Наиболее активно поглощается водой красная часть спектра света, поэтому, как отмечалось, глубоководья морей заселены красными водорослями, способными за счет дополнительных пигментов усваивать зеленый свет. Прозрачность воды определяется несложным прибором – диском Секки, который представляет собой окрашенный в белый цвет круг диаметром 20 см. О степени прозрачности воды судят по глубине, на которой диск становится неразличимым.
Важнейшей характеристикой воды является ее химический состав – содержание солей (в том числе биогенов), газов, ионов водорода (рН). По концентрации биогенов, особенно фосфора и азота, водоемы разделяются на олиготрофные, мезотрофные и эвтрофные. При повышении содержания биогенов, скажем, при загрязнении водоема стоками, происходит процесс эвтрофикации водных экосистем (см. 12.7).
Содержание кислорода в воде примерно в 20 раз ниже, чем в атмосфере, и составляет 6–8 мл/л. Оно снижается при повышении температуры, а также в стоячих водоемах в зимнее время, когда вода изолирована от атмосферы слоем льда. Снижение концентрации кислорода может стать причиной гибели многих обитателей водных экосистем, исключая особо устойчивые к дефициту кислорода виды, подобные карасю или линю, которые могут жить даже при снижении содержания кислорода до 0,5 мл/л.
Содержание углекислого газа в воде, напротив, выше, чем в атмосфере. В морской воде его может содержаться до 40–50 мл/л, что примерно в 150 раз выше, чем в атмосфере. Потребление углекислого газа фитопланктоном при интенсивном фотосинтезе не превышает 0,5 мл/л в сутки.
Концентрация ионов водорода в воде (рН) может меняться в пределах 3,7–7,8. Нейтральными считаются воды с рН от 6,45 до 7,3. Как уже отмечалось, с понижением рН биоразнообразие организмов, населяющих водную среду, быстро убывает. Речной рак, многие виды моллюсков гибнут при рН ниже 6, окунь и щука могут выдержать рН до 5, угорь и голец выживают при понижении рН до 5–4,4. В более кислых водах сохраняются лишь некоторые виды зоопланктона и фитопланктона. Кислотные дожди, связанные с выбросами в атмосферу больших количеств оксидов серы и азота промышленными предприятиями, стали причиной подкисления вод озер Европы и США и резкого обеднения их биологического разнообразия.
Наземно-воздушая среда жизни. Воздух отличается значительно более низкой плотностью по сравнению с водой. По этой причине освоение воздушной среды, которое произошло много позже, чем зарождение жизни и ее развитие в водной среде, сопровождалось усилением развития механических тканей, которые позволили организмам противостоять действию закона всемирного тяготения и ветра (скелет у позвоночных животных, хитиновые панцири у насекомых, склеренхима у растений). В условиях только воздушной среды ни один организм постоянно жить не может, и потому даже лучшие «летуны» (птицы и насекомые) должны периодически опускаться на землю. Перемещение организмов по воздуху возможно за счет специальных приспособлений – крыльев у птиц, насекомых, некоторых видов млекопитающих и даже рыб, парашютики и крылышки у семян, воздушные мешки у пыльцы хвойных пород и т.д.
Воздух – плохой проводник тепла, и потому именно в воздушной среде на суше возникли эндотермные (теплокровные) животные, которым легче сохранить тепло, чем эктотермным обитателям водной среды. Для теплокровных водных животных, включая гигантов-китов, водная среда вторична, предки этих животных когда-то жили на суше.
Для жизни в воздушной среде потребовались более сложные механизмы размножения, которые исключали бы риск высыхания половых клеток (многоклеточные антеридии и архегонии, а затем семязачатки и завязи у растений, внутреннее оплодотворение у животных, яйца с плотной оболочкой у птиц, пресмыкающихся, земноводных и др.).
В целом возможностей для формирования разнообразных сочетаний факторов в условиях наземно-воздушной среды много больше, чем водной. Именно в этой среде особенно ярко проявляются различия климата разных районов (и на разных высотах над уровнем моря в пределах одного района). Поэтому разнообразие наземных организмов много выше, чем водных.
