Страница:
Основные способы регуляции температуры тела у пойкилотермных животных – поведенческие: перемена позы, активный поиск благоприятных мест обитания, целый ряд специализированных форм поведения, направленных на создание микроклимата (рытье нор, сооружение гнезд и др.).
Переменой позы животное может усилить или ослабить нагревание за счет солнечной радиации. Например, пустынная саранча в прохладные утренние часы подставляет солнечным лучам широкую боковую поверхность тела, а в полдень – узкую спинную. Ящерицы даже высоко в горах в период нормальной активности могут поддерживать температуру тела, используя нагревание прямыми солнечными лучами и тепло нагретых скал. По исследованиям на Кавказе, на высоте 4100 м температура тела Lacerta agilis временами на 29 °C превышала температуру воздуха, держась на уровне 32–36 °C. В сильную жару животные прячутся в тень, скрываются в норах, щелях и т. п. В пустынях днем, например, некоторые виды ящериц и змей взбираются на кусты или зарываются в менее нагретые слои песка, избегая соприкосновения с раскаленной поверхностью грунта. Ящерицы при необходимости стремительно перебегают горячие поверхности только на задних ногах, уменьшая тем самым контакт с почвой (рис. 16). К зиме многие животные ищут убежища, где ход температур более сглажен по сравнению с открытыми местами обитания. Еще более сложны формы поведения общественных насекомых: пчел, муравьев, термитов, которые строят гнезда с хорошо регулируемой внутри них температурой, почти постоянной в период их активности.
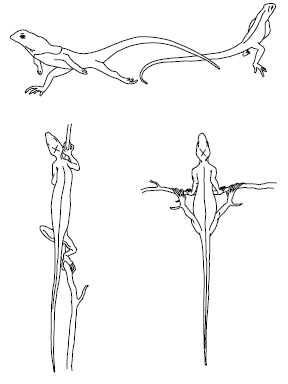
Рис. 16. Поведение ящериц, спасающихся от раскаленной поверхности песка в пустыне
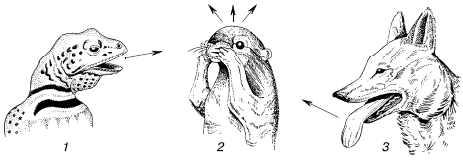
Рис. 17. Испарительная терморегуляция у животных:
1– ящерица – испарение со слизистых при открытом рте;
2– антилоповый суслик – натирание слюной;
3– койот – испарение со слизистых при учащенном дыхании
У ряда пойкилотермных животных эффективно действует и механизм испарительной терморегуляции. Лягушка за час при +20 °C теряет на суше 7770 Дж, что в 300 раз больше ее собственной теплопродукции. Многие рептилии при приближении температуры к верхней критической начинают тяжело дышать или держать рот открытым, усиливая отдачу воды со слизистых оболочек (рис. 17). Пчелы, летающие в жаркую погоду, избегают перегрева, выделяя изо рта каплю жидкости, испарение которой удаляет избыток тепла.
Однако, несмотря на ряд возможностей физической и поведенческой терморегуляции, пойкилотермные животные могут осуществлять ее лишь в узком диапазоне температур. Из-за общего низкого уровня метаболизма они не могут обеспечить постоянство теплового баланса и достаточно активны только вблизи от верхних температурных границ существования. Овладение местообитаниями с постоянно низкими температурами для холоднокровных животных затруднительно. Оно возможно только при развитии специализированной криофилии и в наземных условиях доступно лишь мелким формам, способным использовать малейшие преимущества микроклимата.
3.1.4. Температурные адаптации гомойотермных организмов
3.1.5. Экологические выгоды пойкилотермии и гомойотермии
3.1.6. Сочетание элементов разных стратегий
3.2. Свет
3.2.1. Солнечная радиация
Переменой позы животное может усилить или ослабить нагревание за счет солнечной радиации. Например, пустынная саранча в прохладные утренние часы подставляет солнечным лучам широкую боковую поверхность тела, а в полдень – узкую спинную. Ящерицы даже высоко в горах в период нормальной активности могут поддерживать температуру тела, используя нагревание прямыми солнечными лучами и тепло нагретых скал. По исследованиям на Кавказе, на высоте 4100 м температура тела Lacerta agilis временами на 29 °C превышала температуру воздуха, держась на уровне 32–36 °C. В сильную жару животные прячутся в тень, скрываются в норах, щелях и т. п. В пустынях днем, например, некоторые виды ящериц и змей взбираются на кусты или зарываются в менее нагретые слои песка, избегая соприкосновения с раскаленной поверхностью грунта. Ящерицы при необходимости стремительно перебегают горячие поверхности только на задних ногах, уменьшая тем самым контакт с почвой (рис. 16). К зиме многие животные ищут убежища, где ход температур более сглажен по сравнению с открытыми местами обитания. Еще более сложны формы поведения общественных насекомых: пчел, муравьев, термитов, которые строят гнезда с хорошо регулируемой внутри них температурой, почти постоянной в период их активности.
Рис. 16. Поведение ящериц, спасающихся от раскаленной поверхности песка в пустыне
Рис. 17. Испарительная терморегуляция у животных:
1– ящерица – испарение со слизистых при открытом рте;
2– антилоповый суслик – натирание слюной;
3– койот – испарение со слизистых при учащенном дыхании
У ряда пойкилотермных животных эффективно действует и механизм испарительной терморегуляции. Лягушка за час при +20 °C теряет на суше 7770 Дж, что в 300 раз больше ее собственной теплопродукции. Многие рептилии при приближении температуры к верхней критической начинают тяжело дышать или держать рот открытым, усиливая отдачу воды со слизистых оболочек (рис. 17). Пчелы, летающие в жаркую погоду, избегают перегрева, выделяя изо рта каплю жидкости, испарение которой удаляет избыток тепла.
