Страница:
В этот период было показано, что большинство перечисленных в предыдущем разделе «романтических» законов экологии имеет ограниченные области экстраполяции. Исключений из этих «законов-правил» оказалось так много, что возникло сомнение в их правомочности. В этом непринятии идеи существования универсальных экологических законов, подобных законам физики, – идеология современной экологии.
Принцип экологической индивидуальности видов и их независимого распределения по градиентам среды в соответствии с симметричной колоколовидной кривой оказался не соответствующим действительности. В ряде случаев кривые распределения были асимметричными и даже полимодальными, что свидетельствовало о зависимости распределений видов друг от друга.
Как оказалось, продуктивными и богатыми видами могут быть не только климаксовые, устойчивые, но и сообщества, которые меняются в направлении климакса. Конвергенция всего разнообразия экосистем природного района в одну климаксовую экосистему просто невозможна, в каждом природном районе есть несколько климаксовых экосистем. Сукцессии, ведущие к климаксу, оказались стохастическими, а не жестко детерминированными процессами, в ходе которых виды сменяют друг друга в четкой последовательности. В ходе сукцессии не обязательно происходит улучшение условий, повышение биологической продукции и видового разнообразия, возможно ухудшение условий среды и сооттветственно снижение биологической продукции и видового богатства.
Самой «элегантной» математической модели «хищник – жертва» отказались «подчиняться» большинство пар хищников и жертв в реальных экосистемах. Во-первых, хищники, как правило, переключаются на потребление других жертв, что не предусмотрено моделью. Во-вторых, на динамику численности популяций хищников и жертв действует множество других факторов, которые не учтены моделью (паразиты, биологические ритмы и т.д.).
«Число Линдемана» (10%) оказалось слишком приблизительным выражением эффективности перехода энергии с одного трофического уровня на другой. Если такая эффективность наблюдается в звене «растение – фитофаг», то на высших трофических уровнях она может превышать 50%.
Принцип конкурентного исключения, согласно которому в одной экологической нише не могут сосуществовать два вида, также оказался не универсальным. Во многих случаях при наличии сдерживающего фактора (нарушение, абиотический стресс, влияния конкурента и т.д.) виды могут не расходиться по разным нишам. Кроме того, несколько видов могут поочередно занимать одну и ту же нишу.
Далекой от реальности оказалась и модель формирования биоразнообразия на островах. Разные виды имеют разные шансы попасть на разные острова или быть вытесненными из их экосистем.
И так далее.
Не оправдались надежды и на математическое моделирование, которое Р. Макинтош (McIntosh, 1985) назвал «браком экологии и инженерии под дулом пистолета». Как отмечают в.Н. Тутубалин и др. (1999), построенные модели либо «… просто неадекватны и практически бесполезны для развития теоретической биологии (в том числе экологии, Б.М. и Л.Н.), либо полученные с их помощью биологические результаты в достаточной мере тривиальны и были получены и без их применения…» (с. 11). «Всемогущие» компьютеры в ряде случаев также не улучшили, а ухудшили ситуацию, так как стали, по образному выражению Р. Маргалефа (1992), причиной появления «компьютерного опиума в экологии», то есть утери экологами интуиции при оценке экологических закономерностей природы и очевидной переоценки возможностей их математического описания. Все это вело к созданию видимости «большой науки» там, где ее нет.
Стало очевидным, что разнообразие биологических объектов, как популяционного, так и экосистемного уровня, столь велико, что крайне сложно отыскать всеобщие законы, которые бы объясняли происходящие процессы, пространственные закономерности и позволяли прогнозировать их. Для сужения сферы их действия (определения областей экстраполяции) экологам потребовалось ввести понятия биологического пространства и биологического времени.
Первое измерение связано с размером особей и их подвижностью (не может быть одно пространство у тли, зайца и слона), второе – с продолжительностью жизненного цикла (он также различается у планктонной зеленой водоросли, ветвистоусого рачка, плотвы и сома).
Такое многоуровневое биологическое шкалирование сделало представления экологов более реалистическими. Стало ясно, что популяция или фрагмент сообщества, стабильные в одном масштабе, могут быть нестабильными в другом масштабе (нередко стабильность в крупном масштабе является суммой нестабильностей в мелком масштабе). При этом в последние годы усиливаются подходы «макроэкологии» (Brown, Maurer, 1989), т.е. анализа экологических закономерностей в глобальном масштабе, при котором погашаются «шумы», мешающие выявлению наиболее существенных закономерностей.
Для «новых» экологов экологический мир оказался сложно организованной иерархией в осях биологического пространства и биологического времени, стохастичным, с очень «мягкими» связями между особями и популяциями, динамичным, постоянно отклоняющимся от состояния экологического равновесия.
Таким образом, к концу ХХ в. стала очевидной сложность создания системы «универсальных законов» экологии, и родилась новая «универсальная методология». Внимание исследователей переключилось на изучение более частных пространственных и временных закономерностей, «механизмов» организации популяций и экосистем. Для выполнения этих исследований имелись все необходимые предпосылки: в экологии уже сформировался развитый понятийный аппарат и был накоплен огромный массив эмпирических данных, которые можно сопоставлять с новыми материалами и вовлекать в повторную обработку часто с изменением ранее сформулированных выводов.
Экология стала «полиморфной» (McIntosh, 1980) и «гетерогенной» (Ghilarov, 2001). В ней сохраняется тот плюрализм взглядов и, соответственно, методов исследования, который сформировался к середине ХХ в.: развиваются аутэкологический, популяционный и экосистемный подходы.
«По-видимому, экологическая теория сможет охватить существующую в природе реальность, только когда перестанет трактовать громадное разнообразие ситуаций, в которых оказываются организмы, популяции, сообщества и экосистемы, как некий «шум», мешающий выявлению наиболее существенных закономерностей, и будет рассматривать его как основной предмет своего изучения и важнейший источник информации» (Гиляров, 1998б, с. 82).