Почвенная среда жизни. Большая часть суши покрыта тонким слоем (по сравнению с толщей земной коры) почвы, названной в.И. Вернадским биокосным телом. Почва представляет собой сложный многослойный «пирог» из горизонтов с разными свойствами, причем состав и толщина «пирога» в разных зонах различны. Общеизвестны зональный (от подзолов и серых лесных до черноземов, каштановых и бурых почв) и гидрогенный (от влажно-луговых до болотно-торфянистых) ряды почв. В южных районах почвы могут быть, кроме того, засолены на поверхности (солончаковатые почвы и солончаки) или в глубине (солонцы).
Любая почва представляет собой многофазную систему, в состав которой входят:
– минеральные частицы – от тончайшего ила до песка и гравия;
– органическое вещество – от тел только что умерших животных и отмерших корней растений до гумуса, в котором это органическое вещество подверглось сложной химической обработке;
– газовая (воздушная) фаза, характер которой во многом определяется физическими свойствами почвы – ее структурой и соответственно плотностью и порозностью. Газовая фаза почвы всегда обогащена углекислым газом и парами воды и может быть обеднена кислородом, что сближает условия жизни в почве с условиями водной среды;
– водная фаза. Вода в почве также может содержаться в разных количествах (от избытка до крайнего дефицита) и в разных качествах, быть гравитационной, т.е. свободно перемещающейся по капиллярам и наиболее доступной для корней растений и животных организмов, гигроскопической, т.е. входящей в состав коллоидных частиц, и газовой, т.е. в форме пара.
Эта многофазность почв делает их среду наиболее насыщенной жизнью. В почвах сконцентрирована основная биомасса животных, бактерий, грибов, в ней расположены корни растений, живущих в наземно-воздушной среде, но извлекающих из почвы воду с элементами питания и поставляющие в «темный мир» почвы органическое вещество, накопленное в процессе фотосинтеза на свету. Почва – это главный «цех по переработке» органического вещества, через нее протекает до 90% углерода, возвращаемого в атмосферу.
Гигантское разнообразие жизни в почве включает не только те организмы, которые живут в ней постоянно – позвоночные (кроты), членистоногие, бактерии, водоросли, дождевые черви и т.д., но и те организмы, которые связаны с ней лишь в начале своей «биографии» (саранчовые, многие жуки и т.д.).
Адаптация растений к некоторым вариантам экстремальных почвенных условий (засуха, засоление) будут рассмотрены в следующей главе.
Организмы как среда жизни. Нет ни одного вида многоклеточных организмов, который не был бы заселен другими организмами, и в первую очередь паразитами. Разные организмы и разные их органы, ткани и клетки имеют свою специфику как жизненные среды и свое паразитарное население, которое в разной степени опасно для хозяина, предоставившего им «жилплощадь со столом». Тем не менее, есть общие особенности этой среды жизни: в ней смягчены колебания внешних условий, и практически не ограничены ресурсы пищи.
Далеко не всегда хозяин, который заселен паразитами, оказывается беспомощным перед «назойливыми квартирантами». В этом случае паразитам приходится преодолевать неблагоприятные условия, формируемые в результате защитных реакций хозяина. По этой причине лучшая среда жизни для многих паразитов – это старые ослабленные особи.
Организмы как среду жизни, кроме паразитов, могут использовать многие виды, которые полезны организму-хозяину, т.е. находятся с ним в отношениях взаимовыгодного сотрудничества – мутуализма (см. 8.6).
2. Как организмы адаптируются к жизни в водной среде?
3. Какие факторы определяют способность света «пробивать» водную толщу?
4. Как изменяется спектр солнечного света при прохождении через водную толщу?
5. Какие факторы являются лимитирующими в водной среде?
6. Приведите примеры влияния рН водной среды на организмы.
7. Расскажите об основных отличиях наземно-воздушной и водной сред жизни.
8. Какие изменения в строении организмов произошли в связи с освоением наземно-воздушной среды?
9. Чем можно объяснить более высокий уровень разнообразия организмов, обитающих в наземно-воздушной среде по сравнению с водной средой жизни?
10. Охарактеризуйте почвенную среду жизни как многофазную.
11. Расскажите о разнообразии жизни в почвенной среде.
12. Опишите основные особенности организма как среды жизни.
