Страница:
У выращенных в неволе крупных хищных не было сформировано полноценное охотничье поведение, что приводило либо к голодной смерти хищника, либо к тому, что он начинал охотиться на домашних животных.
Следует обратить внимание еще на одну важную проблему, которая неоднократно обсуждалась на симпозиумах и конференциях, но не нашла отражения в литературе. На симпозиумах в Канаде (1982 г.) и Испании (1993 г.), участниками которых мы были, большое внимание уделялось обсуждению механизма денежной компенсации населению за ущерб, нанесенный реинтродуцированным хищником. В бюллетене программы восстановления мексиканского волка (Mexican Wolf Recovery Program, April, 1998) даны инструкции «What to do» на те случаи, если волк нанес ущерб фермерам или оказался на территории частных владений. В июльском же номере (Mexican Wolf Recovery Program, July, 1998) содержится информация о фактах нападения волка на домашних животных и случаях его непосредственного конфликта с человеком, окончившихся гибелью хищника. Подытоживая все эти сведения, можно сравнительно полно представить себе комплекс вопросов, связанных с восстановлением видов крупных млекопитающих и хищных в частности. Помимо указанных проблем, абсолютно не изучена реакция копытных на процесс восстановления хищных. В связи с этим очень важно определить темп реинтродукции и единовременную численность группы реинтродуцируемых крупных хищных.
Необходимо учесть и имеющийся в литературе небольшой материал относительно реакции диких животных на человека. В свое время Уильям Пруит в письме к А.Н. Формозову (см. Формозов, 1976) описал реакцию волка, с которым ему довелось встретиться на необитаемом острове Девон. До этого случая волк никогда не видел человека. Фотография, сделанная с расстояния двух метров, свидетельствует о буквально исследовательском интересе, проявленном волком к двуногому существу. Сам автор пишет, что волк «собирался играть подобно собаке».
В свое время мы описали звуки, которые издает дикий волк при опасности (Бадридзе, 1987 а, б, 1997; Badridze et al, 1992). Такие же звуки, так называемый «фыркающий лай», волки издают, когда чувствуют запах человека, его следа или слышат его голос. Подобное явление было ранее описано М. Павловым (1976) и А. Никольским с соавторами (1986).
Наблюдения за выращенными в неволе волками, имеющими потомство, а также за дикими волками в природе показали следующее: при сопряжении вышеописанных звуков родителей с признаками человека (запах, голос или комплексное восприятие) у щенков, вырабатывается генерализованная реакция избегания человека по принципу обучения с одной пробы — one trial (Бадридзе, 1987а, б, 1997; Badridze et al, 1992). В дальнейшем при самостоятельном восприятии признаков человека у щенков развивается полноценная реакция избегания.
Чтобы полностью очертить круг проблем, связанных с реинтродукцией крупных млекопитающих в естественную среду обитания, необходимо выяснить, какими поведенческими элементами должны обладать животные для их успешной реинтродукции в природу.
На основе приведенного выше материала можно сделать вывод, что метод реинтродукции в природу крупных млекопитающих фактически не разработан. Тут необходимы фундаментальные исследования. Интересно, что некоторые из них, на первый взгляд, непосредственно к проблеме реинтродукции не имеют никакого отношения. Поэтому целью нашего исследования было изучение закономерностей формирования поведения некоторых крупных млекопитающих, чтобы в дальнейшем, опираясь на эти закономерности, разработать методику реинтродукции.
Анализ литературного материала показал следующее:
К моменту реинтродукции у хищника должно быть сформировано не только полноценное охотничье поведение, необходима его специализация на естественный для данного хищника вид жертвы.
Территория, выбранная для реинтродукции, должна быть хорошо знакома реинтродуцируемому животному.
У животного должна быть выработана прочная реакция избегания незнакомых людей («знакомые», т. е. исследователи должны иметь возможность наблюдения), что предостережет его от браконьеров и возникновения конфликтной ситуации с представителями местного населения.
Реинтродуцируемые животные не должны нападать на домашних животных, что предотвратит возникновение антагонизма к ним со стороны местного населения и избавит от необходимости компенсации фермерам в случае урона, нанесенного хищником.
Наряду с этим необходимо, чтобы животные, отобранные для реинтродукции, были полноценными с точки зрения сформированных в неволе поведенческих комплексов, таких, как пищевой (включая пищедобывательный), социальный и репродуктивный. Наряду с этим необходимо не только знание закономерностей постнатального формирования рассудочной деятельности, но и создание таких условий для развития животных, при которых эта врожденная способность будет полноценно формироваться.
Для выбора оптимального метода исследования следует очертить те основные принципы, на базе которых возможна его дальнейшая разработка, а также избрать модельный вид. Он должен обладать следующими признаками: быть представителем отечественной фауны, иметь высокоразвитую психику и, желательно, совершенную социальную организацию. Последнее требование усложняет достижение поставленной цели, но при положительном результате обеспечит возможность применения этого метода к любым видам наземных крупных млекопитающих. Именно с учетом этих требований в качестве модельного вида нами был выбран волк. Для успешной реинтродукции в природу выращенных в неволе хищных млекопитающих у них должно быть сформировано полноценное охотничье поведение. Кроме того, хищник должен хорошо знать район будущей реинтродукции. Последнее объясняется необходимостью знания мест, выгодных для засады, возможных направлений выгона жертвы и основных путей ее круглосуточных перемещений.
Исходя из вышесказанного, во время подготовительного периода значительное время следует уделить освоению животным будущего места обитания.