2. Расскажите о понятиях «биологическое пространство» и «биологическое время».
3. Как Вы понимаете «универсальную методологию экологии»?
2. «Золотой век» теории экологии.
3. Современная экология: крушение надежд на создание точной науки.
Часть 1. Аутэкология
Глава 2. Факторы среды
2.1. Классификация факторов среды
2.2. Условия и ресурсы
2.2.1. Ресурсы
Принцип экологической индивидуальности видов и их независимого распределения по градиентам среды в соответствии с симметричной колоколовидной кривой оказался не соответствующим действительности. В ряде случаев кривые распределения были асимметричными и даже полимодальными, что свидетельствовало о зависимости распределений видов друг от друга.
Как оказалось, продуктивными и богатыми видами могут быть не только климаксовые, устойчивые, но и сообщества, которые меняются в направлении климакса. Конвергенция всего разнообразия экосистем природного района в одну климаксовую экосистему просто невозможна, в каждом природном районе есть несколько климаксовых экосистем. Сукцессии, ведущие к климаксу, оказались стохастическими, а не жестко детерминированными процессами, в ходе которых виды сменяют друг друга в четкой последовательности. В ходе сукцессии не обязательно происходит улучшение условий, повышение биологической продукции и видового разнообразия, возможно ухудшение условий среды и сооттветственно снижение биологической продукции и видового богатства.
Самой «элегантной» математической модели «хищник – жертва» отказались «подчиняться» большинство пар хищников и жертв в реальных экосистемах. Во-первых, хищники, как правило, переключаются на потребление других жертв, что не предусмотрено моделью. Во-вторых, на динамику численности популяций хищников и жертв действует множество других факторов, которые не учтены моделью (паразиты, биологические ритмы и т.д.).
«Число Линдемана» (10%) оказалось слишком приблизительным выражением эффективности перехода энергии с одного трофического уровня на другой. Если такая эффективность наблюдается в звене «растение – фитофаг», то на высших трофических уровнях она может превышать 50%.
Принцип конкурентного исключения, согласно которому в одной экологической нише не могут сосуществовать два вида, также оказался не универсальным. Во многих случаях при наличии сдерживающего фактора (нарушение, абиотический стресс, влияния конкурента и т.д.) виды могут не расходиться по разным нишам. Кроме того, несколько видов могут поочередно занимать одну и ту же нишу.
Далекой от реальности оказалась и модель формирования биоразнообразия на островах. Разные виды имеют разные шансы попасть на разные острова или быть вытесненными из их экосистем.
И так далее.
Не оправдались надежды и на математическое моделирование, которое Р. Макинтош (McIntosh, 1985) назвал «браком экологии и инженерии под дулом пистолета». Как отмечают в.Н. Тутубалин и др. (1999), построенные модели либо «… просто неадекватны и практически бесполезны для развития теоретической биологии (в том числе экологии, Б.М. и Л.Н.), либо полученные с их помощью биологические результаты в достаточной мере тривиальны и были получены и без их применения…» (с. 11). «Всемогущие» компьютеры в ряде случаев также не улучшили, а ухудшили ситуацию, так как стали, по образному выражению Р. Маргалефа (1992), причиной появления «компьютерного опиума в экологии», то есть утери экологами интуиции при оценке экологических закономерностей природы и очевидной переоценки возможностей их математического описания. Все это вело к созданию видимости «большой науки» там, где ее нет.
Стало очевидным, что разнообразие биологических объектов, как популяционного, так и экосистемного уровня, столь велико, что крайне сложно отыскать всеобщие законы, которые бы объясняли происходящие процессы, пространственные закономерности и позволяли прогнозировать их. Для сужения сферы их действия (определения областей экстраполяции) экологам потребовалось ввести понятия биологического пространства и биологического времени.
Первое измерение связано с размером особей и их подвижностью (не может быть одно пространство у тли, зайца и слона), второе – с продолжительностью жизненного цикла (он также различается у планктонной зеленой водоросли, ветвистоусого рачка, плотвы и сома).
Такое многоуровневое биологическое шкалирование сделало представления экологов более реалистическими. Стало ясно, что популяция или фрагмент сообщества, стабильные в одном масштабе, могут быть нестабильными в другом масштабе (нередко стабильность в крупном масштабе является суммой нестабильностей в мелком масштабе). При этом в последние годы усиливаются подходы «макроэкологии» (Brown, Maurer, 1989), т.е. анализа экологических закономерностей в глобальном масштабе, при котором погашаются «шумы», мешающие выявлению наиболее существенных закономерностей.
Для «новых» экологов экологический мир оказался сложно организованной иерархией в осях биологического пространства и биологического времени, стохастичным, с очень «мягкими» связями между особями и популяциями, динамичным, постоянно отклоняющимся от состояния экологического равновесия.
Таким образом, к концу ХХ в. стала очевидной сложность создания системы «универсальных законов» экологии, и родилась новая «универсальная методология». Внимание исследователей переключилось на изучение более частных пространственных и временных закономерностей, «механизмов» организации популяций и экосистем. Для выполнения этих исследований имелись все необходимые предпосылки: в экологии уже сформировался развитый понятийный аппарат и был накоплен огромный массив эмпирических данных, которые можно сопоставлять с новыми материалами и вовлекать в повторную обработку часто с изменением ранее сформулированных выводов.
Экология стала «полиморфной» (McIntosh, 1980) и «гетерогенной» (Ghilarov, 2001). В ней сохраняется тот плюрализм взглядов и, соответственно, методов исследования, который сформировался к середине ХХ в.: развиваются аутэкологический, популяционный и экосистемный подходы.