2. Свет как фактор-ресурс.
3. Вода как незаменимый фактор жизни организмов.
4. Макро– и микроэлементы в питании растений и животных.
5. Организмы как пищевые ресурсы.
6. Температура как экологический фактор.
7. Характеристика температурного режима конкретного региона (где живет студент).
8. Засоление воды и почвы как экологический фактор.
9. Экологическая роль горизонтального и вертикального перемещения водных масс.
10. Экологическое значение загрязняющих веществ в конкретном регионе (где живет студент).
11. Главные комплексные градиенты в конкретном регионе (где живет студент).
12. Сравнение водной и наземно-воздушной сред жизни.
13. Сравнение водной и почвенной сред жизни.
Глава 3. Основные принципы аутэкологии
3.1. Принцип экологического оптимума
3.2. Принцип индивидуальности экологии видов
3.3. Принцип лимитирующих факторов
Глава 4. Адаптации к абиотическим факторам
4.1. Определение понятия
4.2. Адаптивные комплексы
Важнейшей характеристикой воды является ее химический состав – содержание солей (в том числе биогенов), газов, ионов водорода (рН). По концентрации биогенов, особенно фосфора и азота, водоемы разделяются на олиготрофные, мезотрофные и эвтрофные. При повышении содержания биогенов, скажем, при загрязнении водоема стоками, происходит процесс эвтрофикации водных экосистем (см. 12.7).
Содержание кислорода в воде примерно в 20 раз ниже, чем в атмосфере, и составляет 6–8 мл/л. Оно снижается при повышении температуры, а также в стоячих водоемах в зимнее время, когда вода изолирована от атмосферы слоем льда. Снижение концентрации кислорода может стать причиной гибели многих обитателей водных экосистем, исключая особо устойчивые к дефициту кислорода виды, подобные карасю или линю, которые могут жить даже при снижении содержания кислорода до 0,5 мл/л.
Содержание углекислого газа в воде, напротив, выше, чем в атмосфере. В морской воде его может содержаться до 40–50 мл/л, что примерно в 150 раз выше, чем в атмосфере. Потребление углекислого газа фитопланктоном при интенсивном фотосинтезе не превышает 0,5 мл/л в сутки.
Концентрация ионов водорода в воде (рН) может меняться в пределах 3,7–7,8. Нейтральными считаются воды с рН от 6,45 до 7,3. Как уже отмечалось, с понижением рН биоразнообразие организмов, населяющих водную среду, быстро убывает. Речной рак, многие виды моллюсков гибнут при рН ниже 6, окунь и щука могут выдержать рН до 5, угорь и голец выживают при понижении рН до 5–4,4. В более кислых водах сохраняются лишь некоторые виды зоопланктона и фитопланктона. Кислотные дожди, связанные с выбросами в атмосферу больших количеств оксидов серы и азота промышленными предприятиями, стали причиной подкисления вод озер Европы и США и резкого обеднения их биологического разнообразия.
Наземно-воздушая среда жизни. Воздух отличается значительно более низкой плотностью по сравнению с водой. По этой причине освоение воздушной среды, которое произошло много позже, чем зарождение жизни и ее развитие в водной среде, сопровождалось усилением развития механических тканей, которые позволили организмам противостоять действию закона всемирного тяготения и ветра (скелет у позвоночных животных, хитиновые панцири у насекомых, склеренхима у растений). В условиях только воздушной среды ни один организм постоянно жить не может, и потому даже лучшие «летуны» (птицы и насекомые) должны периодически опускаться на землю. Перемещение организмов по воздуху возможно за счет специальных приспособлений – крыльев у птиц, насекомых, некоторых видов млекопитающих и даже рыб, парашютики и крылышки у семян, воздушные мешки у пыльцы хвойных пород и т.д.
Воздух – плохой проводник тепла, и потому именно в воздушной среде на суше возникли эндотермные (теплокровные) животные, которым легче сохранить тепло, чем эктотермным обитателям водной среды. Для теплокровных водных животных, включая гигантов-китов, водная среда вторична, предки этих животных когда-то жили на суше.