Однако, несмотря на ряд возможностей физической и поведенческой терморегуляции, пойкилотермные животные могут осуществлять ее лишь в узком диапазоне температур. Из-за общего низкого уровня метаболизма они не могут обеспечить постоянство теплового баланса и достаточно активны только вблизи от верхних температурных границ существования. Овладение местообитаниями с постоянно низкими температурами для холоднокровных животных затруднительно. Оно возможно только при развитии специализированной криофилии и в наземных условиях доступно лишь мелким формам, способным использовать малейшие преимущества микроклимата.
3.1.4. Температурные адаптации гомойотермных организмов
Гомойотермия – принципиально иной путь температурных адаптаций, возникший на основе резкого повышения уровня окислительных процессов у птиц и млекопитающих в результате эволюционного совершенствования кровеносной, дыхательной и других систем органов. Потребление кислорода на 1 г массы тела у теплокровных животных в десятки и сотни раз больше, чем у пойкилотермных.
Основные отличия гомойотермных животных от пойкилотермных организмов: 1) мощный поток внутреннего, эндогенного тепла; 2) развитие целостной системы эффективно работающих терморегуляторных механизмов, и в результате 3) постоянное протекание всех физиологических процессов в оптимальном температурном режиме.
Гомойотермные сохраняют постоянный тепловой баланс между теплопродукцией и теплоотдачей и соответственно поддерживают постоянную высокую температуру тела. Организм теплокровного животного не может быть временно «приостановлен» так, как это происходит при гипобиозе или криптобиозе у пойкилотермных.
Гомойотермные животные всегда вырабатывают определенный минимум теплопродукции, обеспечивающий работу кровеносной системы, органов дыхания, выделения и других, даже находясь в покое. Этот минимум получил название базального метаболизма. Переход к активности усиливает выработку тепла и соответственно требует усиления теплоотдачи.
Теплокровным свойственна химическая терморегуляция – рефлекторное увеличение теплопродукции в ответ на понижение температуры среды. Химическая терморегуляция полностью отсутствует у пойкилотермных, у которых, в случае выделения дополнительного тепла, оно генерируется за счет непосредственной двигательной активности животных.
В противоположность пойкилотермным при действии холода в организме теплокровных животных окислительные процессы не ослабевают, а усиливаются, особенно в скелетных мышцах. У многих животных сначала наблюдается мышечная дрожь – несогласованное сокращение мышц, приводящее к выделению тепловой энергии. Кроме того, клетки мышечной и многих других тканей выделяют тепло и без осуществления рабочих функций, приходя в состояние особого терморегуляционного тонуса. При дальнейшем снижении температуры среды тепловой эффект терморегуляционного тонуса возрастает.
При продуцировании дополнительного тепла особенно усиливается обмен липидов, так как нейтральные жиры содержат основной запас химической энергии. Поэтому жировые запасы животных обеспечивают лучшую терморегуляцию. Млекопитающие обладают даже специализированной бурой жировой тканью, в которой вся освобождающаяся химическая энергия, вместо того чтобы переходить в связи ЛТФ, рассеивается в виде тепла, т. е. идет на обогревание организма. Бурая жировая ткань наиболее развита у животных – обитателей холодного климата.
Поддержание температуры за счет возрастания теплопродукции требует большого расхода энергии, поэтому животные при усилении химической терморегуляции либо нуждаются в большом количестве пищи, либо тратят много жировых запасов, накопленных ранее. Например, землеройка бурозубка крошечная имеет исключительно высокий уровень обмена. Чередуя очень короткие периоды сна и активности, она деятельна в любые часы суток и в день съедает корма в 4 раза больше собственной массы. Частота сердцебиения у бурозубок до 1000 в мин. Также и птицам, остающимся на зиму, нужно много корма: им страшны не столько морозы, сколько бескормица. Так, при хорошем урожае семян ели и сосны клесты зимой даже выводят птенцов.
Усиление химической терморегуляции, таким образом, имеет свои пределы, обусловленные возможностью добывания пищи. При недостатке корма зимой такой путь терморегуляции экологически невыгоден. Он, например, слабо развит у всех животных, обитающих за полярным кругом: песцов, моржей, тюленей, белых медведей, северных оленей и др. Для обитателей тропиков химическая терморегуляция также мало характерна, поскольку у них практически не возникает необходимости в дополнительном продуцировании тепла.
В пределах некоторого диапазона внешних температур гомойотермные поддерживают температуру тела, не тратя на это дополнительной энергии, а используя эффективные механизмы физической терморегуляции, позволяющие лучше сохранять или отводить тепло базального метаболизма. Этот диапазон температур, в пределах которого животные чувствуют себя наиболее комфортно, называется термонейтральной зоной. За нижним порогом этой зоны начинается химическая терморегуляция, за верхним – траты энергии на испарение.
Физическая терморегуляция экологически выгодна, так как адаптация к холоду осуществляется не за счет дополнительной выработки тепла, а за счет сохранения его в теле животного. Кроме того, возможна защита от перегрева путем усиления теплоотдачи во внешнюю среду.
Способы физической терморегуляции множественны. В филогенетическом ряду млекопитающих – от насекомоядных к рукокрылым, грызунам и хищникам механизмы физической терморегуляции становятся все более совершенными и разнообразными. К ним следует отнести рефлекторное сужение и расширение кровеносных сосудов кожи, меняющее ее теплопроводность, изменение теплоизолирующих свойств меха и перьевого покрова, противоточный теплообмен путем контакта сосудов при кровоснабжении отдельных органов, регуляцию испарительной теплоотдачи.
Густой мех млекопитающих, перьевой и особенно пуховой покров птиц позволяют сохранять вокруг тела прослойку воздуха с температурой, близкой к температуре тела животного, и тем самым уменьшить теплоизлучение во внешнюю среду. Теплоотдача регулируется наклоном волос и перьев, сезонной сменой меха и оперения. Исключительно теплый зимний мех млекопитающих Заполярья позволяет им в холода обходиться без существенного повышения обмена веществ и снижает потребность в пище. Например, песцы на побережье Северного Ледовитого океана зимой потребляют пищи даже меньше, чем летом.