«По-видимому, экологическая теория сможет охватить существующую в природе реальность, только когда перестанет трактовать громадное разнообразие ситуаций, в которых оказываются организмы, популяции, сообщества и экосистемы, как некий «шум», мешающий выявлению наиболее существенных закономерностей, и будет рассматривать его как основной предмет своего изучения и важнейший источник информации» (Гиляров, 1998б, с. 82).
Контрольные вопросы
1. В чем заключается основное отличие представлений современных экологов от взглядов ученых периода «золотого века экологии»?2. Расскажите о понятиях «биологическое пространство» и «биологическое время».
3. Как Вы понимаете «универсальную методологию экологии»?
Темы докладов на семинарских занятиях
1. Предтечи экологии и их вклад в развитие науки.2. «Золотой век» теории экологии.
3. Современная экология: крушение надежд на создание точной науки.
Часть 1. Аутэкология
Аутэкология, изучающая отношения организмов к условиям среды, – наиболее старый раздел общей экологии. По существу как аутэкологию понимал экологию Э. Геккель. Аутэкологом был и Ч. Дарвин – автор теории приспособления организмов к условиям среды путем естественного отбора.
В состав этого раздела экологии входят характеристика факторов среды (факториальная экология) и способов приспособления (адаптаций) организмов к различным ее условиям. В ХХ в. аутэкология пополнилась новыми разделами о функциональной роли организмов в экосистеме и их жизненных стратегиях.
Аутэкология исследует отношения организмов к условиям среды на уровне видов, что необходимо как для изучения популяций (это позволяет вынести «за скобки» те признаки, которые характерны для всех популяций одного вида), так и для изучения экосистем, элементами которых являются виды.
В состав этого раздела экологии входят характеристика факторов среды (факториальная экология) и способов приспособления (адаптаций) организмов к различным ее условиям. В ХХ в. аутэкология пополнилась новыми разделами о функциональной роли организмов в экосистеме и их жизненных стратегиях.
Аутэкология исследует отношения организмов к условиям среды на уровне видов, что необходимо как для изучения популяций (это позволяет вынести «за скобки» те признаки, которые характерны для всех популяций одного вида), так и для изучения экосистем, элементами которых являются виды.
Глава 2. Факторы среды
Факторы среды исследуются разными науками: климат – климатологией, рельеф – геоморфологией, почвы – почвоведением, закономерности распределения вод и их качество – гидрологией и гидрохимией и т.д. Однако экология изучает факторы среды не сами по себе, а их влияние на организмы, т.е. предлагает как бы взглянуть на каждый фактор «глазами» разных организмов.
2.1. Классификация факторов среды
Факторы среды делятся на абиотические, то есть факторы неорганической, или неживой, природы, и биотические – порожденные жизнедеятельностью организмов.
Совокупность абиотических факторов в пределах однородного участка называется экотопом, вся совокупность факторов, включая биотические, – биотопом.
К абиотическим факторам относятся:
1. климатические – свет, тепло, воздух, вода (включая осадки в различных формах и влажность воздуха), ветер;
2. эдафические, или почвенно-грунтовые, – механический и химический состав почвы, ее водный и температурный режим;
3. топографические – условия рельефа.
Климатические и эдафические факторы во многом определяются географическим положением экотопа – его удаленностью от экватора и от океана и высотой над уровнем моря.
Специфические абиотические факторы в водных экосистемах – глубина водной толщи, характер грунта на дне водоема, химический состав, прозрачность и температура воды, течение (или волны).
Абиотические факторы разделяются на прямые и косвенные.
Прямые факторы непосредственно влияют на организмы. Их примеры: влажность почвы и воздуха, температура, свет, богатство почвы и воды элементами минерального питания, скорость течения воды и др.
Косвенные факторы действуют на организмы опосредствованно – через прямые факторы. Их примеры: географическая широта и удаленность от океана, рельеф (высота над уровнем моря и экспозиция склона), гранулометрический состав почвы, прозрачность воды.
По градиенту географической широты с севера на юг в силу изменения угла падения солнечного света возрастает количество энергии Солнца, поступающей на единицу поверхности Земли, при удалении от океана – падает количество осадков. Эти закономерности известны с конца XVIII столетия, а в начале ХХ столетия стали популярными схемы «идеальных материков» с координатами «расстояние от экватора/расстояние от океана» или среднегодовая температура и среднегодовое количество осадков (рис. 1). На «идеальном материке» показано распределение биомов – крупных экологических вариантов экосистем (см. 11.7). Для европейской части России была построена профильная схема изменения экологических факторов от Северного ледовитого океана до зоны пустыни (рис. 2).
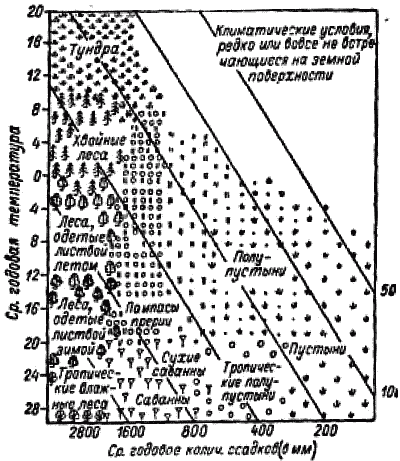 Рис. 1. Схема распределения биомов на «идеальном континенте» в зависимости от основных климатических параметров (по Волобуеву, 1956).
Рис. 1. Схема распределения биомов на «идеальном континенте» в зависимости от основных климатических параметров (по Волобуеву, 1956).