Очевидно, что реализация реакции избегания любого типа возможна только на базе эмоциональной реакции страха. Понятно, что у выросшего в неволе животного выработать данную реакцию на базе естественного звукового раздражителя невозможно. Поэтому единственный путь — вызывать страх искусственно. Достичь этого можно двумя методами: I) электрическим раздражением соответствующих структур лимбической системы мозга; II) на базе эмоциональной реакции, возникающей на фоне боли, вызванной электрическим раздражением кожи.
В обоих случаях необходимо проведение полевых экспериментов, во время которых у животных должна быть возможность свободного передвижения. Поэтому тут требуется использование радиоуправляемых электростимуляторов.
Мы остановили свой выбор на втором методе, потому что он: а) не требует изготовления стереотаксического атласа мозга, что предполагает умерщвление большого количества животных, б) он более пригоден в полевых условиях. Очевидно, что болевое раздражение можно отнести к негуманным формам обращения с животными. В свое время И. Бентам (см. Игнатовская, 1988) писал, что «при этической оценке наших действий по отношению к любому существу, первое, что нам приходит в голову, это — может ли оно страдать?» (с.234). Это, конечно, истина, но вспомним случаи, когда хирургу в экстремальных ситуациях приходится делать операцию без наркоза, и это не считается аморальным. Процесс исчезновения вида — тоже экстремальная ситуация, и если возможно его спасение за счет причинения кратковременных неприятных ощущений нескольким десяткам особей, то не стоит отступать перед этим. Если принять данную позицию, то в процессе реинтродукции станет возможным выработать у животных реакцию избегания незнакомых людей и угасить хищническую реакцию на любых домашних животных. Это позволит исключить антагонизм местного населения к восстанавливаемому виду.
Материал и общая методика
РАЗДЕЛ 1. Отбор животных по поведенческим признакам
Глава 1.1. Значение техники искусственного выкармливания новорожденных хищных млекопитающих для формирования поведения взрослых
Материал и методика
Полученные результаты и их обсуждение
Глава 1.2. Поведение запасания пищи
Следует обратить внимание еще на одну важную проблему, которая неоднократно обсуждалась на симпозиумах и конференциях, но не нашла отражения в литературе. На симпозиумах в Канаде (1982 г.) и Испании (1993 г.), участниками которых мы были, большое внимание уделялось обсуждению механизма денежной компенсации населению за ущерб, нанесенный реинтродуцированным хищником. В бюллетене программы восстановления мексиканского волка (Mexican Wolf Recovery Program, April, 1998) даны инструкции «What to do» на те случаи, если волк нанес ущерб фермерам или оказался на территории частных владений. В июльском же номере (Mexican Wolf Recovery Program, July, 1998) содержится информация о фактах нападения волка на домашних животных и случаях его непосредственного конфликта с человеком, окончившихся гибелью хищника. Подытоживая все эти сведения, можно сравнительно полно представить себе комплекс вопросов, связанных с восстановлением видов крупных млекопитающих и хищных в частности. Помимо указанных проблем, абсолютно не изучена реакция копытных на процесс восстановления хищных. В связи с этим очень важно определить темп реинтродукции и единовременную численность группы реинтродуцируемых крупных хищных.
Необходимо учесть и имеющийся в литературе небольшой материал относительно реакции диких животных на человека. В свое время Уильям Пруит в письме к А.Н. Формозову (см. Формозов, 1976) описал реакцию волка, с которым ему довелось встретиться на необитаемом острове Девон. До этого случая волк никогда не видел человека. Фотография, сделанная с расстояния двух метров, свидетельствует о буквально исследовательском интересе, проявленном волком к двуногому существу. Сам автор пишет, что волк «собирался играть подобно собаке».
В свое время мы описали звуки, которые издает дикий волк при опасности (Бадридзе, 1987 а, б, 1997; Badridze et al, 1992). Такие же звуки, так называемый «фыркающий лай», волки издают, когда чувствуют запах человека, его следа или слышат его голос. Подобное явление было ранее описано М. Павловым (1976) и А. Никольским с соавторами (1986).
Наблюдения за выращенными в неволе волками, имеющими потомство, а также за дикими волками в природе показали следующее: при сопряжении вышеописанных звуков родителей с признаками человека (запах, голос или комплексное восприятие) у щенков, вырабатывается генерализованная реакция избегания человека по принципу обучения с одной пробы — one trial (Бадридзе, 1987а, б, 1997; Badridze et al, 1992). В дальнейшем при самостоятельном восприятии признаков человека у щенков развивается полноценная реакция избегания.
Чтобы полностью очертить круг проблем, связанных с реинтродукцией крупных млекопитающих в естественную среду обитания, необходимо выяснить, какими поведенческими элементами должны обладать животные для их успешной реинтродукции в природу.
На основе приведенного выше материала можно сделать вывод, что метод реинтродукции в природу крупных млекопитающих фактически не разработан. Тут необходимы фундаментальные исследования. Интересно, что некоторые из них, на первый взгляд, непосредственно к проблеме реинтродукции не имеют никакого отношения. Поэтому целью нашего исследования было изучение закономерностей формирования поведения некоторых крупных млекопитающих, чтобы в дальнейшем, опираясь на эти закономерности, разработать методику реинтродукции.
Анализ литературного материала показал следующее:
К моменту реинтродукции у хищника должно быть сформировано не только полноценное охотничье поведение, необходима его специализация на естественный для данного хищника вид жертвы.
Территория, выбранная для реинтродукции, должна быть хорошо знакома реинтродуцируемому животному.
У животного должна быть выработана прочная реакция избегания незнакомых людей («знакомые», т. е. исследователи должны иметь возможность наблюдения), что предостережет его от браконьеров и возникновения конфликтной ситуации с представителями местного населения.