У морских млекопитающих – ластоногих и китов – слой подкожной жировой клетчатки распределен по всему телу. Толщина подкожного жира у отдельных видов тюленей достигает 7–9 см, а общая его масса составляет до 40–50 % от массы тела. Теплоизолирующий эффект такого «жирового чулка» настолько высок, что под тюленями, часами лежащими на снегу, снег не тает, хотя температура тела животного поддерживается на уровне 38 °C. У животных жаркого климата подобное распределение жировых запасов приводило бы к гибели от перегрева из-за невозможности выведения избытка тепла, поэтому жир у них запасается локально, в отдельных частях тела, не мешая теплоизлучению с общей поверхности (верблюды, курдючные овцы, зебу и др.).
Системы противоточного теплообмена, помогающие поддерживать постоянную температуру внутренних органов, обнаружены в лапах и хвостах у сумчатых, ленивцев, муравьедов, полуобезьян, ластоногих, китов, пингвинов, журавлей и др. При этом сосуды, по которым нагретая кровь движется от центра тела, тесно контактируют со стенками сосудов, направляющих охлажденную кровь от периферии к центру, и отдают им свое тепло.
Немаловажное значение для поддержания температурного баланса имеет отношение поверхности тела к его объему, так как в конечном счете масштабы продуцирования тепла зависят от массы животного, а теплообмен идет через его покровы.
Связь размеров и пропорций тела животных с климатическими условиями их обитания была подмечена еще в XIX в. Согласно правилу Бергмана (1848), если два близких вида теплокровных животных отличаются размерами, то более крупный обитает в более холодном, а более мелкий – в теплом климате. Бергман подчеркивал, что эта закономерность проявляется лишь в том случае, если виды не отличаются другими приспособлениями к терморегуляции. Из проанализированных им 75 групп птиц в фауне Германии около трети удовлетворяло найденному правилу. Д. Лллен в 1877 г. подметил, что у многих млекопитающих и птиц северного полушария относительные размеры конечностей и различных выступающих частей тела (хвостов, ушей, клювов) увеличиваются к югу – правило Аллена. Терморегуляционное значение отдельных участков тела далеко не равноценно. Выступающие части имеют большую относительную поверхность, которая выгодна в условиях жаркого климата. У ряда млекопитающих, например, особое значение для поддержания теплового баланса имеют уши, снабженные, как правило, большим количеством кровеносных сосудов (рис. 18). Огромные уши африканского слона, маленькой пустынной лисички-фенека, американского зайца превратились в специализированные органы терморегуляции. При адаптации к холоду проявляется закон экономии поверхности, так как компактная форма тела с минимальным отношением площади к объему наиболее выгодна для сохранения тепла.

Рис. 18. Африканский длинноухий тушканчик (по В. Е. Соколову и др., 1977)
Если температура среды превышает верхнюю границу термонейтральной зоны, животным приходится затрачивать дополнительную энергию на испарительную терморегуляцию.
Эффективным механизмом отдачи тепла служит испарение воды путем потоотделения или через влажные слизистые оболочки полости рта и верхних дыхательных путей. Способность к образованию пота у разных видов очень различна. Человек при сильной жаре может выделить до 12 л пота в день, рассеяв при этом тепла в десять раз больше по сравнению с нормой. Выделяемая вода, естественно, должна возмещаться через питье. У некоторых животных испарение идет только через слизистые. У собаки, для которой одышка – единственный способ испарительной терморегуляции, частота дыхания при этом доходит до 300–400 вдохов в минуту. Регуляция температуры через испарение требует траты организмом воды и поэтому возможна не во всех условиях существования.
Поведенческие способы регуляции теплообмена для теплокровных животных не менее важны, чем для пойкилотермных, и также чрезвычайно разнообразны – от изменения позы и поисков укрытий до сооружения сложных нор, гнезд, осуществления ближних и дальних миграций.
В норах роющих животных ход температур сглажен тем сильнее, чем больше глубина норы. В средних широтах на расстоянии 150 см от поверхности почвы перестают ощущаться даже сезонные колебания температуры. В особенно искусно построенных гнездах также поддерживается ровный, благоприятный микроклимат (рис. 19). В войлокообразном гнезде синицы-ремеза, имеющем лишь один узкий боковой вход, тепло и сухо в любую погоду.
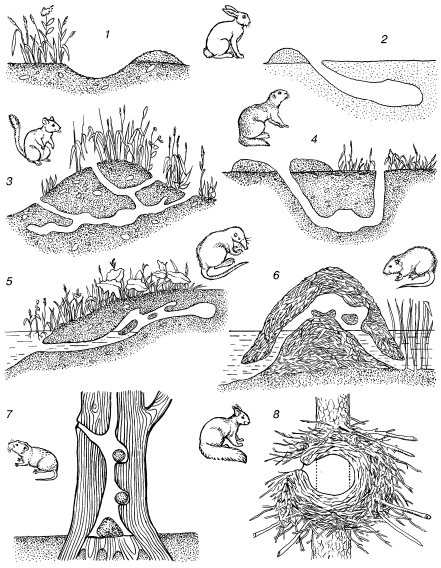
Рис. 19. Особенности строения нор и расположения гнезд разных видов млекопитающих (по Н. П. Наумову, 1963, И. И. Барабаш-Никифорову, Л. Н. Формозову, 1963):
1– логово зайца-русака в песчаных дюнах; 2 – снежная нора зайца-русака; 3 – летняя нора полуденной песчанки; 4 – нора малого суслика; 5 – нора выхухоли; 6 – хатка ондатры; 7 – гнезда рыжей полевки в дупле дуба; 8 – зимнее гнездо обыкновенной белки
В ряде случаев гомойотермные животные используют в целях терморегуляции групповое поведение. Например, некоторые пингвины в сильный мороз и бураны сбиваются в плотную кучу, так называемую черепаху. Особи, оказавшиеся с краю, через некоторое время пробиваются внутрь, и «черепаха» медленно кружится и перемещается. Внутри такого скопления температура поддерживается около +37 °C даже в самые сильные морозы. Обитатели пустынь верблюды в жару также сбиваются вместе, прижимаясь друг к другу боками, но этим достигается противоположный эффект – предотвращение сильного нагревания всей поверхности тела солнечными лучами. Температура в центре скопления животных равна температуре их тела, 39 °C, тогда как шерсть на спине и боках крайних животных нагревается до 70 °C.