Для жизни в воздушной среде потребовались более сложные механизмы размножения, которые исключали бы риск высыхания половых клеток (многоклеточные антеридии и архегонии, а затем семязачатки и завязи у растений, внутреннее оплодотворение у животных, яйца с плотной оболочкой у птиц, пресмыкающихся, земноводных и др.).
В целом возможностей для формирования разнообразных сочетаний факторов в условиях наземно-воздушной среды много больше, чем водной. Именно в этой среде особенно ярко проявляются различия климата разных районов (и на разных высотах над уровнем моря в пределах одного района). Поэтому разнообразие наземных организмов много выше, чем водных.
Почвенная среда жизни. Большая часть суши покрыта тонким слоем (по сравнению с толщей земной коры) почвы, названной в.И. Вернадским биокосным телом. Почва представляет собой сложный многослойный «пирог» из горизонтов с разными свойствами, причем состав и толщина «пирога» в разных зонах различны. Общеизвестны зональный (от подзолов и серых лесных до черноземов, каштановых и бурых почв) и гидрогенный (от влажно-луговых до болотно-торфянистых) ряды почв. В южных районах почвы могут быть, кроме того, засолены на поверхности (солончаковатые почвы и солончаки) или в глубине (солонцы).
Любая почва представляет собой многофазную систему, в состав которой входят:
– минеральные частицы – от тончайшего ила до песка и гравия;
– органическое вещество – от тел только что умерших животных и отмерших корней растений до гумуса, в котором это органическое вещество подверглось сложной химической обработке;
– газовая (воздушная) фаза, характер которой во многом определяется физическими свойствами почвы – ее структурой и соответственно плотностью и порозностью. Газовая фаза почвы всегда обогащена углекислым газом и парами воды и может быть обеднена кислородом, что сближает условия жизни в почве с условиями водной среды;
– водная фаза. Вода в почве также может содержаться в разных количествах (от избытка до крайнего дефицита) и в разных качествах, быть гравитационной, т.е. свободно перемещающейся по капиллярам и наиболее доступной для корней растений и животных организмов, гигроскопической, т.е. входящей в состав коллоидных частиц, и газовой, т.е. в форме пара.
Эта многофазность почв делает их среду наиболее насыщенной жизнью. В почвах сконцентрирована основная биомасса животных, бактерий, грибов, в ней расположены корни растений, живущих в наземно-воздушной среде, но извлекающих из почвы воду с элементами питания и поставляющие в «темный мир» почвы органическое вещество, накопленное в процессе фотосинтеза на свету. Почва – это главный «цех по переработке» органического вещества, через нее протекает до 90% углерода, возвращаемого в атмосферу.
Гигантское разнообразие жизни в почве включает не только те организмы, которые живут в ней постоянно – позвоночные (кроты), членистоногие, бактерии, водоросли, дождевые черви и т.д., но и те организмы, которые связаны с ней лишь в начале своей «биографии» (саранчовые, многие жуки и т.д.).
Адаптация растений к некоторым вариантам экстремальных почвенных условий (засуха, засоление) будут рассмотрены в следующей главе.
Организмы как среда жизни. Нет ни одного вида многоклеточных организмов, который не был бы заселен другими организмами, и в первую очередь паразитами. Разные организмы и разные их органы, ткани и клетки имеют свою специфику как жизненные среды и свое паразитарное население, которое в разной степени опасно для хозяина, предоставившего им «жилплощадь со столом». Тем не менее, есть общие особенности этой среды жизни: в ней смягчены колебания внешних условий, и практически не ограничены ресурсы пищи.
Далеко не всегда хозяин, который заселен паразитами, оказывается беспомощным перед «назойливыми квартирантами». В этом случае паразитам приходится преодолевать неблагоприятные условия, формируемые в результате защитных реакций хозяина. По этой причине лучшая среда жизни для многих паразитов – это старые ослабленные особи.
Организмы как среду жизни, кроме паразитов, могут использовать многие виды, которые полезны организму-хозяину, т.е. находятся с ним в отношениях взаимовыгодного сотрудничества – мутуализма (см. 8.6).
Контрольные вопросы
1. Расскажите об основных параметрах водной среды жизни.2. Как организмы адаптируются к жизни в водной среде?
3. Какие факторы определяют способность света «пробивать» водную толщу?