Реинтродуцируемые животные не должны нападать на домашних животных, что предотвратит возникновение антагонизма к ним со стороны местного населения и избавит от необходимости компенсации фермерам в случае урона, нанесенного хищником.
Наряду с этим необходимо, чтобы животные, отобранные для реинтродукции, были полноценными с точки зрения сформированных в неволе поведенческих комплексов, таких, как пищевой (включая пищедобывательный), социальный и репродуктивный. Наряду с этим необходимо не только знание закономерностей постнатального формирования рассудочной деятельности, но и создание таких условий для развития животных, при которых эта врожденная способность будет полноценно формироваться.
* * *
Для выбора оптимального метода исследования следует очертить те основные принципы, на базе которых возможна его дальнейшая разработка, а также избрать модельный вид. Он должен обладать следующими признаками: быть представителем отечественной фауны, иметь высокоразвитую психику и, желательно, совершенную социальную организацию. Последнее требование усложняет достижение поставленной цели, но при положительном результате обеспечит возможность применения этого метода к любым видам наземных крупных млекопитающих. Именно с учетом этих требований в качестве модельного вида нами был выбран волк. Для успешной реинтродукции в природу выращенных в неволе хищных млекопитающих у них должно быть сформировано полноценное охотничье поведение. Кроме того, хищник должен хорошо знать район будущей реинтродукции. Последнее объясняется необходимостью знания мест, выгодных для засады, возможных направлений выгона жертвы и основных путей ее круглосуточных перемещений.
Исходя из вышесказанного, во время подготовительного периода значительное время следует уделить освоению животным будущего места обитания.
Очевидно, что реализация реакции избегания любого типа возможна только на базе эмоциональной реакции страха. Понятно, что у выросшего в неволе животного выработать данную реакцию на базе естественного звукового раздражителя невозможно. Поэтому единственный путь — вызывать страх искусственно. Достичь этого можно двумя методами: I) электрическим раздражением соответствующих структур лимбической системы мозга; II) на базе эмоциональной реакции, возникающей на фоне боли, вызванной электрическим раздражением кожи.
В обоих случаях необходимо проведение полевых экспериментов, во время которых у животных должна быть возможность свободного передвижения. Поэтому тут требуется использование радиоуправляемых электростимуляторов.
Мы остановили свой выбор на втором методе, потому что он: а) не требует изготовления стереотаксического атласа мозга, что предполагает умерщвление большого количества животных, б) он более пригоден в полевых условиях. Очевидно, что болевое раздражение можно отнести к негуманным формам обращения с животными. В свое время И. Бентам (см. Игнатовская, 1988) писал, что «при этической оценке наших действий по отношению к любому существу, первое, что нам приходит в голову, это — может ли оно страдать?» (с.234). Это, конечно, истина, но вспомним случаи, когда хирургу в экстремальных ситуациях приходится делать операцию без наркоза, и это не считается аморальным. Процесс исчезновения вида — тоже экстремальная ситуация, и если возможно его спасение за счет причинения кратковременных неприятных ощущений нескольким десяткам особей, то не стоит отступать перед этим. Если принять данную позицию, то в процессе реинтродукции станет возможным выработать у животных реакцию избегания незнакомых людей и угасить хищническую реакцию на любых домашних животных. Это позволит исключить антагонизм местного населения к восстанавливаемому виду.
Материал и общая методика
Приведенный в данной работе материал собран с апреля 1974 года по февраль 1995 г. включительно. Исследования проводились на содержащихся в неволе 96 волках (Canis Lupus cubanensis Ognev, — 12 групп) до 1-месячного возраста, из них на 78 волках (10 групп) — они продолжались до 5-месячного возраста, а над 39 животными (9 групп) — до их полового созревания и впоследствии.
Эксперименты и наблюдения проводились как в полевых условиях, так и в вольерах (100 кв. м). Для сравнительного этологического анализа мы приводим материал, полученный в результате наблюдений за следующими животными: 3 тигра (P. t. atlaica — 1 самец и 1 самка; P. t. corbetti — 1 самец), 3 леопарда (Pantera pardus — 1 самка и 2 самца), 20 сторожевых собак, агрессивных к незнакомым людям (10 групп, от рождения до достижения половой зрелости); 24 взрослые козы, 16 овец, 11 лошадей и 16 ослов.
Непосредственно к реинтродукции готовили 22 выращенных в неволе волка (4 группы). Причину, по которой был избран именно этот вид, мы обосновали выше. Кроме того, наблюдения проводились и за шестью семьями дикоживущих волков, привыкших к моему присутствию (продолжительность наблюдений — 1463 часа). Под наблюдением также находились 7 диких рысей с котятами (Felis (lynx) lynx orientalis — 2 часа), 860 самок оленей с оленятами (Cervus elaphus maral), 300 дагестанских туров с козлятами (Capra cylindricornis) и 37 стай одичавших собак (общая продолжительность наблюдений за последними — 584 часа). Одним из способов анализа полученного материала было сравнение данных, полученных на различных видах (диких, выращенных в неволе и домашних). Необходимость этого продиктована тем, что поведение выращенных в неволе животных может видоизменяться. Кроме того, при наблюдении за дикоживущими животными, вследствие естественных трудностей, не всегда удается полноценно проследить закономерности формирования поведения. Поэтому наиболее полную возможность выявить общие закономерности формирования и развития поведения дает сопоставление материалов, полученных обоими методами. Во всех случаях достоверность полученного материала рассчитывали по t критерию Стьюдента (Лакин, 1980).