Сочетание эффективных способов химической, физической и поведенческой терморегуляции при общем высоком уровне окислительных процессов в организме позволяет гомойотермным животным поддерживать свой тепловой баланс на фоне широких колебаний внешней температуры.
Основные отличия гомойотермных животных от пойкилотермных организмов: 1) мощный поток внутреннего, эндогенного тепла; 2) развитие целостной системы эффективно работающих терморегуляторных механизмов, и в результате 3) постоянное протекание всех физиологических процессов в оптимальном температурном режиме.
Гомойотермные сохраняют постоянный тепловой баланс между теплопродукцией и теплоотдачей и соответственно поддерживают постоянную высокую температуру тела. Организм теплокровного животного не может быть временно «приостановлен» так, как это происходит при гипобиозе или криптобиозе у пойкилотермных.
Гомойотермные животные всегда вырабатывают определенный минимум теплопродукции, обеспечивающий работу кровеносной системы, органов дыхания, выделения и других, даже находясь в покое. Этот минимум получил название базального метаболизма. Переход к активности усиливает выработку тепла и соответственно требует усиления теплоотдачи.
Теплокровным свойственна химическая терморегуляция – рефлекторное увеличение теплопродукции в ответ на понижение температуры среды. Химическая терморегуляция полностью отсутствует у пойкилотермных, у которых, в случае выделения дополнительного тепла, оно генерируется за счет непосредственной двигательной активности животных.
В противоположность пойкилотермным при действии холода в организме теплокровных животных окислительные процессы не ослабевают, а усиливаются, особенно в скелетных мышцах. У многих животных сначала наблюдается мышечная дрожь – несогласованное сокращение мышц, приводящее к выделению тепловой энергии. Кроме того, клетки мышечной и многих других тканей выделяют тепло и без осуществления рабочих функций, приходя в состояние особого терморегуляционного тонуса. При дальнейшем снижении температуры среды тепловой эффект терморегуляционного тонуса возрастает.
При продуцировании дополнительного тепла особенно усиливается обмен липидов, так как нейтральные жиры содержат основной запас химической энергии. Поэтому жировые запасы животных обеспечивают лучшую терморегуляцию. Млекопитающие обладают даже специализированной бурой жировой тканью, в которой вся освобождающаяся химическая энергия, вместо того чтобы переходить в связи ЛТФ, рассеивается в виде тепла, т. е. идет на обогревание организма. Бурая жировая ткань наиболее развита у животных – обитателей холодного климата.
Поддержание температуры за счет возрастания теплопродукции требует большого расхода энергии, поэтому животные при усилении химической терморегуляции либо нуждаются в большом количестве пищи, либо тратят много жировых запасов, накопленных ранее. Например, землеройка бурозубка крошечная имеет исключительно высокий уровень обмена. Чередуя очень короткие периоды сна и активности, она деятельна в любые часы суток и в день съедает корма в 4 раза больше собственной массы. Частота сердцебиения у бурозубок до 1000 в мин. Также и птицам, остающимся на зиму, нужно много корма: им страшны не столько морозы, сколько бескормица. Так, при хорошем урожае семян ели и сосны клесты зимой даже выводят птенцов.
Усиление химической терморегуляции, таким образом, имеет свои пределы, обусловленные возможностью добывания пищи. При недостатке корма зимой такой путь терморегуляции экологически невыгоден. Он, например, слабо развит у всех животных, обитающих за полярным кругом: песцов, моржей, тюленей, белых медведей, северных оленей и др. Для обитателей тропиков химическая терморегуляция также мало характерна, поскольку у них практически не возникает необходимости в дополнительном продуцировании тепла.
В пределах некоторого диапазона внешних температур гомойотермные поддерживают температуру тела, не тратя на это дополнительной энергии, а используя эффективные механизмы физической терморегуляции, позволяющие лучше сохранять или отводить тепло базального метаболизма. Этот диапазон температур, в пределах которого животные чувствуют себя наиболее комфортно, называется термонейтральной зоной. За нижним порогом этой зоны начинается химическая терморегуляция, за верхним – траты энергии на испарение.
Физическая терморегуляция экологически выгодна, так как адаптация к холоду осуществляется не за счет дополнительной выработки тепла, а за счет сохранения его в теле животного. Кроме того, возможна защита от перегрева путем усиления теплоотдачи во внешнюю среду.
Способы физической терморегуляции множественны. В филогенетическом ряду млекопитающих – от насекомоядных к рукокрылым, грызунам и хищникам механизмы физической терморегуляции становятся все более совершенными и разнообразными. К ним следует отнести рефлекторное сужение и расширение кровеносных сосудов кожи, меняющее ее теплопроводность, изменение теплоизолирующих свойств меха и перьевого покрова, противоточный теплообмен путем контакта сосудов при кровоснабжении отдельных органов, регуляцию испарительной теплоотдачи.
Густой мех млекопитающих, перьевой и особенно пуховой покров птиц позволяют сохранять вокруг тела прослойку воздуха с температурой, близкой к температуре тела животного, и тем самым уменьшить теплоизлучение во внешнюю среду. Теплоотдача регулируется наклоном волос и перьев, сезонной сменой меха и оперения. Исключительно теплый зимний мех млекопитающих Заполярья позволяет им в холода обходиться без существенного повышения обмена веществ и снижает потребность в пище. Например, песцы на побережье Северного Ледовитого океана зимой потребляют пищи даже меньше, чем летом.