4. Как изменяется спектр солнечного света при прохождении через водную толщу?
5. Какие факторы являются лимитирующими в водной среде?
6. Приведите примеры влияния рН водной среды на организмы.
7. Расскажите об основных отличиях наземно-воздушной и водной сред жизни.
8. Какие изменения в строении организмов произошли в связи с освоением наземно-воздушной среды?
9. Чем можно объяснить более высокий уровень разнообразия организмов, обитающих в наземно-воздушной среде по сравнению с водной средой жизни?
10. Охарактеризуйте почвенную среду жизни как многофазную.
11. Расскажите о разнообразии жизни в почвенной среде.
12. Опишите основные особенности организма как среды жизни.
Темы докладов на семинарских занятиях
1. Главные комплексные градиенты, определяющие характер экосистем конкретного региона (где живет студент).2. Свет как фактор-ресурс.
3. Вода как незаменимый фактор жизни организмов.
4. Макро– и микроэлементы в питании растений и животных.
5. Организмы как пищевые ресурсы.
6. Температура как экологический фактор.
7. Характеристика температурного режима конкретного региона (где живет студент).
8. Засоление воды и почвы как экологический фактор.
9. Экологическая роль горизонтального и вертикального перемещения водных масс.
10. Экологическое значение загрязняющих веществ в конкретном регионе (где живет студент).
11. Главные комплексные градиенты в конкретном регионе (где живет студент).
12. Сравнение водной и наземно-воздушной сред жизни.
13. Сравнение водной и почвенной сред жизни.
Глава 3. Основные принципы аутэкологии
В любой таксономической группе (среди насекомых, ракообразных, мхов, голосеменных и др.) виды разнообразны по требованиям к условиям среды и способности переносить неблагоприятные условия. Тем не менее, есть несколько экологических правил – принципов аутэкологии, которые проявляются в любой группе, хотя из этих правил всегда возможны исключения.
3.1. Принцип экологического оптимума
На градиенте любого экологического фактора распространение вида ограничено пределами толерантности (рис. 4). Между этими пределами есть отрезок, на котором условия для конкретного вида наиболее благоприятны и потому формируется самая большая биомасса и высокая плотность популяции. Это его экологический оптимум. Слева и справа от оптимума условия для жизни вида менее благоприятны. Это зоны пессимума, т.е. угнетения организмов, когда падает плотность популяции и вид становится наиболее уязвимыми к действию неблагоприятных экологических факторов (включая и влияние человека).
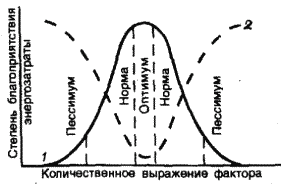 Рис. 4. Схема распределения вида на градиенте среды (по Шилову, 1998). 1 – степень благоприятствования условий среды для организма, 2 – величина энергозатрат на поддержание жизнедеятельности.
Рис. 4. Схема распределения вида на градиенте среды (по Шилову, 1998). 1 – степень благоприятствования условий среды для организма, 2 – величина энергозатрат на поддержание жизнедеятельности.
В зоне оптимума жизнь организма наиболее комфортна, и он затрачивает на ее поддержание минимальное количество энергии. В зонах пессимума для поддержания жизнедеятельности приходится тратить энергии много больше и включать специальные «механизмы выживания». Например для согревания в холоде теплокровные животные затрачивают энергию, накопленную в жировых тканях. Растения в условиях пессимума затрачивают большую часть продуктов фотосинтеза на дыхание и медленно растут.
Симметричная колоколовидная кривая, показанная на рисунке 4, является достаточно схематизированным приближением к реальной картине отношений видов к факторам среды. Часто распределения видов имеют асимметричный характер, т.е. кривая распределения более пологая с одной стороны и более крутая – с другой. Кроме того, иногда распределения могут быть двухвершинными или даже многовершинными, так как на некоторых отрезках градиента вид может быть подавлен более сильными конкурентами.
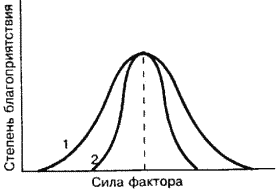 Рис. 5. Варианты отношения видов к факторам среды (по Шилову, 1998). 1 – эврибионтный вид, 2 – стенобионтный вид.
