов (трупоядных животных). В свою очередь, каждую из этих групп можно подразделить на более мелкие, имеющие свою специфику в характере питания.
Иначе можно построить классификацию по способу добывания пищи. Среди животных выявляются, например, такие группы, как филътраторы (мелкие рачки, беззубка, кит и др.), пасущиеся формы (копытные, жуки-листоеды), собиратели (дятлы, кроты, землеройки, куриные), охотники на движущуюся добычу (волки, львы, мухи-ктыри и т. п.) и целый ряд других групп. Так, несмотря на большое несходство в организации, одинаковый способ овладения добычей приводит у львов и мух-ктырей к ряду аналогий в их охотничьих повадках и общих чертах строения: поджарости тела, сильному развитию мускулатуры, способности развивать кратковременно большую скорость и т. п.
Экологические классификации помогают выявлять возможные в природе пути приспособления организмов к среде.
В экологии разнообразие и разноплановость способов и путей адаптации к среде создают необходимость множественных классификаций. Используя какой-либо единственный критерий, нельзя отразить все стороны приспособленности организмов к среде. Экологические классификации отражают сходство, возникающее у представителей самых разных групп, если они используют сходные пути адаптации. Например, если мы классифицируем животных по способам движения, то в экологическую группу видов, передвигающихся в воде реактивным путем, попадут такие разные по систематическому положению животные, как медузы, головоногие моллюски, некоторые инфузории и жгутиковые, личинки ряда стрекоз и др. (рис. 7). В основу экологических классификаций могут быть положены самые разнообразные критерии: способы питания, передвижения, отношение к температуре, влажности, солености среды, давлению и т. п. Разделение всех организмов на эврибионтных и стенобионтных по широте диапазона приспособлений к среде представляет пример простейшей экологической классификации.
Рис. 7. Представители экологической группы организмов, передвигающихся в воде реактивным способом (по С. A. Зернову, 1949):
1– жгутиковое Medusochloris phiale;
2– инфузория Craspedotella pileosus;
3– медуза Cytaeis vulgaris;
4– пелагическая голотурия Pelagothuria;
5– личинка стрекозы-коромысла;
6– плывущий осьминог Octopus vulgaris:
а– направление струи воды;
б– направление движения животного
Другой пример – разделение организмов на группы по характеру питания. Автотрофы – это организмы, использующие в качестве источника для построения своего тела неорганические соединения. Гетеротрофы – все живые существа, нуждающиеся в пище органического происхождения. В свою очередь, автотрофы делятся на фототрофов и хемотрофов. Первые для синтеза органических молекул используют энергию солнечного света, вторые – энергию химических связей. Гетеротрофов делят на сапрофитов, использующих растворы простых органических соединений, и голозоев. Голозои обладают сложным комплексом пищеварительных ферментов и могут употреблять в пищу сложные органические соединения, разлагая их на более простые составные компоненты. Голозои делятся на сапрофагов (питаются мертвыми растительными остатками), фитофагов (потребителей живых растений), зоофагов (нуждающихся в живой пище) и некрофагов (трупоядных животных). В свою очередь, каждую из этих групп можно подразделить на более мелкие, имеющие свою специфику в характере питания.
Иначе можно построить классификацию по способу добывания пищи. Среди животных выявляются, например, такие группы, как филътраторы (мелкие рачки, беззубка, кит и др.), пасущиеся формы (копытные, жуки-листоеды), собиратели (дятлы, кроты, землеройки, куриные), охотники на движущуюся добычу (волки, львы, мухи-ктыри и т. п.) и целый ряд других групп. Так, несмотря на большое несходство в организации, одинаковый способ овладения добычей приводит у львов и мух-ктырей к ряду аналогий в их охотничьих повадках и общих чертах строения: поджарости тела, сильному развитию мускулатуры, способности развивать кратковременно большую скорость и т. п.
Экологические классификации помогают выявлять возможные в природе пути приспособления организмов к среде.
2.5. Активная и скрытая жизнь
Обмен веществ – одно из главнейших свойств жизни, определяющее тесную вещественно-энергетическую связь организмов со средой. Метаболизм проявляет сильную зависимость от условий существования. В природе мы наблюдаем два основных состояния жизни: активную жизнедеятельность и покой. При активной жизнедеятельности организмы питаются, растут, передвигаются, развиваются, размножаются, характеризуясь при этом интенсивным метаболизмом. Покой может быть разным по глубине и продолжительности, многие функции организма при этом ослабевают или не выполняются совсем, так как уровень обмена веществ падает под влиянием внешних и внутренних факторов.