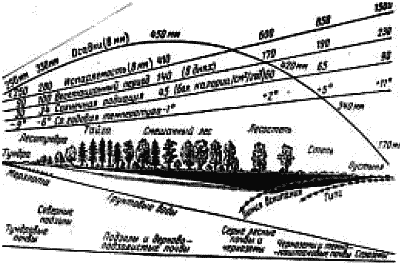 Рис. 2. Схема изменения основных экологических факторов на профиле «тундра – пустыня» в европейской части России.
Рис. 2. Схема изменения основных экологических факторов на профиле «тундра – пустыня» в европейской части России.
С подъемом в горы также изменяется климат (количество осадков и температурный режим); экспозиция и крутизна склона влияют на интенсивность прогревания поверхности почвы и режим ее увлажнения. В среднем с повышением высоты над уровнем моря на 100 м среднегодовая температура уменьшается на 0,5°C, однако изменения климата на этом градиенте зависят от географической широты и удаленности от океана: широтный, долготный и высотный градиенты взаимодействуют.
Так нижняя граница леса в южных районах (Кавказ, Тянь-Шань) проходит на высоте около 2000 м, а в лесной зоне средней полосы лесной пояс начинается «от нуля». Велики различия климата, связанные с экспозицией, от которой зависит интенсивность поступления на поверхность солнечной энергии. Климатические пояса и соответствующие им пояса растительности всегда «наклонены» на север. В степной зоне пояс леса на северных склонах спускается на 100–200 м ниже, чем на южных, а в зоне пустынь по южным склонам леса нет вообще.
В горах Санта-Каталина Р. Уиттекером (1980) было изучено распределение экосистем в зависимости от высоты над уровнем моря и экспозиции («градиента топографического увлажнения»). По хорошо прогреваемым южным склонам все типы экосистем поднимаются выше, чем по холодным северным (рис. 3).
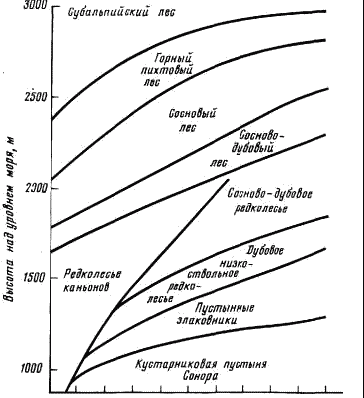 Рис. 3. Распределение основных типов экосистем в зависимости от высоты над уровнем моря и экспозиции в горах Санта-Котолина (по Уиттекеру, 1980).
Рис. 3. Распределение основных типов экосистем в зависимости от высоты над уровнем моря и экспозиции в горах Санта-Котолина (по Уиттекеру, 1980).
Гранулометрический состав почвы влияет на растения и почвенную фауну через режим увлажнения и динамику питательных элементов.
Биотические факторы являются следствием взаимоотношений организмов. Для растений – это конкуренция, влияние животных (фитофаги, паразиты, опылители, распространители плодов и семян), грибов (микоризные, паразитические), бактерий (азотфиксирующие и болезнетворные), вирусов. Для животных – это конкуренция, влияние хищников, патогенных микроорганизмов, растений (для фитофагов).
Факторы, связанные с влиянием человека, выделяются в отдельную группу антропогенных. К наиболее существенным антропогенным факторам относятся следующие: химическое загрязнение воды, атмосферы и почвы, техногенное нарушение экосистем при разработке полезных ископаемых, выпас скота, рекреационное влияние, промысел животных (включая лов рыбы), заготовка растительного сырья. Особую роль человек играет как агент переселения видов из одного района в другой. Биологические инвазии, спровоцированные человеком, в настоящее время приняли катастрофические масштабы (см. 12.9).
В настоящее время роль антропогенных факторов резко возросла, и потому изучение последствий их влияния и разработка способов регулирования отношений человека и природы являются важнейшими проблемами прикладной экологии (инвайронменталистики).
2. Расскажите о косвенных экологических факторах.
3. Дайте общую характеристику антропогенным факторам.
Совокупность абиотических факторов в пределах однородного участка называется экотопом, вся совокупность факторов, включая биотические, – биотопом.
К абиотическим факторам относятся:
1. климатические – свет, тепло, воздух, вода (включая осадки в различных формах и влажность воздуха), ветер;
2. эдафические, или почвенно-грунтовые, – механический и химический состав почвы, ее водный и температурный режим;
3. топографические – условия рельефа.
Климатические и эдафические факторы во многом определяются географическим положением экотопа – его удаленностью от экватора и от океана и высотой над уровнем моря.
Специфические абиотические факторы в водных экосистемах – глубина водной толщи, характер грунта на дне водоема, химический состав, прозрачность и температура воды, течение (или волны).
Абиотические факторы разделяются на прямые и косвенные.
Прямые факторы непосредственно влияют на организмы. Их примеры: влажность почвы и воздуха, температура, свет, богатство почвы и воды элементами минерального питания, скорость течения воды и др.
Косвенные факторы действуют на организмы опосредствованно – через прямые факторы. Их примеры: географическая широта и удаленность от океана, рельеф (высота над уровнем моря и экспозиция склона), гранулометрический состав почвы, прозрачность воды.
По градиенту географической широты с севера на юг в силу изменения угла падения солнечного света возрастает количество энергии Солнца, поступающей на единицу поверхности Земли, при удалении от океана – падает количество осадков. Эти закономерности известны с конца XVIII столетия, а в начале ХХ столетия стали популярными схемы «идеальных материков» с координатами «расстояние от экватора/расстояние от океана» или среднегодовая температура и среднегодовое количество осадков (рис. 1). На «идеальном материке» показано распределение биомов – крупных экологических вариантов экосистем (см. 11.7). Для европейской части России была построена профильная схема изменения экологических факторов от Северного ледовитого океана до зоны пустыни (рис. 2).