У морских млекопитающих – ластоногих и китов – слой подкожной жировой клетчатки распределен по всему телу. Толщина подкожного жира у отдельных видов тюленей достигает 7–9 см, а общая его масса составляет до 40–50 % от массы тела. Теплоизолирующий эффект такого «жирового чулка» настолько высок, что под тюленями, часами лежащими на снегу, снег не тает, хотя температура тела животного поддерживается на уровне 38 °C. У животных жаркого климата подобное распределение жировых запасов приводило бы к гибели от перегрева из-за невозможности выведения избытка тепла, поэтому жир у них запасается локально, в отдельных частях тела, не мешая теплоизлучению с общей поверхности (верблюды, курдючные овцы, зебу и др.).
Системы противоточного теплообмена, помогающие поддерживать постоянную температуру внутренних органов, обнаружены в лапах и хвостах у сумчатых, ленивцев, муравьедов, полуобезьян, ластоногих, китов, пингвинов, журавлей и др. При этом сосуды, по которым нагретая кровь движется от центра тела, тесно контактируют со стенками сосудов, направляющих охлажденную кровь от периферии к центру, и отдают им свое тепло.
Немаловажное значение для поддержания температурного баланса имеет отношение поверхности тела к его объему, так как в конечном счете масштабы продуцирования тепла зависят от массы животного, а теплообмен идет через его покровы.
Связь размеров и пропорций тела животных с климатическими условиями их обитания была подмечена еще в XIX в. Согласно правилу Бергмана (1848), если два близких вида теплокровных животных отличаются размерами, то более крупный обитает в более холодном, а более мелкий – в теплом климате. Бергман подчеркивал, что эта закономерность проявляется лишь в том случае, если виды не отличаются другими приспособлениями к терморегуляции. Из проанализированных им 75 групп птиц в фауне Германии около трети удовлетворяло найденному правилу. Д. Лллен в 1877 г. подметил, что у многих млекопитающих и птиц северного полушария относительные размеры конечностей и различных выступающих частей тела (хвостов, ушей, клювов) увеличиваются к югу – правило Аллена. Терморегуляционное значение отдельных участков тела далеко не равноценно. Выступающие части имеют большую относительную поверхность, которая выгодна в условиях жаркого климата. У ряда млекопитающих, например, особое значение для поддержания теплового баланса имеют уши, снабженные, как правило, большим количеством кровеносных сосудов (рис. 18). Огромные уши африканского слона, маленькой пустынной лисички-фенека, американского зайца превратились в специализированные органы терморегуляции. При адаптации к холоду проявляется закон экономии поверхности, так как компактная форма тела с минимальным отношением площади к объему наиболее выгодна для сохранения тепла.
Рис. 18. Африканский длинноухий тушканчик (по В. Е. Соколову и др., 1977)
Если температура среды превышает верхнюю границу термонейтральной зоны, животным приходится затрачивать дополнительную энергию на испарительную терморегуляцию.
Эффективным механизмом отдачи тепла служит испарение воды путем потоотделения или через влажные слизистые оболочки полости рта и верхних дыхательных путей. Способность к образованию пота у разных видов очень различна. Человек при сильной жаре может выделить до 12 л пота в день, рассеяв при этом тепла в десять раз больше по сравнению с нормой. Выделяемая вода, естественно, должна возмещаться через питье. У некоторых животных испарение идет только через слизистые. У собаки, для которой одышка – единственный способ испарительной терморегуляции, частота дыхания при этом доходит до 300–400 вдохов в минуту. Регуляция температуры через испарение требует траты организмом воды и поэтому возможна не во всех условиях существования.
Поведенческие способы регуляции теплообмена для теплокровных животных не менее важны, чем для пойкилотермных, и также чрезвычайно разнообразны – от изменения позы и поисков укрытий до сооружения сложных нор, гнезд, осуществления ближних и дальних миграций.
В норах роющих животных ход температур сглажен тем сильнее, чем больше глубина норы. В средних широтах на расстоянии 150 см от поверхности почвы перестают ощущаться даже сезонные колебания температуры. В особенно искусно построенных гнездах также поддерживается ровный, благоприятный микроклимат (рис. 19). В войлокообразном гнезде синицы-ремеза, имеющем лишь один узкий боковой вход, тепло и сухо в любую погоду.
Рис. 19. Особенности строения нор и расположения гнезд разных видов млекопитающих (по Н. П. Наумову, 1963, И. И. Барабаш-Никифорову, Л. Н. Формозову, 1963):
1– логово зайца-русака в песчаных дюнах; 2 – снежная нора зайца-русака; 3 – летняя нора полуденной песчанки; 4 – нора малого суслика; 5 – нора выхухоли; 6 – хатка ондатры; 7 – гнезда рыжей полевки в дупле дуба; 8 – зимнее гнездо обыкновенной белки
В ряде случаев гомойотермные животные используют в целях терморегуляции групповое поведение. Например, некоторые пингвины в сильный мороз и бураны сбиваются в плотную кучу, так называемую черепаху. Особи, оказавшиеся с краю, через некоторое время пробиваются внутрь, и «черепаха» медленно кружится и перемещается. Внутри такого скопления температура поддерживается около +37 °C даже в самые сильные морозы. Обитатели пустынь верблюды в жару также сбиваются вместе, прижимаясь друг к другу боками, но этим достигается противоположный эффект – предотвращение сильного нагревания всей поверхности тела солнечными лучами. Температура в центре скопления животных равна температуре их тела, 39 °C, тогда как шерсть на спине и боках крайних животных нагревается до 70 °C.
Сочетание эффективных способов химической, физической и поведенческой терморегуляции при общем высоком уровне окислительных процессов в организме позволяет гомойотермным животным поддерживать свой тепловой баланс на фоне широких колебаний внешней температуры.