В состоянии глубокого покоя, т. е. пониженного вещественно-энергетического обмена, организмы становятся менее зависимыми от среды, приобретают высокую степень устойчивости и способны переносить условия, которые не могли бы выдержать при активной жизнедеятельности. Эти два состояния чередуются в жизни многих видов, являясь адаптацией к местообитаниям с нестабильным климатом, резкими сезонными изменениями, что характерно для большей части планеты.
При глубоком подавлении обмена веществ организмы могут вообще не проявлять видимых признаков жизни. Вопрос о том, возможна ли полная остановка обмена веществ с последующим возвращением к активной жизнедеятельности, т. е. своего рода «воскрешение из мертвых», дискутировался в науке более двух столетий.
Впервые явление мнимой смерти было обнаружено в 1702 г. Антони ван Левенгуком – открывателем микроскопического мира живых существ. Наблюдаемые им «анималькули» (коловратки) при высыхании капли воды сморщивались, выглядели мертвыми и могли пребывать в таком состоянии длительное время (рис. 8). Помещенные вновь в воду, они набухали и переходили к активной жизни. Левенгук объяснил это явление тем, что оболочка «анималькулей», очевидно, «не позволяет ни малейшего испарения» и они остаются живыми в сухих условиях. Однако через несколько десятилетий естествоиспытатели уже спорили о возможности того, что «жизнь может быть полностью прекращена» и восстановлена вновь «через 20, 40, 100 лет или более».
В 70-х годах XVIII в. явление «воскрешения» после высыхания было обнаружено и подтверждено многочисленными опытами у ряда других мелких организмов – пшеничных угриц, свободноживущих нематод и тихоходок. Ж. Бюффон, повторив опыты Дж. Нидгема с угрицами, утверждал, что «эти организмы можно заставить сколько угодно раз подряд умирать и вновь оживать». Л. Спалланцани впервые обратил внимание на глубокий покой семян и спор растений, расценив его как сохранение их во времени.
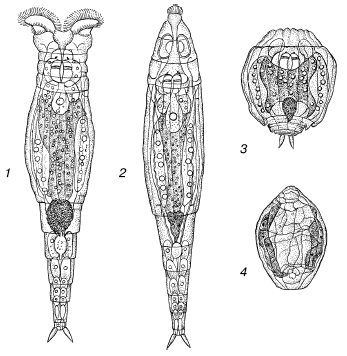
Рис. 8. Коловратка Philidina roseola на разных стадиях высыхания (по П. Ю. Шмидту, 1948):
1– активная; 2 – начинающая сокращаться; 3 – полностью сократившаяся перед высыханием; 4 – в состоянии анабиоза
В середине XIX в. было убедительно установлено, что устойчивость сухих коловраток, тихоходок и нематод к высоким и низким температурам, недостатку или отсутствию кислорода возрастает пропорционально степени их обезвоживания. Однако оставался открытым вопрос, происходит ли при этом полное прерывание жизни или лишь ее глубокое угнетение. В 1878 г. Клод Бернал выдвинул понятие «скрытая жизнь», которую он характеризовал прекращением обмена веществ и «перерывом отношений между существом и средой».
Окончательно этот вопрос был решен лишь в первой трети XX столетия с развитием техники глубокого вакуумного обезвоживания. Опыты Г. Рама, П. Беккереля и других ученых показали возможность полной обратимой остановки жизни. В сухом состоянии, когда в клетках оставалось не более 2 % воды в химически связанном виде, такие организмы, как коловратки, тихоходки, мелкие нематоды, семена и споры растений, споры бактерий и грибов выдерживали пребывание в жидком кислороде (-218,4 °C), жидком водороде (-259,4 °C), жидком гелии (-269,0 °C), т. е. температуры, близкие к абсолютному нулю. При этом содержимое клеток затвердевает, отсутствует даже тепловое движение молекул, и всякий обмен веществ, естественно, прекращен. После помещения в нормальные условия эти организмы продолжают развитие. У некоторых видов остановка обмена веществ при сверхнизких температурах возможна и без высушивания, при условии замерзания воды не в кристаллическом, а в аморфном состоянии.
Полная временная остановка жизни получила название анабиоза. Термин был предложен В. Прейером еще в 1891 г. В состоянии анабиоза организмы становятся устойчивыми к самым разнообразным воздействиям. Например, тихоходки выдерживали в эксперименте ионизирующее облучение до 570 тыс. рентген в течение 24 ч. Обезвоженные личинки одного из африканских комаров-хирономусов – Polypodium vanderplanki – сохраняют способность оживать после воздействия температуры в +102 °C.