С подъемом в горы также изменяется климат (количество осадков и температурный режим); экспозиция и крутизна склона влияют на интенсивность прогревания поверхности почвы и режим ее увлажнения. В среднем с повышением высоты над уровнем моря на 100 м среднегодовая температура уменьшается на 0,5°C, однако изменения климата на этом градиенте зависят от географической широты и удаленности от океана: широтный, долготный и высотный градиенты взаимодействуют.
Так нижняя граница леса в южных районах (Кавказ, Тянь-Шань) проходит на высоте около 2000 м, а в лесной зоне средней полосы лесной пояс начинается «от нуля». Велики различия климата, связанные с экспозицией, от которой зависит интенсивность поступления на поверхность солнечной энергии. Климатические пояса и соответствующие им пояса растительности всегда «наклонены» на север. В степной зоне пояс леса на северных склонах спускается на 100–200 м ниже, чем на южных, а в зоне пустынь по южным склонам леса нет вообще.
В горах Санта-Каталина Р. Уиттекером (1980) было изучено распределение экосистем в зависимости от высоты над уровнем моря и экспозиции («градиента топографического увлажнения»). По хорошо прогреваемым южным склонам все типы экосистем поднимаются выше, чем по холодным северным (рис. 3).
Гранулометрический состав почвы влияет на растения и почвенную фауну через режим увлажнения и динамику питательных элементов.
Биотические факторы являются следствием взаимоотношений организмов. Для растений – это конкуренция, влияние животных (фитофаги, паразиты, опылители, распространители плодов и семян), грибов (микоризные, паразитические), бактерий (азотфиксирующие и болезнетворные), вирусов. Для животных – это конкуренция, влияние хищников, патогенных микроорганизмов, растений (для фитофагов).
Факторы, связанные с влиянием человека, выделяются в отдельную группу антропогенных. К наиболее существенным антропогенным факторам относятся следующие: химическое загрязнение воды, атмосферы и почвы, техногенное нарушение экосистем при разработке полезных ископаемых, выпас скота, рекреационное влияние, промысел животных (включая лов рыбы), заготовка растительного сырья. Особую роль человек играет как агент переселения видов из одного района в другой. Биологические инвазии, спровоцированные человеком, в настоящее время приняли катастрофические масштабы (см. 12.9).
В настоящее время роль антропогенных факторов резко возросла, и потому изучение последствий их влияния и разработка способов регулирования отношений человека и природы являются важнейшими проблемами прикладной экологии (инвайронменталистики).
Контрольные вопросы
1. Перечислите основные абиотические факторы среды.2. Расскажите о косвенных экологических факторах.
3. Дайте общую характеристику антропогенным факторам.
2.2. Условия и ресурсы
Прямые абиотические факторы подразделяются на факторы-условия и факторы-ресурсы.
По М. Бигону и др. (1989), условия – это изменяющиеся во времени и пространстве факторы среды обитания, на которые организмы реагируют по-разному, но эти составляющие среды не расходуются: один организм не может сделать их более доступными или недоступными для других. К числу факторов-условий относятся температура, влажность воздуха, соленость воды и скорость ее течения, реакция (рН) почвенного раствора, содержание в воде и почве загрязняющих веществ, которые не используются растениями как элементы питания.
В отличие от факторов-условий, факторы-ресурсы расходуются организмами в процессе жизнедеятельности, и потому один более сильный организм может «съесть» ресурсов больше, а другому, более слабому, их останется меньше.
По М. Бигону и др. (1989), условия – это изменяющиеся во времени и пространстве факторы среды обитания, на которые организмы реагируют по-разному, но эти составляющие среды не расходуются: один организм не может сделать их более доступными или недоступными для других. К числу факторов-условий относятся температура, влажность воздуха, соленость воды и скорость ее течения, реакция (рН) почвенного раствора, содержание в воде и почве загрязняющих веществ, которые не используются растениями как элементы питания.
В отличие от факторов-условий, факторы-ресурсы расходуются организмами в процессе жизнедеятельности, и потому один более сильный организм может «съесть» ресурсов больше, а другому, более слабому, их останется меньше.
2.2.1. Ресурсы
Для растений ресурсами являются свет, вода, элементы минерального питания, диоксид углерода, для насекомоопыляемых – насекомые-опылители (ветер как опылитель является фактором-условием). Для животных-фитофагов ресурсом являются растения, для зоофагов (хищников) – живые животные, для детритофагов-сапротрофов и редуцентов (бактерии, грибы) – мертвое органическое вещество. Для большинства организмов необходимым ресурсом является кислород.
Свет. Это основной источник энергии для наземных и водных экосистем. При этом из всех щедрот солнечной энергии, поступающей на Землю, на фотосинтез расходуется сравнительно небольшая часть света. Только культуре микроскопических морских водорослей удалось достичь использования для целей фотосинтеза 4,5%. В наземных экосистемах усвоение солнечной энергии для фотосинтеза не превышает 1–3% (тропические леса) и составляет в лесах умеренных широт 0,6–1,2%, а в посевах сельскохозяйственных культур даже с наиболее плотным пологом растений он не выше 0,6%.
Все экосистемы Земли используют в процессе фотосинтеза не более 0,001% от всего потока энергии, поступающей с солнечным светом на Землю. В 30–40 раз больше растения используют тепловой энергии солнечного света на испарение (транспирацию). В результате транспирации через корни, стебли и листья растений прогоняется раствор элементов питания, необходимый для их жизнедеятельности. Кроме того, это спасает растения от перегрева.