Эксперименты и наблюдения проводились как в полевых условиях, так и в вольерах (100 кв. м). Для сравнительного этологического анализа мы приводим материал, полученный в результате наблюдений за следующими животными: 3 тигра (P. t. atlaica — 1 самец и 1 самка; P. t. corbetti — 1 самец), 3 леопарда (Pantera pardus — 1 самка и 2 самца), 20 сторожевых собак, агрессивных к незнакомым людям (10 групп, от рождения до достижения половой зрелости); 24 взрослые козы, 16 овец, 11 лошадей и 16 ослов.
Непосредственно к реинтродукции готовили 22 выращенных в неволе волка (4 группы). Причину, по которой был избран именно этот вид, мы обосновали выше. Кроме того, наблюдения проводились и за шестью семьями дикоживущих волков, привыкших к моему присутствию (продолжительность наблюдений — 1463 часа). Под наблюдением также находились 7 диких рысей с котятами (Felis (lynx) lynx orientalis — 2 часа), 860 самок оленей с оленятами (Cervus elaphus maral), 300 дагестанских туров с козлятами (Capra cylindricornis) и 37 стай одичавших собак (общая продолжительность наблюдений за последними — 584 часа). Одним из способов анализа полученного материала было сравнение данных, полученных на различных видах (диких, выращенных в неволе и домашних). Необходимость этого продиктована тем, что поведение выращенных в неволе животных может видоизменяться. Кроме того, при наблюдении за дикоживущими животными, вследствие естественных трудностей, не всегда удается полноценно проследить закономерности формирования поведения. Поэтому наиболее полную возможность выявить общие закономерности формирования и развития поведения дает сопоставление материалов, полученных обоими методами. Во всех случаях достоверность полученного материала рассчитывали по t критерию Стьюдента (Лакин, 1980).
РАЗДЕЛ 1. Отбор животных по поведенческим признакам
Обсуждаемые в данном разделе вопросы касаются в основном поведенческих признаков волка, так как именно он был для нас модельным видом. Естественно, что для исследования проблемы реинтродукции в природу выращенных в неволе других видов крупных хищных млекопитающих придется рассматривать другой спектр поведенческих элементов и, соответственно, производить отбор на основе признаков их сформированности.
Глава 1.1. Значение техники искусственного выкармливания новорожденных хищных млекопитающих для формирования поведения взрослых
Поведенческие реакции, сформированные к моменту рождения незрелорождающихся животных и способствующие их первому контакту с окружающей средой, довольно немногочисленны и, судя по литературным данным (Уждавини, 1958; Уждавини, Шепелова, 1966; Слоним, 1967, 1976; Хайнд, 1975; Бадридзе, 1982, 1983 б, 1985, 1987 а, б; Harlow amp; Zimmerman, 1959 и др.), довольно хорошо изучены. Они «как бы пригнаны к условиям существования сразу же после рождения» (Уждавини, Шепелова, 1966, с.106). О подготовленности реакций новорожденных к экологии вида писал и академик П.К. Анохин (1962). Опыты, проведенные в лаборатории А.Д. Слонима (1976), показали, что наиболее выражены, запрограммированные в эмбриональном периоде реакции на кормящую мать. Авторы установили, что можно не только наблюдать врожденные реакции на мать как на внешний фактор, но и моделировать эти внешние факторы. Подобное моделирование проводили и мы (Бадридзе, 1983 а, б; 1 985 1987 а, б). Такой подход способствует выявлению тех раздражителей, на которые возникают конкретные реакции новорожденного животного, что позволяет заполнить пробелы в изучении экологии неонатального периода тех видов, условия обитания которых не дают или ограничивают возможность исследований.
Даже в таких значительных работах, как исследования Фокса (Fox, 1971) и Мича (Mech, 1970), касающихся поведения и экологии представителей семейства Canidae и, в частности, волка, нет каких-либо существенных данных об экологической адекватности поведения новорожденных животных. Это можно объяснить большой сложностью наблюдений в природе за контактом новорожденных животных (особенно рождающихся в логове) с матерью и остальной внешней средой, в ответ на воздействие которых развиваются строго определенные врожденные реакции. В лабораторных условиях Фокс (Fox, 1964, 1971) детально исследовал и описал неонатальные реакции Canidae и, в том числе, волка. К сожалению, ни в одной из упомянутых работ нет данных о том, какую роль играет полноценная реализация неонатальных реакций для формирования нормального поведения в постлактационном периоде. Поэтому в данной главе мы попытаемся показать значение полноценной реализации врожденных поведенческих элементов, связанных с процессом питания новорожденных хищных млекопитающих для дальнейшего формирования поведения в пределах нормы.
Даже в таких значительных работах, как исследования Фокса (Fox, 1971) и Мича (Mech, 1970), касающихся поведения и экологии представителей семейства Canidae и, в частности, волка, нет каких-либо существенных данных об экологической адекватности поведения новорожденных животных. Это можно объяснить большой сложностью наблюдений в природе за контактом новорожденных животных (особенно рождающихся в логове) с матерью и остальной внешней средой, в ответ на воздействие которых развиваются строго определенные врожденные реакции. В лабораторных условиях Фокс (Fox, 1964, 1971) детально исследовал и описал неонатальные реакции Canidae и, в том числе, волка. К сожалению, ни в одной из упомянутых работ нет данных о том, какую роль играет полноценная реализация неонатальных реакций для формирования нормального поведения в постлактационном периоде. Поэтому в данной главе мы попытаемся показать значение полноценной реализации врожденных поведенческих элементов, связанных с процессом питания новорожденных хищных млекопитающих для дальнейшего формирования поведения в пределах нормы.