Состояние анабиоза намного расширяет границы сохранения жизни, в том числе и во времени. Например, в толще ледника Антарктиды при глубоком бурении были обнаружены микроорганизмы (споры бактерий, грибов и дрожжей), развившиеся впоследствии на обычных питательных средах. Возраст соответствующих горизонтов льда достигает 10–13 тыс. лет. Споры некоторых жизнеспособных бактерий выделены и из более глубоких слоев возрастом в сотни тысяч лет.
Анабиоз, однако, – достаточно редкое явление. Он возможен далеко не для всех видов и является крайним состоянием покоя в живой природе. Его необходимое условие – сохранение неповрежденными тонких внутриклеточных структур (органелл и мембран) при высушивании или глубоком охлаждении организмов. Это условие невыполнимо для большинства видов, имеющих сложную организацию клеток, тканей и органов.
Способность к анабиозу обнаруживается у видов, имеющих простое или упрощенное строение и обитающих в условиях резкого колебания влажности (пересыхающие мелкие водоемы, верхние слои почвы, подушки мхов и лишайников и т. п.).
Гораздо шире распространены в природе другие формы покоя, связанные с состоянием пониженной жизнедеятельности в результате частичного угнетения метаболизма. Любая степень снижения уровня обмена веществ повышает устойчивость организмов и позволяет более экономно тратить энергию.
Формы покоя в состоянии пониженной жизнедеятельности делят на гипобиоз и криптобиоз, или покой вынужденный и покой физиологический. При гипобиозе торможение активности, или оцепенение, возникает под прямым давлением неблагоприятных условий и прекращается почти сразу после того, как эти условия возвращаются к норме (рис. 9). Подобное подавление процессов жизнедеятельности может возникать при недостатке тепла, воды, кислорода, при повышении осмотического давления и т. п. В соответствии с ведущим внешним фактором вынужденного покоя различают криобиоз (при низких температурах), ангидробиоз (при недостатке воды), аноксибиоз (в анаэробных условиях), гиперосмобиоз (при высоком содержании солей в воде) и др.
He только в арктических и антарктических, но и в средних широтах некоторые морозостойкие виды членистоногих (коллемболы, ряд мух, жужелицы и др.) зимуют в состоянии оцепенения, быстро оттаивая и переходя к активности под лучами солнца, а затем вновь теряют подвижность при снижении температуры. Взошедшие весной растения прекращают и возобновляют рост и развитие вслед за похолоданием и потеплением. После выпавшего дождя голый грунт часто зеленеет за счет быстрого размножения почвенных водорослей, находившихся в вынужденном покое.
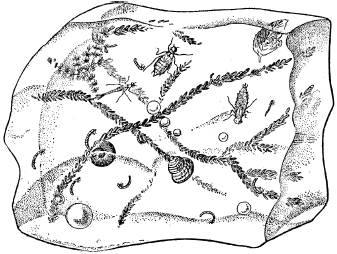
Рис. 9. Пагон – кусок льда со вмерзшими в него пресноводными обитателями (из С. А. Зернова, 1949)
Глубина и продолжительность подавления обмена веществ при гипобиозе зависит от длительности и интенсивности действия угнетающего фактора. Вынужденный покой наступает на любой стадии онтогенеза. Выгоды гипобиоза – быстрое восстановление активной жизнедеятельности. Однако это относительно неустойчивое состояние организмов и при большой длительности может быть повреждающим из-за разбалансированности метаболических процессов, истощения энергетических ресурсов, накопления недоокисленных продуктов обмена и других неблагоприятных физиологических изменений.
Криптобиоз – принципиально другой тип покоя. Он связан с комплексом эндогенных физиологических перестроек, которые происходят заблаговременно, до наступления неблагоприятных сезонных изменений, и организмы оказываются к ним готовы. Криптобиоз является адаптацией прежде всего к сезонной или иной периодичности абиотических факторов внешней среды, их регулярной цикличности. Он составляет часть жизненного цикла организмов, возникает не на любой, а на определенной стадии индивидуального развития, приуроченной к переживанию критических периодов года.
Переход в состояние физиологического покоя требует времени. Ему предшествует накопление резервных веществ, частичная дегидратация тканей и органов, уменьшение интенсивности окислительных процессов и ряд других изменений, понижающих в целом тканевый метаболизм. В состоянии криптобиоза организмы становятся во много раз более устойчивыми к неблагоприятным воздействиям внешней среды (рис. 10). Основные биохимические перестройки при этом являются во многом общими для растений, животных и микроорганизмов (например, переключение метаболизма в разной степени на путь гликолиза за счет резервных углеводов и т. п.). Выход из криптобиоза также требует времени и затрат энергии и не может быть осуществлен простым прекращением отрицательного действия фактора. Для этого необходимы особые условия, различные для разных видов (например, промораживание, присутствие капельно-жидкой воды, определенная продолжительность светового дня, определенное качество света, обязательные колебания температуры и др.).