3.1.5. Экологические выгоды пойкилотермии и гомойотермии
Пойкилотермия и гомойотермия – это, по сути дела, две альтернативные природные стратегии выживания организмов в условиях колеблющихся температур. Каждая из них имеет свои преимущества и недостатки.
Пойкилотермность – это подчинение организмов ходу внешних температур. Производя сравнительно мало эндогенного тепла и обладая лишь отдельными терморегуляторными реакциями, пойкилотермные организмы не могут обеспечить постоянства теплообмена, поэтому при колебаниях температуры среды их активность прерывиста. Переход в неактивное состояние связан с развитием механизмов толерантности к изменениям температуры тела.
Подчинение температуре среды имеет, однако, ряд преимуществ. Снижение уровня обмена при действии холода экономит энергетические затраты, резко уменьшает потребность в пище. В условиях сухого жаркого климата пойкилотермность позволяет избегать излишних потерь воды, так как практическое отсутствие различий между температурами тела и среды не вызывает дополнительного испарения. Высокие температуры пойкилотермные животные переносят легче и с меньшими энергетическими затратами, чем гомойотермные, которые тратят много энергии на удаление избытка тепла из тела.
Гомойотермность – это стратегия сопротивления влиянию факторов среды. Организм гомойотермного животного всегда функционирует только в узких температурных границах. За этими пределами для них невозможно не только сохранение биологической активности, но и переживание в угнетенном состоянии, так как выносливость к значительным колебаниям температуры тела ими потеряна. Зато, отличаясь высокой интенсивностью окислительных процессов в организме и обладая мощным комплексом терморегуляторных средств, гомойотермные животные могут поддерживать для себя постоянный температурный оптимум при значительных отклонениях внешних температур, что позволяет им шире осваивать внешние условия.
Работа механизмов терморегуляции требует больших энергетических затрат, для восполнения которых животные нуждаются в усиленном питании, поэтому единственно возможным состоянием животных с регулируемой температурой тела является состояние постоянной активности. В холодных районах ограничивающим фактором в их распространении является не температура, а возможность регулярного добывания пищи.
Пойкилотермность – это подчинение организмов ходу внешних температур. Производя сравнительно мало эндогенного тепла и обладая лишь отдельными терморегуляторными реакциями, пойкилотермные организмы не могут обеспечить постоянства теплообмена, поэтому при колебаниях температуры среды их активность прерывиста. Переход в неактивное состояние связан с развитием механизмов толерантности к изменениям температуры тела.
Подчинение температуре среды имеет, однако, ряд преимуществ. Снижение уровня обмена при действии холода экономит энергетические затраты, резко уменьшает потребность в пище. В условиях сухого жаркого климата пойкилотермность позволяет избегать излишних потерь воды, так как практическое отсутствие различий между температурами тела и среды не вызывает дополнительного испарения. Высокие температуры пойкилотермные животные переносят легче и с меньшими энергетическими затратами, чем гомойотермные, которые тратят много энергии на удаление избытка тепла из тела.
Гомойотермность – это стратегия сопротивления влиянию факторов среды. Организм гомойотермного животного всегда функционирует только в узких температурных границах. За этими пределами для них невозможно не только сохранение биологической активности, но и переживание в угнетенном состоянии, так как выносливость к значительным колебаниям температуры тела ими потеряна. Зато, отличаясь высокой интенсивностью окислительных процессов в организме и обладая мощным комплексом терморегуляторных средств, гомойотермные животные могут поддерживать для себя постоянный температурный оптимум при значительных отклонениях внешних температур, что позволяет им шире осваивать внешние условия.
Работа механизмов терморегуляции требует больших энергетических затрат, для восполнения которых животные нуждаются в усиленном питании, поэтому единственно возможным состоянием животных с регулируемой температурой тела является состояние постоянной активности. В холодных районах ограничивающим фактором в их распространении является не температура, а возможность регулярного добывания пищи.
3.1.6. Сочетание элементов разных стратегий
Ряд животных способны к сочетанию преимуществ обеих стратегий теплообмена. У некоторых видов гомойотермных, осваивающих экстремальные условия на пределе возможности поддержания теплового баланса, проявляются элементы пойкилотермии, позволяющие им экономить энергию.
У верблюдов в жаркое летнее время дневные колебания температуры тела достигают 6–7 °C. Утром животные имеют температуру 34–35 °C, а затем постепенно нагреваются под палящими лучами солнца, т. е. ведут себя как пойкилотермные организмы. Испарительная терморегуляция через потовые железы и слизистые начинается у верблюда лишь при достижении температуры тела в 40,7 °C, что близко к пределу тепловой выносливости. Вечером, когда температура воздуха падает ниже этой величины, верблюд освобождается от накопившегося избыточного тепла путем прямого излучения. Эти адаптации позволяют животному экономить до 5 л воды, которые потребовались бы для поддержания его средней температуры в пределах 37–38 °C.
Противоположный пример ежесуточного остывания на 5–6 °C в условиях суровых зим на северо-востоке России дают птицы кедровки (рис. 20). За короткий световой день птицы не успевают собрать достаточно пищи, чтобы противостоять низким (иногда до -40 °C) ночным температурам. К утру температура их тела снижается до 34–35 °C, что позволяет выжить, экономя энергию.

Рис. 20. Птицы северных широт, не улетающие на юг: 1– клест; 2 – кедровка; 3 – щур; 4 – глухарь

Рис. 21. Суслик (1) и соня-полчок (2) во время зимней спячки
Гетеротермностъ – особая адаптивная стратегия среди птиц и млекопитающих, при которой закономерно сочетается использование выгод как постоянства, так и перемены температуры тела. Основные формы проявления гетеротермности – способность впадать в спячку или торпидное состояние (оцепенение).