Материал и методика
Эксперименты проводили на 96 новорожденных волчатах, 67 щенках собак (далее — щенки), 30 новорожденных котятах. 10 волчат, 15 щенков и 10 котят были взяты у матери в момент рождения. Возраст отнятия от матери 37 волчат и 15 щенков составлял до 40 часов, а 49 волчат и 37 щенков — 7—10 дней. Всех животных вскармливали с помощью бутылок с соской. Во время кормления 20 волчат и 15 щенков (I контрольная группа) на бутылку с соской надевали диск, диаметр которого составлял 200 мм (рис. 2А), чтобы у новорожденных была возможность упираться лапами во время сосания. Прокол в соске был таков, чтобы не позволять молоку вытекать без сосательных усилий.
10 волчат, 15 щенков и 10 котят (I экспериментальная группа) с момента рождения кормили из сосок без диска, тем самым лишая их возможности упираться во что-либо передними лапками. Остальных котят до окончания лактационного периода вскармливала мать.
Во всех случаях у основания бутылки было проделано отверстие диаметром 1 мм для уравнивания давления во время сосания внутри бутылки с атмосферным. Значение этого в свое время было описано (например, Бадридзе, 1987 а, б).
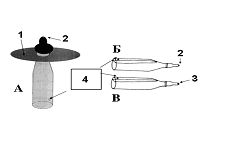
Рисунок 2.
Типы применяемых сосок (А, Б, В). 1. Диск; 2. Прокол; 3. Отверстие — 2мм; 4. Отверстия.
При вскармливании животных II экспериментальной группы (10 волчат и 10 щенков, отнятых у матери в возрасте 6—40 часов) также применяли два типа сосок: I тип — с проколом, который не позволял молоку вытекать без сосательных усилий, тем самым приближая процесс сосания к естественному (рис. 2Б) и II тип — с отверстием диаметром 2 мм (рис. 2В). При сосании соски с большим отверстием детенышам не приходилось прилагать значительных усилий, и желудок у них наполнялся после двух-четырех сосательных движений.
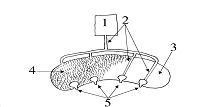 Рисунок 3.
Рисунок 3.
Приспособление, имитирующее тело матери.
1-резервуар с молоком; 2-трубки, проводящие молоко; 3-резиновый баллон с водой 36—38°C; 4-кроличья шкурка; 5-искусственные сосцы.
Остальных животных (II контрольная группа, 56 волчат и 27 щенков) вскармливали с помощью сконструированной нами установки, имитирующей тело матери. Установка представляла собой покрытый кроличьими шкурками изогнутый дугой резиновый баллон (рис.3). Сосцы прикреплялись к баллону со стороны меньшего радиуса. Они соединялись мягкими пластиковыми трубками, пронизывающими баллон. В полость баллона заливалась теплая вода (36—38 градусов), которая нагревала находящееся в трубках молоко. Пастеризованное молоко для вскармливания щенков и волчат приготавливали из молочного порошка, имитируя концентрацию собачьего молока (Барабаш — Никифоров, Формозов, 1963).
В результате наблюдения за животными в условиях искусственного вскармливания нами были изучены следующие вопросы:
Влияние полноценной реализации «реакции массажа лапками молочной железы» во время сосания на формирование психически уравновешенного взрослого животного.
Влияние скорости поступления молока во время сосания в первую неделю жизни на дальнейшее формирование порога сенсорного насыщения.
10 волчат, 15 щенков и 10 котят (I экспериментальная группа) с момента рождения кормили из сосок без диска, тем самым лишая их возможности упираться во что-либо передними лапками. Остальных котят до окончания лактационного периода вскармливала мать.
Во всех случаях у основания бутылки было проделано отверстие диаметром 1 мм для уравнивания давления во время сосания внутри бутылки с атмосферным. Значение этого в свое время было описано (например, Бадридзе, 1987 а, б).
Рисунок 2.
Типы применяемых сосок (А, Б, В). 1. Диск; 2. Прокол; 3. Отверстие — 2мм; 4. Отверстия.
При вскармливании животных II экспериментальной группы (10 волчат и 10 щенков, отнятых у матери в возрасте 6—40 часов) также применяли два типа сосок: I тип — с проколом, который не позволял молоку вытекать без сосательных усилий, тем самым приближая процесс сосания к естественному (рис. 2Б) и II тип — с отверстием диаметром 2 мм (рис. 2В). При сосании соски с большим отверстием детенышам не приходилось прилагать значительных усилий, и желудок у них наполнялся после двух-четырех сосательных движений.
Приспособление, имитирующее тело матери.
1-резервуар с молоком; 2-трубки, проводящие молоко; 3-резиновый баллон с водой 36—38°C; 4-кроличья шкурка; 5-искусственные сосцы.
Остальных животных (II контрольная группа, 56 волчат и 27 щенков) вскармливали с помощью сконструированной нами установки, имитирующей тело матери. Установка представляла собой покрытый кроличьими шкурками изогнутый дугой резиновый баллон (рис.3). Сосцы прикреплялись к баллону со стороны меньшего радиуса. Они соединялись мягкими пластиковыми трубками, пронизывающими баллон. В полость баллона заливалась теплая вода (36—38 градусов), которая нагревала находящееся в трубках молоко. Пастеризованное молоко для вскармливания щенков и волчат приготавливали из молочного порошка, имитируя концентрацию собачьего молока (Барабаш — Никифоров, Формозов, 1963).