Для фотосинтеза используется лишь часть световых волн – в диапазоне 400–700 нм. Эта часть солнечной энергии составляет около 40% поступающего на Землю света и называется фотосинтетически активной радиацией (ФАР). Наибольшее значение в составе ФАР имеют оранжево-красные и сине-фиолетовые лучи. При прохождении через большую толщу воды эти части света отфильтровываются, и до глубоких слоев доходят в основном зеленые лучи. Однако если эти лучи плохо усваиваются зелеными растениями, то за счет дополнительных пигментов их могут использовать красные водоросли (Rhodophyta). Бактерии-фототрофы также используют часть света, но с иным диапазоном длины волны – 800–900 нм.
Во многих случаях количество света избыточно, и потому интенсивность фотосинтеза не лимитируется поступающей солнечной энергией. Дефицит света наблюдается в затененных местообитаниях, например под густым пологом древостоя. В таких условиях у теневыносливых растений (сциофитов) выражен специальный синдром признаков теневыносливости, позволяющий усвоить больше света (тонкие листья, высокое содержание хлорофилла). Типичные сциофиты – папоротники, обитающие в расщелинах скал, например, листовика (Phyllitis scolopendrium).
Эффективность усвоения света в сообществах растений повышается за счет специальных приспособлений: вертикальное расположение листьев злаков, использующих свет, падающий на лист под острым углом (луга и степи); многослойная крона листьев (леса). Показателем числа слоев листьев, через которые проходит свет, является индекс листовой поверхности (ИЛП), который определяется как отношение площади листьев к площади поверхности почвы, над которой они находятся. В разомкнутых сообществах пустынь ИЛП составляет доли единицы, в большинстве луговых сообществ – равен 4–6, а в еловом лесу – может достигать 12, то есть на 1 гектар леса приходится 12 гектаров поверхности листьев (Работнов, 1992).
Экологическое значение имеют и невидимые лучи, т.е. не воспринимаемые глазом человека. Так самые короткие ультрафиолетовые лучи при высокой интенсивности ослабляют иммунную систему животных, в особенности человека, при умеренной интенсивности они способствуют образованию витамина D в животных организмах. Инфракрасные (тепловые) лучи влияют на температурный режим теплокровных животных, при повышении их интенсивности снижается активность окислительных процессов.
Свет является неисчерпаемым ресурсом, который постоянно поступает на Землю в результате солнечной радиации.
Вода. Необходимым фактором жизни любого организма является его обводнение, так как именно вода является средой, в которой протекают все основные метаболические процессы. Ни один организм не обладает надежной системой сохранения воды, содержащейся в его клетках, и потому этот ресурс нуждается в постоянном пополнении. Вода – важнейший ресурс, участвующий в фотосинтезе, хотя основная ее часть, которая всасывается корнями растений, расходуется на испарение, что связано во многом с процессом поглощения через устьица диоксида углерода для фотосинтеза (мембран, которые способны «впускать» углекислый газ и «не выпускать» воду, нет).
Специальные приспособления характерны для растений, обитающих в условиях дефицита влаги (см. 4.4.3), и растений избыточно увлажненных местообитаний (у водных растений проводящая система замещена воздухоносной тканью – аэренхимой).
Нет необходимости говорить о различиях водных и наземных животных. Среди наземных животных существуют виды с разной потребностью в воде. Так животные пустыни, где постоянно ощущается дефицит воды, значительное количество воды получают при разложении жиров, которые выступают запасниками потенциальной влаги. «Депо» жиров у тушканчиков, песчанок находится в хвосте, у верблюда – в горбе.
Степень доступности воды накладывает ограничения на распространение многих видов животных и на потребление ими других ресурсов. Не только домашний скот, но и дикие копытные животные могут разрушить травостой в результате перевыпаса только близ водопоев, хотя радиусы пастьбы у них значительно больше, чем у коров или овец. По этой причине отдаленные от воды участки злаковника или саванны оказываются лучше сохранившимися.
Круговорот воды в биосфере делает ее неисчерпаемым (возобновимым) ресурсом, однако под влиянием человека этот круговорот изменился (см. 13.2.2). Кроме того, во многих районах вода сильно загрязнена, что ограничивает возможность использования ее организмами многих видов, включая человека.
Диоксид углерода. Этот ресурс необходим для фотосинтеза, но его содержание в атмосфере столь велико, что в естественных условиях он не лимитирует процесс синтеза органического вещества. Аналогично не лимитирует интенсивность фотосинтеза водных растений содержание диоксида углерода в воде.
Диоксид углерода является не только прямым фактором-ресурсом, но и косвенным фактором, влияющим на климат. В результате сжигания больших количеств топлива, содержащего углерод, концентрация диоксида углерода в атмосфере повышается. В итоге происходит потепление климата (см. 13.2.1).
Элементы питания. Элементы, необходимые для жизни организмов, называются биогенными. Из 54 элементов периодической таблицы, которые встречаются в природе, около половины их важны либо для животных, либо для растений. Основные биогены называются макроэлементами, шесть из них нужны всем живым существам и в больших количествах. Чтобы запомнить их, экологи составили из латинских букв, соответствующих химическим символам, смешное слово CHNOPS («ЧНОПС»: С – углерод, Н – водород, N – азот, О – кислород, Р – фосфор, S – сера).
Из других макроэлементов важны: кальций, калий, магний, причем кальций в больших количествах необходим позвоночным и моллюскам для построения скелета или раковин, а магний – растениям, так как он входит в состав молекулы хлорофилла.
Остальные элементы нужны организмам в меньших количествах и называются микроэлементами. Растениям необходимы 10 микроэлементов, в том числе для фотосинтеза – марганец, железо, хлор, цинк, ванадий; для азотного обмена – молибден, бор, кобальт, железо; для прочих метаболических реакций – марганец, бор, кобальт, медь, кремний. Все эти элементы, кроме бора, нужны и животным. Кроме того, животным необходимы селен, хром, никель, йод, фтор, олово, мышьяк.