Зимняя спячка характерна для ряда млекопитающих, распространена у однопроходных, насекомоядных, грызунов и летучих мышей. Впадая в спячку, животные прекращают борьбу за поддержание высокой температуры тела, снижая ее всего до нескольких градусов выше нуля. Это сберегает энергетические ресурсы организма. Животные проводят в спячке часто долгие месяцы, иногда до полугода при резко пониженном уровне обмена веществ, постепенно расходуя накопленные резервы (рис. 21). У европейского суслика, например, частота сердцебиений падает с 400 в мин всего до 4–7, у хомяка обыкновенного – с 200 до 12–15, у сони-полчка – с 450 до 35. Потребление кислорода уменьшается в 20-100 раз (рис. 22). Однако состояние спячки не соответствует оцепенению пойкилотермных. Спячка – это хорошо регулируемое физиологическое состояние, при котором терморегуляторная система организма переключается на более низкий уровень. Когда температура в укрытии приближается к 0 °C, животные (хомяки, сурки, сони) начинают генерировать дополнительное тепло, поддерживая температуру тела на уровне не менее 2–3 °C За время спячки крылатые кожаны теряют 30–35 % веса, а суслики – до 50 %. Некоторые виды в зимний период неоднократно пробуждаются и подкармливаются запасами. Суслики до 7 % времени находятся в период спячки в бодрствующем состоянии. Пробуждение млекопитающих происходит путем самосогревания и требует затрат энергии. Возрастает частота сокращений сердца, учащается ритм дыхания. Для выхода из спячки сусликам требуется около 3 часов. Летучие мыши согреваются быстрее и через 15–30 минут уже готовы к полету.
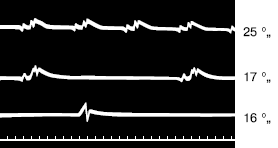
Рис. 22. Изменение числа сердцебиений у сурка вовремя спячки при температуре тела (сверху вниз) 25, 17 и 16 °C. Одно деление шкалы равно 0,2 с (по Эндрес и др., 1930)
Торпидное состояние, или оцепенелость, сопровождает зимнюю спячку животных, но возникает и в других условиях как самостоятельная адаптация. Ряд мелких видов птиц и млекопитающих понижает уровень обмена и температуру тела при неблагоприятных погодных изменениях или регулярно в суточных циклах. У колибри, например, с наступлением ночи температура тела падает с 36–40 °C до 18 °C, птицы садятся на ветки и оцепеневают. Летучие мыши, наоборот, активны в сумеречное и ночное время и впадают в торпидное состояние днем. Переход к оцепенению стимулируется также недостатком пищи. При одних и тех же температурных условиях обеспеченные кормом животные могут продолжать активный образ жизни и генерировать тепло на терморегуляцию, а при нехватке пищи оцепеневают. Кратковременное торпидное состояние, как и спячка, – также регулируемый процесс, но температура тела при этом не падает столь низко и обычно остается на уровне, близком к 20 °C.
Между кратковременным оцепенением гетеротермных животных и длительной зимней спячкой существует вся гамма переходов. Зимний сон медведей, например, не называют спячкой, так как температура этих зверей понижается всего на 3–6 °C, а снижение уровня метаболизма не столь значительно. Мышечная активность обычно подавлена, но большинство физиологических функций сохраняется на прежнем уровне. Роды и выкармливание медвежат молоком у медведиц, например, приходятся на время зимнего сна. Медведи способны относительно легко пробуждаться и переходить в активное состояние.
В пустынях ряд мелких млекопитающих (мышевидные грызуны, сони, некоторые суслики, насекомоядные тенреки и др.) впадают не только в зимнюю, но и в летнюю спячку, которая называется эстивацией (от лат. aestes– лето). Температура их тела падает при этом не столь значительно – до 25–27 °C, но соответственное понижение уровня обмена веществ позволяет экономить не только энергию, но и воду.
Таким образом, широкое распространение гетеротермности позволяет предполагать фундаментальное значение этого явления для выживания животных в неблагоприятных условия.
У верблюдов в жаркое летнее время дневные колебания температуры тела достигают 6–7 °C. Утром животные имеют температуру 34–35 °C, а затем постепенно нагреваются под палящими лучами солнца, т. е. ведут себя как пойкилотермные организмы. Испарительная терморегуляция через потовые железы и слизистые начинается у верблюда лишь при достижении температуры тела в 40,7 °C, что близко к пределу тепловой выносливости. Вечером, когда температура воздуха падает ниже этой величины, верблюд освобождается от накопившегося избыточного тепла путем прямого излучения. Эти адаптации позволяют животному экономить до 5 л воды, которые потребовались бы для поддержания его средней температуры в пределах 37–38 °C.
Противоположный пример ежесуточного остывания на 5–6 °C в условиях суровых зим на северо-востоке России дают птицы кедровки (рис. 20). За короткий световой день птицы не успевают собрать достаточно пищи, чтобы противостоять низким (иногда до -40 °C) ночным температурам. К утру температура их тела снижается до 34–35 °C, что позволяет выжить, экономя энергию.
Рис. 20. Птицы северных широт, не улетающие на юг: 1– клест; 2 – кедровка; 3 – щур; 4 – глухарь
Рис. 21. Суслик (1) и соня-полчок (2) во время зимней спячки
Гетеротермностъ – особая адаптивная стратегия среди птиц и млекопитающих, при которой закономерно сочетается использование выгод как постоянства, так и перемены температуры тела. Основные формы проявления гетеротермности – способность впадать в спячку или торпидное состояние (оцепенение).