В результате наблюдения за животными в условиях искусственного вскармливания нами были изучены следующие вопросы:
Влияние полноценной реализации «реакции массажа лапками молочной железы» во время сосания на формирование психически уравновешенного взрослого животного.
Влияние скорости поступления молока во время сосания в первую неделю жизни на дальнейшее формирование порога сенсорного насыщения.
Полученные результаты и их обсуждение
В свое время мы показали, что «реакция массажа лапками молочной железы» во время сосания в постлактационный период трансформируется в манипуляторную активность при разделке пищи (Бадридзе, 1982, 1983 б, 1985, 1987 а, б. Рис. 4 А, Б).
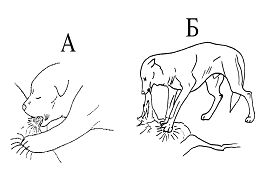 Рисунок 4.
Рисунок 4.
Функциональная трансформация реакции массажа лапками молочной железы во время сосания.
А — лактационный период;
Б — постлактационный период.
«Реакция массажа лапками молочной железы» во время сосания представляет собой результат последовательного сокращения разгибательных и сгибательных мышц передних конечностей. В наших экспериментах, когда животных лишали возможности упора лапками во что-нибудь (I экспериментальная группа — 15 щенков, 10 волчат и 10 котят), у них появлялось тоническое напряжение передних конечностей. Это выражалось в растопыривании пальцев и развитии одновременного напряжения сгибательных и разгибательных мышц. Подобное состояние продолжалось до окончания акта сосания. Те новорожденные (I и II контрольные группы), у которых постоянно была возможность опираться на что-либо передними лапками (бутылочка с диском, либо прибор, имитирующий тело матери), осуществлялась нормальная «реакция массажа лапками», т. е. последовательное сокращение реципрокных мышц передних конечностей.
В возрасте, когда хищники переходят на полный мясной рацион (40—50 дней), животным начали давать по крупному куску мяса. При первом же его предъявлении все животные контрольных групп начинали его разделывать причем с первого же момента все манипуляционные движения лап были полностью координированы.
Иная картина наблюдалась в I экспериментальной группе. Здесь для формирования координированной манипуляторной активности требовалось 3—4 дня. У этих животных заметно проявлялось т. н. «переступание лапами на месте». Параллельно с этим, животные время от времени растопыривали пальцы. Со временем тик ослабевал, но в малозаметной форме сохранялся до конца эксперимента. Кроме того, у всех животных в течение всего периода наблюдений была заметна эмоциональная неуравновешенность, что проявлялось в сравнительно низком пороге оборонительного поведения и частых фрустрациях. Эти индивиды часто создавали в группе (с т. н. нормальными) конфликтные ситуации. Соотношение животных контрольной и экспериментальной групп, провоцирующих конфликт, составляло 1/1,25. Волчата и щенки контрольных групп развивались эмоционально нормальными и способны были создать группу с социальными отношениями в пределах нормы.
До обсуждения возможных причин подобного различия считаем нужным напомнить данные, приведенные Г. Шепердом (1987), согласно которым импульсация от мышечных и суставных рецепторов поступает через таламус в соответствующую область коры головного мозга. Общеизвестно, что у незрелорождающихся млекопитающих формирование мозговых систем продолжается после рождения и, судя по всему, оптимальный уровень центростремительной импульсации имеет большое значение для их нормального формирования и функционирования. Во время сосания у животных I экспериментальной группы постоянно развивалось тоническое напряжение мышц передних конечностей, что должно было определять центростремительную гиперимпульсацию к соответствующим мозговым структурам.
Из сказанного можно предположить, что в период формирования мозговых систем подобная гиперимпульсация может создавать очаги с постоянно высоким уровнем возбуждения, что, в свою очередь, влияет на уровень эмоциональной уравновешенности животного. Из приведенного материала следует, что животные, выращенные при подобных условиях питания в неонатальном периоде, не могут быть пригодными для дальнейшей их реинтродукции.
Сравнивая различия в уровне эмоциональной уравновешенности между животными I экспериментальной и контрольными группами, можно заключить, что из-за невозможности нормальной реализации «реакции массажа лапками молочной железы во время сосания» (I экспериментальная группа), развивается одновременное тоническое напряжение всех групп мышц передних конечностей, вызывает возникновение в мозгу очагов постоянно высокого возбуждения. Судя по всему, это и стало причиной эмоциональной неуравновешенности животных этой группы. Животные же контрольных групп имели возможность реализации данной реакции, что, по видимому, и определило нормальное формирование их эмоциональной возбудимости, т. к. у них в мозгу не возникают очаги с постоянно высоким возбуждением.
Как было сказано ранее, животных II экспериментальной группы кормили с помощью сосок с отверстием диаметра 2 мм. У волчат и щенков, вскармливаемых из такой соски, желудки наполнялись после нескольких сосательных движений, в течение 0,5 минуты (60—80 мл). Во время подобных экспериментов все животные, несмотря на быстрое наполнение желудка, продолжали проявлять сосательную активность. В свое время нами было показано, что у новорожденных щенят и волчат пищевая мотивация возникает к концу первой недели жизни (Бадридзе, 1987 а, б), а до этого процесс приема пищи осуществляется за счет потребности в реализации сосательной активности. Животные этой группы продолжали принятие чрезмерного количества пищи и после формирования пищевой мотивационной системы. Стремление к перееданию сохраняется в течение всей жизни, что создает постоянные внутригрупповые конфликты на почве конкуренции за пищу. В свое время было показано (Анохин, 1962; Бакурадзе, Чхенкели, 1970 а, б; Бадридзе, 1987 а, б и др.), что включение центральных механизмов головного мозга, тормозящих акт питания, происходит за счет сенсорного насыщения, т. е. центростремительной импульсации, возникающей при растяжении стенок желудка.