У разных растений отмечаются свои «пристрастия» к микроэлементам. Так некоторым папоротникам для нормального развития необходим алюминий, диатомовым водорослям – кремний, а некоторым зеленым водорослям – селен. Для успешного симбиоза азотфиксирующих бактерий и бобовых (см. 8.6) необходим кобальт.
Кислород. Этот элемент необходим для дыхания подавляющему большинству организмов, однако его дефицит наблюдается только в водных экосистемах и переувлажненных почвах, что связано с низкой растворимостью кислорода в воде. Если в 1 л воздуха содержится 210 см 3 кислорода, то в воде его содержание не превышает 10 см 3 л, причем растворимость кислорода снижается при повышении температуры и солености. Это делает кислород фактором, ограничивающим возможности жизни многих обитателей водоемов. Они гибнут летом при повышении температуры и зимой при заморозк, когда вода изолирована от атмосферы слоем льда и весь кислород израсходован организмами.
Пополнение запаса кислорода в воде происходит за счет его поступления из воздуха, причем этот медленный процесс может ускорить сильный ветер. Кислород выделяют водные растения, в первую очередь фитопланктон, в процессе фотосинтеза. По этой причине содержание кислорода связано с количеством света, который проникает в водную толщу, что в свою очередь зависит от прозрачности воды. Поэтому, чем вода прозрачнее, тем выше в ней содержание кислорода. Все это объясняет сложную динамику содержания кислорода в воде в зависимости от типа водоема, времени суток и времени года.
В подтапливаемых почвах, т.е. с близким уровнем грунтовых вод, корни древесных растений избегают глубоких пересыщенных водой горизонтов. В зоне дефицита кислорода они практически не всасывают воду и растворенные в ней элементы минерального питания.
Пространство. Физическое пространство является ресурсом, потому что любые факторы-ресурсы, которые потребляются организмами, занимают определенную территорию. Растения, чтобы проходить нормальный жизненный цикл, должны получить определенную площадь «под солнцем» и некоторый объем почвы для потребления воды и элементов минерального питания (площадь питания). Животным-фитофагам нужен «участок пастбища» (для тли это будет часть листа, для косяка лошадей – десятки гектаров степи, для стада слонов – десятки квадратных километров), плотоядным животным – охотничьи наделы.
Свет. Это основной источник энергии для наземных и водных экосистем. При этом из всех щедрот солнечной энергии, поступающей на Землю, на фотосинтез расходуется сравнительно небольшая часть света. Только культуре микроскопических морских водорослей удалось достичь использования для целей фотосинтеза 4,5%. В наземных экосистемах усвоение солнечной энергии для фотосинтеза не превышает 1–3% (тропические леса) и составляет в лесах умеренных широт 0,6–1,2%, а в посевах сельскохозяйственных культур даже с наиболее плотным пологом растений он не выше 0,6%.
Все экосистемы Земли используют в процессе фотосинтеза не более 0,001% от всего потока энергии, поступающей с солнечным светом на Землю. В 30–40 раз больше растения используют тепловой энергии солнечного света на испарение (транспирацию). В результате транспирации через корни, стебли и листья растений прогоняется раствор элементов питания, необходимый для их жизнедеятельности. Кроме того, это спасает растения от перегрева.
Для фотосинтеза используется лишь часть световых волн – в диапазоне 400–700 нм. Эта часть солнечной энергии составляет около 40% поступающего на Землю света и называется фотосинтетически активной радиацией (ФАР). Наибольшее значение в составе ФАР имеют оранжево-красные и сине-фиолетовые лучи. При прохождении через большую толщу воды эти части света отфильтровываются, и до глубоких слоев доходят в основном зеленые лучи. Однако если эти лучи плохо усваиваются зелеными растениями, то за счет дополнительных пигментов их могут использовать красные водоросли (Rhodophyta). Бактерии-фототрофы также используют часть света, но с иным диапазоном длины волны – 800–900 нм.
Во многих случаях количество света избыточно, и потому интенсивность фотосинтеза не лимитируется поступающей солнечной энергией. Дефицит света наблюдается в затененных местообитаниях, например под густым пологом древостоя. В таких условиях у теневыносливых растений (сциофитов) выражен специальный синдром признаков теневыносливости, позволяющий усвоить больше света (тонкие листья, высокое содержание хлорофилла). Типичные сциофиты – папоротники, обитающие в расщелинах скал, например, листовика (Phyllitis scolopendrium).
Эффективность усвоения света в сообществах растений повышается за счет специальных приспособлений: вертикальное расположение листьев злаков, использующих свет, падающий на лист под острым углом (луга и степи); многослойная крона листьев (леса). Показателем числа слоев листьев, через которые проходит свет, является индекс листовой поверхности (ИЛП), который определяется как отношение площади листьев к площади поверхности почвы, над которой они находятся. В разомкнутых сообществах пустынь ИЛП составляет доли единицы, в большинстве луговых сообществ – равен 4–6, а в еловом лесу – может достигать 12, то есть на 1 гектар леса приходится 12 гектаров поверхности листьев (Работнов, 1992).
Экологическое значение имеют и невидимые лучи, т.е. не воспринимаемые глазом человека. Так самые короткие ультрафиолетовые лучи при высокой интенсивности ослабляют иммунную систему животных, в особенности человека, при умеренной интенсивности они способствуют образованию витамина D в животных организмах. Инфракрасные (тепловые) лучи влияют на температурный режим теплокровных животных, при повышении их интенсивности снижается активность окислительных процессов.
Свет является неисчерпаемым ресурсом, который постоянно поступает на Землю в результате солнечной радиации.