Криптобиоз как стратегия выживания в периодически неблагоприятных для активной жизни условиях – это продукт длительной эволюции и естественного отбора. Он широко распространен в живой природе. Состояние криптобиоза характерно, например, для семян растений, цист и спор различных микроорганизмов, грибов, водорослей. Диапауза членистоногих, спячка млекопитающих, глубокий покой растений – также различные типы криптобиоза.
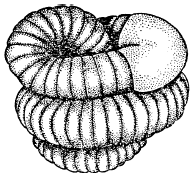
Рис. 10. Дождевой червь в состоянии диапаузы (по В. Тишлеру, 1971)
Состояния гипобиоза, криптобиоза и анабиоза обеспечивают выживание видов в природных условиях разных широт, часто экстремальных, позволяют сохранять организмы в течение длительных неблагоприятных периодов, расселяться в пространстве и во многом раздвигают границы возможности и распространения жизни в целом.
В состоянии глубокого покоя, т. е. пониженного вещественно-энергетического обмена, организмы становятся менее зависимыми от среды, приобретают высокую степень устойчивости и способны переносить условия, которые не могли бы выдержать при активной жизнедеятельности. Эти два состояния чередуются в жизни многих видов, являясь адаптацией к местообитаниям с нестабильным климатом, резкими сезонными изменениями, что характерно для большей части планеты.
При глубоком подавлении обмена веществ организмы могут вообще не проявлять видимых признаков жизни. Вопрос о том, возможна ли полная остановка обмена веществ с последующим возвращением к активной жизнедеятельности, т. е. своего рода «воскрешение из мертвых», дискутировался в науке более двух столетий.
Впервые явление мнимой смерти было обнаружено в 1702 г. Антони ван Левенгуком – открывателем микроскопического мира живых существ. Наблюдаемые им «анималькули» (коловратки) при высыхании капли воды сморщивались, выглядели мертвыми и могли пребывать в таком состоянии длительное время (рис. 8). Помещенные вновь в воду, они набухали и переходили к активной жизни. Левенгук объяснил это явление тем, что оболочка «анималькулей», очевидно, «не позволяет ни малейшего испарения» и они остаются живыми в сухих условиях. Однако через несколько десятилетий естествоиспытатели уже спорили о возможности того, что «жизнь может быть полностью прекращена» и восстановлена вновь «через 20, 40, 100 лет или более».
В 70-х годах XVIII в. явление «воскрешения» после высыхания было обнаружено и подтверждено многочисленными опытами у ряда других мелких организмов – пшеничных угриц, свободноживущих нематод и тихоходок. Ж. Бюффон, повторив опыты Дж. Нидгема с угрицами, утверждал, что «эти организмы можно заставить сколько угодно раз подряд умирать и вновь оживать». Л. Спалланцани впервые обратил внимание на глубокий покой семян и спор растений, расценив его как сохранение их во времени.
Рис. 8. Коловратка Philidina roseola на разных стадиях высыхания (по П. Ю. Шмидту, 1948):
1– активная; 2 – начинающая сокращаться; 3 – полностью сократившаяся перед высыханием; 4 – в состоянии анабиоза
В середине XIX в. было убедительно установлено, что устойчивость сухих коловраток, тихоходок и нематод к высоким и низким температурам, недостатку или отсутствию кислорода возрастает пропорционально степени их обезвоживания. Однако оставался открытым вопрос, происходит ли при этом полное прерывание жизни или лишь ее глубокое угнетение. В 1878 г. Клод Бернал выдвинул понятие «скрытая жизнь», которую он характеризовал прекращением обмена веществ и «перерывом отношений между существом и средой».
Окончательно этот вопрос был решен лишь в первой трети XX столетия с развитием техники глубокого вакуумного обезвоживания. Опыты Г. Рама, П. Беккереля и других ученых показали возможность полной обратимой остановки жизни. В сухом состоянии, когда в клетках оставалось не более 2 % воды в химически связанном виде, такие организмы, как коловратки, тихоходки, мелкие нематоды, семена и споры растений, споры бактерий и грибов выдерживали пребывание в жидком кислороде (-218,4 °C), жидком водороде (-259,4 °C), жидком гелии (-269,0 °C), т. е. температуры, близкие к абсолютному нулю. При этом содержимое клеток затвердевает, отсутствует даже тепловое движение молекул, и всякий обмен веществ, естественно, прекращен. После помещения в нормальные условия эти организмы продолжают развитие. У некоторых видов остановка обмена веществ при сверхнизких температурах возможна и без высушивания, при условии замерзания воды не в кристаллическом, а в аморфном состоянии.