Зимняя спячка характерна для ряда млекопитающих, распространена у однопроходных, насекомоядных, грызунов и летучих мышей. Впадая в спячку, животные прекращают борьбу за поддержание высокой температуры тела, снижая ее всего до нескольких градусов выше нуля. Это сберегает энергетические ресурсы организма. Животные проводят в спячке часто долгие месяцы, иногда до полугода при резко пониженном уровне обмена веществ, постепенно расходуя накопленные резервы (рис. 21). У европейского суслика, например, частота сердцебиений падает с 400 в мин всего до 4–7, у хомяка обыкновенного – с 200 до 12–15, у сони-полчка – с 450 до 35. Потребление кислорода уменьшается в 20-100 раз (рис. 22). Однако состояние спячки не соответствует оцепенению пойкилотермных. Спячка – это хорошо регулируемое физиологическое состояние, при котором терморегуляторная система организма переключается на более низкий уровень. Когда температура в укрытии приближается к 0 °C, животные (хомяки, сурки, сони) начинают генерировать дополнительное тепло, поддерживая температуру тела на уровне не менее 2–3 °C За время спячки крылатые кожаны теряют 30–35 % веса, а суслики – до 50 %. Некоторые виды в зимний период неоднократно пробуждаются и подкармливаются запасами. Суслики до 7 % времени находятся в период спячки в бодрствующем состоянии. Пробуждение млекопитающих происходит путем самосогревания и требует затрат энергии. Возрастает частота сокращений сердца, учащается ритм дыхания. Для выхода из спячки сусликам требуется около 3 часов. Летучие мыши согреваются быстрее и через 15–30 минут уже готовы к полету.
Рис. 22. Изменение числа сердцебиений у сурка вовремя спячки при температуре тела (сверху вниз) 25, 17 и 16 °C. Одно деление шкалы равно 0,2 с (по Эндрес и др., 1930)
Торпидное состояние, или оцепенелость, сопровождает зимнюю спячку животных, но возникает и в других условиях как самостоятельная адаптация. Ряд мелких видов птиц и млекопитающих понижает уровень обмена и температуру тела при неблагоприятных погодных изменениях или регулярно в суточных циклах. У колибри, например, с наступлением ночи температура тела падает с 36–40 °C до 18 °C, птицы садятся на ветки и оцепеневают. Летучие мыши, наоборот, активны в сумеречное и ночное время и впадают в торпидное состояние днем. Переход к оцепенению стимулируется также недостатком пищи. При одних и тех же температурных условиях обеспеченные кормом животные могут продолжать активный образ жизни и генерировать тепло на терморегуляцию, а при нехватке пищи оцепеневают. Кратковременное торпидное состояние, как и спячка, – также регулируемый процесс, но температура тела при этом не падает столь низко и обычно остается на уровне, близком к 20 °C.
Между кратковременным оцепенением гетеротермных животных и длительной зимней спячкой существует вся гамма переходов. Зимний сон медведей, например, не называют спячкой, так как температура этих зверей понижается всего на 3–6 °C, а снижение уровня метаболизма не столь значительно. Мышечная активность обычно подавлена, но большинство физиологических функций сохраняется на прежнем уровне. Роды и выкармливание медвежат молоком у медведиц, например, приходятся на время зимнего сна. Медведи способны относительно легко пробуждаться и переходить в активное состояние.
В пустынях ряд мелких млекопитающих (мышевидные грызуны, сони, некоторые суслики, насекомоядные тенреки и др.) впадают не только в зимнюю, но и в летнюю спячку, которая называется эстивацией (от лат. aestes– лето). Температура их тела падает при этом не столь значительно – до 25–27 °C, но соответственное понижение уровня обмена веществ позволяет экономить не только энергию, но и воду.
Таким образом, широкое распространение гетеротермности позволяет предполагать фундаментальное значение этого явления для выживания животных в неблагоприятных условия.
3.2. Свет
3.2.1. Солнечная радиация
Всем живым организмам для осуществления процессов жизнедеятельности необходима энергия, поступающая извне. Основным источником ее является солнечная радиация, на которую приходится около 99,9 % в общем балансе энергии Земли.
Если принять солнечную энергию, достигающую Земли, за 100 %, то примерно 19 % ее поглощается при прохождении через атмосферу, 34 % отражается обратно в космическое пространство и 47 % достигает земной поверхности в виде прямой и рассеянной радиации (рис. 23). Прямая солнечная радиация – это континуум электромагнитного излучения с длинами волн от 0,1 до 30000 нм. На ультрафиолетовую часть спектра приходится от 1 до 5 %, на видимую – от 16 до 45 % и на инфракрасную – от 49 до 84 %.
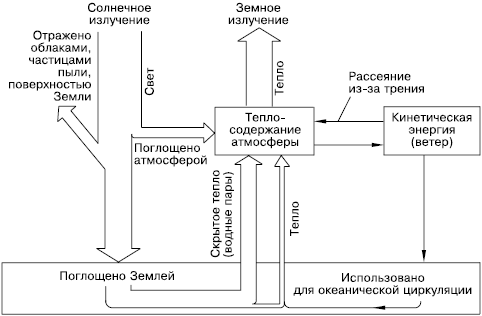
Рис. 23. Пути расходования солнечной энергии на поверхности Земли (по Э. Оорту, 1972)
Распределение энергии по спектру существенно зависит от массы атмосферы и меняется при различных высотах Солнца. Количество рассеянной радиации (отраженные лучи) возрастает с уменьшением высоты стояния Солнца и увеличением мутности атмосферы. Спектральный состав радиации безоблачного неба характеризуется максимумом энергии в 400–480 нм.
Если принять солнечную энергию, достигающую Земли, за 100 %, то примерно 19 % ее поглощается при прохождении через атмосферу, 34 % отражается обратно в космическое пространство и 47 % достигает земной поверхности в виде прямой и рассеянной радиации (рис. 23). Прямая солнечная радиация – это континуум электромагнитного излучения с длинами волн от 0,1 до 30000 нм. На ультрафиолетовую часть спектра приходится от 1 до 5 %, на видимую – от 16 до 45 % и на инфракрасную – от 49 до 84 %.
Рис. 23. Пути расходования солнечной энергии на поверхности Земли (по Э. Оорту, 1972)
Распределение энергии по спектру существенно зависит от массы атмосферы и меняется при различных высотах Солнца. Количество рассеянной радиации (отраженные лучи) возрастает с уменьшением высоты стояния Солнца и увеличением мутности атмосферы. Спектральный состав радиации безоблачного неба характеризуется максимумом энергии в 400–480 нм.