Анализируя полученный нами материал, можно заключить, что чрезмерный прием пищи в раннем неонатальном периоде может увеличить объем желудка и, следовательно, порог сенсорного (первичного) насыщения, что и определяет развитие относительной гиперфагии. Такие животные активно конкурируют из-за пищи до наступления вторичного насыщения, чем постоянно создают конфликтную ситуацию во время еды. Вместе с тем у них, как правило, не развивается поведение запасания пищи, значение которого будет обсуждаться в следующей главе.
Из всего вышесказанного следует, что для нормального развития новорожденных животных, отлученных от матери, решающее значение имеют т. н. «технические» условия при приеме пищи, которые максимально должны быть приближены к естественным. Животные, выращенные в иных условиях, не могут быть пригодны для реинтродукции, т. к. не способны поддерживать в группе полноценные социальные взаимоотношения.
Из полученного нами материала можно сделать следующий основной вывод — при искусственном вскармливании хищных млекопитающих для нормального формирования поведения особое значение имеет неонатальный период, который в значительной мере определяет дальнейшее развитие поведения в пределах нормы или патологии.
В частности:
— для формирования оптимального уровня эмоциональной возбудимости большое значение имеет возможность полноценной реализации реакции «массажа лапками молочной железы во время сосания»;
— для исключения высокого порога сенсорного насыщения и, исходя из этого, высокого уровня конкурентных ситуаций в группе во время питания, необходимо условия выкармливания животных максимально приблизить к естественному процессу сосания.
Функциональная трансформация реакции массажа лапками молочной железы во время сосания.
А — лактационный период;
Б — постлактационный период.
«Реакция массажа лапками молочной железы» во время сосания представляет собой результат последовательного сокращения разгибательных и сгибательных мышц передних конечностей. В наших экспериментах, когда животных лишали возможности упора лапками во что-нибудь (I экспериментальная группа — 15 щенков, 10 волчат и 10 котят), у них появлялось тоническое напряжение передних конечностей. Это выражалось в растопыривании пальцев и развитии одновременного напряжения сгибательных и разгибательных мышц. Подобное состояние продолжалось до окончания акта сосания. Те новорожденные (I и II контрольные группы), у которых постоянно была возможность опираться на что-либо передними лапками (бутылочка с диском, либо прибор, имитирующий тело матери), осуществлялась нормальная «реакция массажа лапками», т. е. последовательное сокращение реципрокных мышц передних конечностей.
В возрасте, когда хищники переходят на полный мясной рацион (40—50 дней), животным начали давать по крупному куску мяса. При первом же его предъявлении все животные контрольных групп начинали его разделывать причем с первого же момента все манипуляционные движения лап были полностью координированы.
Иная картина наблюдалась в I экспериментальной группе. Здесь для формирования координированной манипуляторной активности требовалось 3—4 дня. У этих животных заметно проявлялось т. н. «переступание лапами на месте». Параллельно с этим, животные время от времени растопыривали пальцы. Со временем тик ослабевал, но в малозаметной форме сохранялся до конца эксперимента. Кроме того, у всех животных в течение всего периода наблюдений была заметна эмоциональная неуравновешенность, что проявлялось в сравнительно низком пороге оборонительного поведения и частых фрустрациях. Эти индивиды часто создавали в группе (с т. н. нормальными) конфликтные ситуации. Соотношение животных контрольной и экспериментальной групп, провоцирующих конфликт, составляло 1/1,25. Волчата и щенки контрольных групп развивались эмоционально нормальными и способны были создать группу с социальными отношениями в пределах нормы.
До обсуждения возможных причин подобного различия считаем нужным напомнить данные, приведенные Г. Шепердом (1987), согласно которым импульсация от мышечных и суставных рецепторов поступает через таламус в соответствующую область коры головного мозга. Общеизвестно, что у незрелорождающихся млекопитающих формирование мозговых систем продолжается после рождения и, судя по всему, оптимальный уровень центростремительной импульсации имеет большое значение для их нормального формирования и функционирования. Во время сосания у животных I экспериментальной группы постоянно развивалось тоническое напряжение мышц передних конечностей, что должно было определять центростремительную гиперимпульсацию к соответствующим мозговым структурам.
Из сказанного можно предположить, что в период формирования мозговых систем подобная гиперимпульсация может создавать очаги с постоянно высоким уровнем возбуждения, что, в свою очередь, влияет на уровень эмоциональной уравновешенности животного. Из приведенного материала следует, что животные, выращенные при подобных условиях питания в неонатальном периоде, не могут быть пригодными для дальнейшей их реинтродукции.
Сравнивая различия в уровне эмоциональной уравновешенности между животными I экспериментальной и контрольными группами, можно заключить, что из-за невозможности нормальной реализации «реакции массажа лапками молочной железы во время сосания» (I экспериментальная группа), развивается одновременное тоническое напряжение всех групп мышц передних конечностей, вызывает возникновение в мозгу очагов постоянно высокого возбуждения. Судя по всему, это и стало причиной эмоциональной неуравновешенности животных этой группы. Животные же контрольных групп имели возможность реализации данной реакции, что, по видимому, и определило нормальное формирование их эмоциональной возбудимости, т. к. у них в мозгу не возникают очаги с постоянно высоким возбуждением.