Вода. Необходимым фактором жизни любого организма является его обводнение, так как именно вода является средой, в которой протекают все основные метаболические процессы. Ни один организм не обладает надежной системой сохранения воды, содержащейся в его клетках, и потому этот ресурс нуждается в постоянном пополнении. Вода – важнейший ресурс, участвующий в фотосинтезе, хотя основная ее часть, которая всасывается корнями растений, расходуется на испарение, что связано во многом с процессом поглощения через устьица диоксида углерода для фотосинтеза (мембран, которые способны «впускать» углекислый газ и «не выпускать» воду, нет).
Специальные приспособления характерны для растений, обитающих в условиях дефицита влаги (см. 4.4.3), и растений избыточно увлажненных местообитаний (у водных растений проводящая система замещена воздухоносной тканью – аэренхимой).
Нет необходимости говорить о различиях водных и наземных животных. Среди наземных животных существуют виды с разной потребностью в воде. Так животные пустыни, где постоянно ощущается дефицит воды, значительное количество воды получают при разложении жиров, которые выступают запасниками потенциальной влаги. «Депо» жиров у тушканчиков, песчанок находится в хвосте, у верблюда – в горбе.
Степень доступности воды накладывает ограничения на распространение многих видов животных и на потребление ими других ресурсов. Не только домашний скот, но и дикие копытные животные могут разрушить травостой в результате перевыпаса только близ водопоев, хотя радиусы пастьбы у них значительно больше, чем у коров или овец. По этой причине отдаленные от воды участки злаковника или саванны оказываются лучше сохранившимися.
Круговорот воды в биосфере делает ее неисчерпаемым (возобновимым) ресурсом, однако под влиянием человека этот круговорот изменился (см. 13.2.2). Кроме того, во многих районах вода сильно загрязнена, что ограничивает возможность использования ее организмами многих видов, включая человека.
Диоксид углерода. Этот ресурс необходим для фотосинтеза, но его содержание в атмосфере столь велико, что в естественных условиях он не лимитирует процесс синтеза органического вещества. Аналогично не лимитирует интенсивность фотосинтеза водных растений содержание диоксида углерода в воде.
Диоксид углерода является не только прямым фактором-ресурсом, но и косвенным фактором, влияющим на климат. В результате сжигания больших количеств топлива, содержащего углерод, концентрация диоксида углерода в атмосфере повышается. В итоге происходит потепление климата (см. 13.2.1).
Элементы питания. Элементы, необходимые для жизни организмов, называются биогенными. Из 54 элементов периодической таблицы, которые встречаются в природе, около половины их важны либо для животных, либо для растений. Основные биогены называются макроэлементами, шесть из них нужны всем живым существам и в больших количествах. Чтобы запомнить их, экологи составили из латинских букв, соответствующих химическим символам, смешное слово CHNOPS («ЧНОПС»: С – углерод, Н – водород, N – азот, О – кислород, Р – фосфор, S – сера).
Из других макроэлементов важны: кальций, калий, магний, причем кальций в больших количествах необходим позвоночным и моллюскам для построения скелета или раковин, а магний – растениям, так как он входит в состав молекулы хлорофилла.
Остальные элементы нужны организмам в меньших количествах и называются микроэлементами. Растениям необходимы 10 микроэлементов, в том числе для фотосинтеза – марганец, железо, хлор, цинк, ванадий; для азотного обмена – молибден, бор, кобальт, железо; для прочих метаболических реакций – марганец, бор, кобальт, медь, кремний. Все эти элементы, кроме бора, нужны и животным. Кроме того, животным необходимы селен, хром, никель, йод, фтор, олово, мышьяк.
У разных растений отмечаются свои «пристрастия» к микроэлементам. Так некоторым папоротникам для нормального развития необходим алюминий, диатомовым водорослям – кремний, а некоторым зеленым водорослям – селен. Для успешного симбиоза азотфиксирующих бактерий и бобовых (см. 8.6) необходим кобальт.
Кислород. Этот элемент необходим для дыхания подавляющему большинству организмов, однако его дефицит наблюдается только в водных экосистемах и переувлажненных почвах, что связано с низкой растворимостью кислорода в воде. Если в 1 л воздуха содержится 210 см 3 кислорода, то в воде его содержание не превышает 10 см 3 л, причем растворимость кислорода снижается при повышении температуры и солености. Это делает кислород фактором, ограничивающим возможности жизни многих обитателей водоемов. Они гибнут летом при повышении температуры и зимой при заморозк, когда вода изолирована от атмосферы слоем льда и весь кислород израсходован организмами.
Пополнение запаса кислорода в воде происходит за счет его поступления из воздуха, причем этот медленный процесс может ускорить сильный ветер. Кислород выделяют водные растения, в первую очередь фитопланктон, в процессе фотосинтеза. По этой причине содержание кислорода связано с количеством света, который проникает в водную толщу, что в свою очередь зависит от прозрачности воды. Поэтому, чем вода прозрачнее, тем выше в ней содержание кислорода. Все это объясняет сложную динамику содержания кислорода в воде в зависимости от типа водоема, времени суток и времени года.
В подтапливаемых почвах, т.е. с близким уровнем грунтовых вод, корни древесных растений избегают глубоких пересыщенных водой горизонтов. В зоне дефицита кислорода они практически не всасывают воду и растворенные в ней элементы минерального питания.
Пространство. Физическое пространство является ресурсом, потому что любые факторы-ресурсы, которые потребляются организмами, занимают определенную территорию. Растения, чтобы проходить нормальный жизненный цикл, должны получить определенную площадь «под солнцем» и некоторый объем почвы для потребления воды и элементов минерального питания (площадь питания). Животным-фитофагам нужен «участок пастбища» (для тли это будет часть листа, для косяка лошадей – десятки гектаров степи, для стада слонов – десятки квадратных километров), плотоядным животным – охотничьи наделы.