Полная временная остановка жизни получила название анабиоза. Термин был предложен В. Прейером еще в 1891 г. В состоянии анабиоза организмы становятся устойчивыми к самым разнообразным воздействиям. Например, тихоходки выдерживали в эксперименте ионизирующее облучение до 570 тыс. рентген в течение 24 ч. Обезвоженные личинки одного из африканских комаров-хирономусов – Polypodium vanderplanki – сохраняют способность оживать после воздействия температуры в +102 °C.
Состояние анабиоза намного расширяет границы сохранения жизни, в том числе и во времени. Например, в толще ледника Антарктиды при глубоком бурении были обнаружены микроорганизмы (споры бактерий, грибов и дрожжей), развившиеся впоследствии на обычных питательных средах. Возраст соответствующих горизонтов льда достигает 10–13 тыс. лет. Споры некоторых жизнеспособных бактерий выделены и из более глубоких слоев возрастом в сотни тысяч лет.
Анабиоз, однако, – достаточно редкое явление. Он возможен далеко не для всех видов и является крайним состоянием покоя в живой природе. Его необходимое условие – сохранение неповрежденными тонких внутриклеточных структур (органелл и мембран) при высушивании или глубоком охлаждении организмов. Это условие невыполнимо для большинства видов, имеющих сложную организацию клеток, тканей и органов.
Способность к анабиозу обнаруживается у видов, имеющих простое или упрощенное строение и обитающих в условиях резкого колебания влажности (пересыхающие мелкие водоемы, верхние слои почвы, подушки мхов и лишайников и т. п.).
Гораздо шире распространены в природе другие формы покоя, связанные с состоянием пониженной жизнедеятельности в результате частичного угнетения метаболизма. Любая степень снижения уровня обмена веществ повышает устойчивость организмов и позволяет более экономно тратить энергию.
Формы покоя в состоянии пониженной жизнедеятельности делят на гипобиоз и криптобиоз, или покой вынужденный и покой физиологический. При гипобиозе торможение активности, или оцепенение, возникает под прямым давлением неблагоприятных условий и прекращается почти сразу после того, как эти условия возвращаются к норме (рис. 9). Подобное подавление процессов жизнедеятельности может возникать при недостатке тепла, воды, кислорода, при повышении осмотического давления и т. п. В соответствии с ведущим внешним фактором вынужденного покоя различают криобиоз (при низких температурах), ангидробиоз (при недостатке воды), аноксибиоз (в анаэробных условиях), гиперосмобиоз (при высоком содержании солей в воде) и др.
He только в арктических и антарктических, но и в средних широтах некоторые морозостойкие виды членистоногих (коллемболы, ряд мух, жужелицы и др.) зимуют в состоянии оцепенения, быстро оттаивая и переходя к активности под лучами солнца, а затем вновь теряют подвижность при снижении температуры. Взошедшие весной растения прекращают и возобновляют рост и развитие вслед за похолоданием и потеплением. После выпавшего дождя голый грунт часто зеленеет за счет быстрого размножения почвенных водорослей, находившихся в вынужденном покое.
Рис. 9. Пагон – кусок льда со вмерзшими в него пресноводными обитателями (из С. А. Зернова, 1949)
Глубина и продолжительность подавления обмена веществ при гипобиозе зависит от длительности и интенсивности действия угнетающего фактора. Вынужденный покой наступает на любой стадии онтогенеза. Выгоды гипобиоза – быстрое восстановление активной жизнедеятельности. Однако это относительно неустойчивое состояние организмов и при большой длительности может быть повреждающим из-за разбалансированности метаболических процессов, истощения энергетических ресурсов, накопления недоокисленных продуктов обмена и других неблагоприятных физиологических изменений.
Криптобиоз – принципиально другой тип покоя. Он связан с комплексом эндогенных физиологических перестроек, которые происходят заблаговременно, до наступления неблагоприятных сезонных изменений, и организмы оказываются к ним готовы. Криптобиоз является адаптацией прежде всего к сезонной или иной периодичности абиотических факторов внешней среды, их регулярной цикличности. Он составляет часть жизненного цикла организмов, возникает не на любой, а на определенной стадии индивидуального развития, приуроченной к переживанию критических периодов года.