Как было сказано ранее, животных II экспериментальной группы кормили с помощью сосок с отверстием диаметра 2 мм. У волчат и щенков, вскармливаемых из такой соски, желудки наполнялись после нескольких сосательных движений, в течение 0,5 минуты (60—80 мл). Во время подобных экспериментов все животные, несмотря на быстрое наполнение желудка, продолжали проявлять сосательную активность. В свое время нами было показано, что у новорожденных щенят и волчат пищевая мотивация возникает к концу первой недели жизни (Бадридзе, 1987 а, б), а до этого процесс приема пищи осуществляется за счет потребности в реализации сосательной активности. Животные этой группы продолжали принятие чрезмерного количества пищи и после формирования пищевой мотивационной системы. Стремление к перееданию сохраняется в течение всей жизни, что создает постоянные внутригрупповые конфликты на почве конкуренции за пищу. В свое время было показано (Анохин, 1962; Бакурадзе, Чхенкели, 1970 а, б; Бадридзе, 1987 а, б и др.), что включение центральных механизмов головного мозга, тормозящих акт питания, происходит за счет сенсорного насыщения, т. е. центростремительной импульсации, возникающей при растяжении стенок желудка.
Анализируя полученный нами материал, можно заключить, что чрезмерный прием пищи в раннем неонатальном периоде может увеличить объем желудка и, следовательно, порог сенсорного (первичного) насыщения, что и определяет развитие относительной гиперфагии. Такие животные активно конкурируют из-за пищи до наступления вторичного насыщения, чем постоянно создают конфликтную ситуацию во время еды. Вместе с тем у них, как правило, не развивается поведение запасания пищи, значение которого будет обсуждаться в следующей главе.
Из всего вышесказанного следует, что для нормального развития новорожденных животных, отлученных от матери, решающее значение имеют т. н. «технические» условия при приеме пищи, которые максимально должны быть приближены к естественным. Животные, выращенные в иных условиях, не могут быть пригодны для реинтродукции, т. к. не способны поддерживать в группе полноценные социальные взаимоотношения.
Из полученного нами материала можно сделать следующий основной вывод — при искусственном вскармливании хищных млекопитающих для нормального формирования поведения особое значение имеет неонатальный период, который в значительной мере определяет дальнейшее развитие поведения в пределах нормы или патологии.
В частности:
— для формирования оптимального уровня эмоциональной возбудимости большое значение имеет возможность полноценной реализации реакции «массажа лапками молочной железы во время сосания»;
— для исключения высокого порога сенсорного насыщения и, исходя из этого, высокого уровня конкурентных ситуаций в группе во время питания, необходимо условия выкармливания животных максимально приблизить к естественному процессу сосания.
Глава 1.2. Поведение запасания пищи
Запасание корма является одним из врожденных видов пищевого поведения, широко распространенного среди животных (Свириденко, 1957; Слоним, 1967; Хайнд, 1975 и др.) Из хищных млекопитающих пищей запасаются собаки, волки, лисицы и др. (Мертц, 1953; Кудактин, 1977; Поярков, 1980; Корытин, 1981; Jeselnik, Brisbin, 1980 и др.) Наиболее полные из имеющихся в литературе сведений по этому вопросу можно найти у Мича (Mech, 1970), однако и их явно недостаточно для исчерпывающего понимания феномена запасания пищи волками.
Мич также подчеркивает, что о запасании волками пищи и о том, как это происходит, известно очень мало. Упоминания об этом поведении волка содержатся и в работах Н.А. Зворыкина (1936) и Б.Т. Семенова (1979). В свое время мы изучали закономерность формирования поведения волков при запасании пищи, зависимость способа транспортировки запасаемой пищи от социального статуса особи и специфику памяти на местонахождение кладовых (Бадридзе, 1979, 1987 а, б; Бадридзе и соавт., 1997). К сожалению, мы не встречали в литературных источниках данных, касающихся значения данного вида поведения как для индивида, так и для всей группы в целом и, исходя из этого, для всей популяции. Неизвестно также, может ли существовать естественная патология в формировании данного поведения. Мы имеем в виду естественную патологию, так как причины, из-за которых не формируется поведение запасания пищи при искусственном выкармливании новорожденных животных, уже обсуждались в главе 1.1.
Поэтому в данной главе будут освещены вопросы, касающиеся значения поведения запасания для пищевого благополучия индивида, группы и, исходя их этого, для популяции в целом. Изучение данного вопроса должно иметь большое значение для отбора животных по признаку сформированности этого поведения к моменту реинтродукции в природу.
Мич также подчеркивает, что о запасании волками пищи и о том, как это происходит, известно очень мало. Упоминания об этом поведении волка содержатся и в работах Н.А. Зворыкина (1936) и Б.Т. Семенова (1979). В свое время мы изучали закономерность формирования поведения волков при запасании пищи, зависимость способа транспортировки запасаемой пищи от социального статуса особи и специфику памяти на местонахождение кладовых (Бадридзе, 1979, 1987 а, б; Бадридзе и соавт., 1997). К сожалению, мы не встречали в литературных источниках данных, касающихся значения данного вида поведения как для индивида, так и для всей группы в целом и, исходя из этого, для всей популяции. Неизвестно также, может ли существовать естественная патология в формировании данного поведения. Мы имеем в виду естественную патологию, так как причины, из-за которых не формируется поведение запасания пищи при искусственном выкармливании новорожденных животных, уже обсуждались в главе 1.1.
Поэтому в данной главе будут освещены вопросы, касающиеся значения поведения запасания для пищевого благополучия индивида, группы и, исходя их этого, для популяции в целом. Изучение данного вопроса должно иметь большое значение для отбора животных по признаку сформированности этого поведения к моменту реинтродукции в природу.