Переход в состояние физиологического покоя требует времени. Ему предшествует накопление резервных веществ, частичная дегидратация тканей и органов, уменьшение интенсивности окислительных процессов и ряд других изменений, понижающих в целом тканевый метаболизм. В состоянии криптобиоза организмы становятся во много раз более устойчивыми к неблагоприятным воздействиям внешней среды (рис. 10). Основные биохимические перестройки при этом являются во многом общими для растений, животных и микроорганизмов (например, переключение метаболизма в разной степени на путь гликолиза за счет резервных углеводов и т. п.). Выход из криптобиоза также требует времени и затрат энергии и не может быть осуществлен простым прекращением отрицательного действия фактора. Для этого необходимы особые условия, различные для разных видов (например, промораживание, присутствие капельно-жидкой воды, определенная продолжительность светового дня, определенное качество света, обязательные колебания температуры и др.).
Криптобиоз как стратегия выживания в периодически неблагоприятных для активной жизни условиях – это продукт длительной эволюции и естественного отбора. Он широко распространен в живой природе. Состояние криптобиоза характерно, например, для семян растений, цист и спор различных микроорганизмов, грибов, водорослей. Диапауза членистоногих, спячка млекопитающих, глубокий покой растений – также различные типы криптобиоза.
Рис. 10. Дождевой червь в состоянии диапаузы (по В. Тишлеру, 1971)
Состояния гипобиоза, криптобиоза и анабиоза обеспечивают выживание видов в природных условиях разных широт, часто экстремальных, позволяют сохранять организмы в течение длительных неблагоприятных периодов, расселяться в пространстве и во многом раздвигают границы возможности и распространения жизни в целом.
Глава 3. ВАЖНЕЙШИЕ АБИОТИЧЕСКИЕ ФАКТОРЫ И АДАПТАЦИИ К НИМ ОРГАНИЗМОВ
3.1. Температура
Температура отражает среднюю кинетическую скорость атомов и молекул в какой-либо системе. От температуры зависит и скорость в организме биохимических реакций, составляющих обмен веществ. Повышение температуры увеличивает количество молекул, обладающих энергией активации. Коэффициент, показывающий, во сколько раз изменяется скорость реакций при изменении температуры на 10 °C, обозначают Q10. Для большинства химических реакций величина этого коэффициента равна 2–3 (закон Вант-Гоффа). Изменения температуры приводят также к изменениям стереохимической специфичности макромолекул: третичной и четвертичной структуры белков, строения нуклеиновых кислот, организации мембран и других структур клетки. Так как величина Q10 для разных биохимических реакций различна, то изменения температуры могут сильно нарушить сбалансированность обмена веществ, если скорости сопряженных процессов изменятся различным образом. Сильное понижение температуры вызывает опасность такого замедления обмена веществ, при котором окажется невозможным осуществление основных жизненных функций организма. Критическим моментом является замерзание воды в клетках, так как появление кристалликов льда несовместимо с сохранением целостности внутриклеточных структур. Повышение температуры ведет к денатурации белков, в среднем в области около 60 °C, но рассогласование биохимических и физиологических процессов начинается раньше, уже при некотором превышении 42–43 °C. Излишнее усиление метаболизма при высоких температурах тела также может вывести организм из строя еще задолго до теплового разрушения ферментов, так как резко возрастают потребности в питательных веществах и кислороде, которые далеко не всегда могут быть удовлетворены. Таким образом, жизнь организмов в среде с низкими, высокими и колеблющимися температурами представляет сложную задачу адаптации, решаемую в ходе эволюции и индивидуального развития.
В процессе эволюции у живых организмов выработались разнообразные приспособления, позволяющие регулировать обмен веществ при изменениях температуры окружающей среды. Это достигается двумя путями: 1) различными биохимическими и физиологическими перестройками (изменение набора, концентрации и активности ферментов, обезвоживание, понижение точки замерзания растворов тела и т. д.); 2) поддержанием температуры тела на более стабильном уровне, чем температура окружающей среды, что позволяет не слишком нарушать сложившийся ход биохимических реакций.
В процессе эволюции у живых организмов выработались разнообразные приспособления, позволяющие регулировать обмен веществ при изменениях температуры окружающей среды. Это достигается двумя путями: 1) различными биохимическими и физиологическими перестройками (изменение набора, концентрации и активности ферментов, обезвоживание, понижение точки замерзания растворов тела и т. д.); 2) поддержанием температуры тела на более стабильном уровне, чем температура окружающей среды, что позволяет не слишком нарушать сложившийся ход биохимических реакций.
3.1.1. Температурные границы существования видов
В среднем активная жизнедеятельность организмов требует довольно узкого диапазона температур, ограниченного критическими порогами замерзания воды и тепловой денатурации белков, примерно в пределах от 0 до +50 °C.Границы оптимальных температур соответственно должны быть еще более узкими. Однако реально эти границы преодолеваются в природе у многих видов за счет специфических адаптаций. Существуют экологические группы организмов, оптимум которых сдвинут в сторону низких или высоких температур.
Криофилы – виды, предпочитающие холод и специализированные к жизни в этих условиях. Свыше 80 % земной биосферы относится к постоянно холодным областям с температурой ниже +5 °C – это глубины Мирового океана, арктические и антарктические пустыни, тундры, высокогорья. Обитающие здесь виды обладают повышенной холодостойкостью. Основные механизмы этих адаптаций биохимические. Ферменты холодолюбивых организмов обладают такими особенностями строения, которые позволяют им эффективно понижать энергию активации молекул и поддерживать клеточный метаболизм при температурах, близких к 0 °C. Большую роль играют также механизмы, предотвращающие образование льда внутри клеток. При этом реализуются два основных пути – противостояние замерзанию (резистентность) и устойчивость к замерзанию (толерантность).
Биохимический путь противостояния замерзанию – накопление в клетках макромолекулярных веществ – антифризов, которые понижают точку замерзания жидкостей тела и препятствуют образованию кристаллов льда в организме. Такого типа холодовые адаптации обнаружены, например, у антарктических рыб семейства нототениевых, которые живут при температуре тела -1,86 °C, плавая под поверхностью сплошного льда в воде с такою же температурой (рис. 11). Мелкая тресковая рыба сайка в Северном Ледовитом океане плавает в водах с температурой не выше +5 °C, а нерестится зимой в переохлажденных водах у побережья. Глубоководные рыбы в приполярных районах все время находятся в переохлажденном состоянии.
Предельная температура, при которой еще возможна активность клеток, зафиксирована у микроорганизмов. В холодильных камерах мясные продукты могут быть испорчены за счет деятельности бактерий при температурах до -10-12 °C. Ниже этих температур роста и развития одноклеточных организмов не происходит.
Рис. 11. Антарктическая рыба трематом-пестряк с температурой тела -1,98 °C (см.: Жизнь животных. Т. 4. 1971)
Другой путь холодостойкости – выносливость к замерзанию – связан с временным прекращением активного состояния (гипобиозом или криптобиозом). Образование кристалликов льда внутри клеток необратимо нарушает их ультраструктуру и приводит к гибели. Но многие криофилы способны переносить образование льда во внеклеточных жидкостях. Этот процесс приводит к частичной дегидратации клеток, что повышает их устойчивость. У насекомых накопление защитных органических веществ, таких как глицерин, сорбит, маннит и других, препятствует кристаллизации внутриклеточных растворов и позволяет переживать критические морозные периоды в состоянии оцепенения. Так, жуки-жужелицы в тундрах выдерживают переохлаждение до -35 °C, накапливая к зиме до 25 % глицерина и снижая содержание воды в теле с 65 до 54 %. Летом глицерин в их теле не обнаруживается. Некоторые насекомые выдерживают зимой до -47 и даже -50 °C с замерзанием внеклеточной, но не внутриклеточной влаги. Морские обитатели практически не сталкиваются с температурами ниже -2 °C, но беспозвоночные приливно-отливной зоны (моллюски, усоногие раки и др.) зимой во время отлива переносят замерзание до – (15–20) °С. Клетки под микроскопом выглядят сморщенными, но кристаллов льда в них не обнаруживается. Устойчивость к замерзанию может проявляться и у эвритермных видов, оптимальные температуры развития которых далеки от 0 °C.
Термофилы – это экологическая группа видов, оптимум жизнедеятельности которых приурочен к области высоких температур. Термофилией отличаются многие представители микроорганизмов, растений и животных, встречающихся в горячих источниках, на поверхности прогреваемых почв, в разлагающихся органических остатках при их саморазогревании и т. п.
Верхние температурные пределы активной жизни отличаются у разных групп организмов. Наиболее устойчивы бактерии. У одного из видов архебактерий, распространенных на глубинах вокруг термальных источников («курильщиков»), экспериментально обнаружена способность к росту и делению клеток при температурах, превышающих +110 °C. Некоторые бактерии, окисляющие серу, как, например, Sulfolobus acidocaldarius, размножаются при +(85–90)°С. Обнаружена даже способность ряда видов расти в практически кипящей воде. Естественно, не все бактерии активны при столь высоких температурах, но разнообразие таких видов достаточно велико.
Верхние температурные пороги развития цианобактерий (сине-зеленых водорослей) и других фотосинтезирующих прокариот лежат в более низких пределах от +70 до +73 °C. Термофилы, растущие при +(60–75) °С, есть как среди аэробных, так и анаэробных бактерий, спорообразующих, молочнокислых, актиномицетов, метанообразующих и др. В неактивном состоянии спорообразующие бактерии выдерживают до +200 °C в течение десятков минут, что демонстрирует режим стерилизации предметов в автоклавах.
Криофилы – виды, предпочитающие холод и специализированные к жизни в этих условиях. Свыше 80 % земной биосферы относится к постоянно холодным областям с температурой ниже +5 °C – это глубины Мирового океана, арктические и антарктические пустыни, тундры, высокогорья. Обитающие здесь виды обладают повышенной холодостойкостью. Основные механизмы этих адаптаций биохимические. Ферменты холодолюбивых организмов обладают такими особенностями строения, которые позволяют им эффективно понижать энергию активации молекул и поддерживать клеточный метаболизм при температурах, близких к 0 °C. Большую роль играют также механизмы, предотвращающие образование льда внутри клеток. При этом реализуются два основных пути – противостояние замерзанию (резистентность) и устойчивость к замерзанию (толерантность).
Биохимический путь противостояния замерзанию – накопление в клетках макромолекулярных веществ – антифризов, которые понижают точку замерзания жидкостей тела и препятствуют образованию кристаллов льда в организме. Такого типа холодовые адаптации обнаружены, например, у антарктических рыб семейства нототениевых, которые живут при температуре тела -1,86 °C, плавая под поверхностью сплошного льда в воде с такою же температурой (рис. 11). Мелкая тресковая рыба сайка в Северном Ледовитом океане плавает в водах с температурой не выше +5 °C, а нерестится зимой в переохлажденных водах у побережья. Глубоководные рыбы в приполярных районах все время находятся в переохлажденном состоянии.
Предельная температура, при которой еще возможна активность клеток, зафиксирована у микроорганизмов. В холодильных камерах мясные продукты могут быть испорчены за счет деятельности бактерий при температурах до -10-12 °C. Ниже этих температур роста и развития одноклеточных организмов не происходит.
Рис. 11. Антарктическая рыба трематом-пестряк с температурой тела -1,98 °C (см.: Жизнь животных. Т. 4. 1971)
Другой путь холодостойкости – выносливость к замерзанию – связан с временным прекращением активного состояния (гипобиозом или криптобиозом). Образование кристалликов льда внутри клеток необратимо нарушает их ультраструктуру и приводит к гибели. Но многие криофилы способны переносить образование льда во внеклеточных жидкостях. Этот процесс приводит к частичной дегидратации клеток, что повышает их устойчивость. У насекомых накопление защитных органических веществ, таких как глицерин, сорбит, маннит и других, препятствует кристаллизации внутриклеточных растворов и позволяет переживать критические морозные периоды в состоянии оцепенения. Так, жуки-жужелицы в тундрах выдерживают переохлаждение до -35 °C, накапливая к зиме до 25 % глицерина и снижая содержание воды в теле с 65 до 54 %. Летом глицерин в их теле не обнаруживается. Некоторые насекомые выдерживают зимой до -47 и даже -50 °C с замерзанием внеклеточной, но не внутриклеточной влаги. Морские обитатели практически не сталкиваются с температурами ниже -2 °C, но беспозвоночные приливно-отливной зоны (моллюски, усоногие раки и др.) зимой во время отлива переносят замерзание до – (15–20) °С. Клетки под микроскопом выглядят сморщенными, но кристаллов льда в них не обнаруживается. Устойчивость к замерзанию может проявляться и у эвритермных видов, оптимальные температуры развития которых далеки от 0 °C.
Термофилы – это экологическая группа видов, оптимум жизнедеятельности которых приурочен к области высоких температур. Термофилией отличаются многие представители микроорганизмов, растений и животных, встречающихся в горячих источниках, на поверхности прогреваемых почв, в разлагающихся органических остатках при их саморазогревании и т. п.
Верхние температурные пределы активной жизни отличаются у разных групп организмов. Наиболее устойчивы бактерии. У одного из видов архебактерий, распространенных на глубинах вокруг термальных источников («курильщиков»), экспериментально обнаружена способность к росту и делению клеток при температурах, превышающих +110 °C. Некоторые бактерии, окисляющие серу, как, например, Sulfolobus acidocaldarius, размножаются при +(85–90)°С. Обнаружена даже способность ряда видов расти в практически кипящей воде. Естественно, не все бактерии активны при столь высоких температурах, но разнообразие таких видов достаточно велико.
Верхние температурные пороги развития цианобактерий (сине-зеленых водорослей) и других фотосинтезирующих прокариот лежат в более низких пределах от +70 до +73 °C. Термофилы, растущие при +(60–75) °С, есть как среди аэробных, так и анаэробных бактерий, спорообразующих, молочнокислых, актиномицетов, метанообразующих и др. В неактивном состоянии спорообразующие бактерии выдерживают до +200 °C в течение десятков минут, что демонстрирует режим стерилизации предметов в автоклавах.