Рис. 5. Варианты отношения видов к факторам среды (по Шилову, 1998). 1 – эврибионтный вид, 2 – стенобионтный вид.
Разные виды имеют различные амплитуды распределения по градиентам факторов среды. Виды с узкой амплитудой называются стенобионтными, с широкой – эврибионтными (рис. 5). Так стенотермные и эвритермные организмы – это виды соответственно неутойчивые и устойчивые к колебаниям температуры. К эвритермным относятся многие насекомые (муравьи, жуки-ксилофаги и др.). Пример стенотермности в животном мире – веслоногий рачок (Copilla mirabile), который не выдерживает изменений температуры за пределами 23–29°C (Шилов, 1998). Стенотермными являются деревья тропического леса, которые выдерживают колебания температуры в интервале 5–8°C. Классический пример эвритермности – Лиственница Гмелина (Larix gmelinii), которая в Якутии выдерживает колебания температуры от плюс 30 до минус 70 градусов. Большинство видов деревьев средней полосы Европы выдерживает колебания температуры в пределах 60 градусов. Эта эвритермность во многом связана со способностью деревьев переживать холодное время года в состоянии покоя.
Большой опыт изучения отношения видов к разным факторам среды накоплен ботаниками. Они определяли диапазоны распределения видов по отношению к увлажнению, богатству почвы элементами минерального питания, интенсивности пастбищной нагрузки и т.д. с целью последующей оценки характера экотопов по составу видов растительных сообществ.
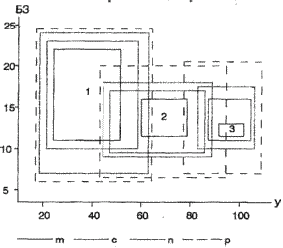 Рис. 6. Экологические ареалы трех видов растений по отношению к факторам увлажнения (У) и богатства почвы элементами минерального питания (БЗ, по Раменскому и др., 1956). Градации проективного покрытия: m – более 8%, c – 8–2,5%, n – 2,4–3 %, p – 0,2—0,1 %.
Рис. 6. Экологические ареалы трех видов растений по отношению к факторам увлажнения (У) и богатства почвы элементами минерального питания (БЗ, по Раменскому и др., 1956). Градации проективного покрытия: m – более 8%, c – 8–2,5%, n – 2,4–3 %, p – 0,2—0,1 %.
На рис. 6 показаны экологические ареалы трех видов растений разной экологии, связанных с сухими местообитаниями – типчака (Festuca valesiaca, 1), условиями умеренного увлажнения – овсяницы луговой (Festuca pratensis, 2) и переувлажненными почвами – осоки лисьей (Carex vulpina, 3). Рисунок иллюстрирует общеизвестные экологические закономерности:
– при уменьшении обилия расширяется экологическая амплитуда вида;
– кривые распределения видов по градиентам факторов среды могут налагаться друг на друга при невысоком обилии, но оказываются разомкнутыми при высоком обилии.
2. Какие формы может иметь кривая распределения вида на градиенте экологического фактора, кроме колоколовидной?
3. Приведите примеры стенобионтных и эврибионтных видов.
В зоне оптимума жизнь организма наиболее комфортна, и он затрачивает на ее поддержание минимальное количество энергии. В зонах пессимума для поддержания жизнедеятельности приходится тратить энергии много больше и включать специальные «механизмы выживания». Например для согревания в холоде теплокровные животные затрачивают энергию, накопленную в жировых тканях. Растения в условиях пессимума затрачивают большую часть продуктов фотосинтеза на дыхание и медленно растут.
Симметричная колоколовидная кривая, показанная на рисунке 4, является достаточно схематизированным приближением к реальной картине отношений видов к факторам среды. Часто распределения видов имеют асимметричный характер, т.е. кривая распределения более пологая с одной стороны и более крутая – с другой. Кроме того, иногда распределения могут быть двухвершинными или даже многовершинными, так как на некоторых отрезках градиента вид может быть подавлен более сильными конкурентами.
Разные виды имеют различные амплитуды распределения по градиентам факторов среды. Виды с узкой амплитудой называются стенобионтными, с широкой – эврибионтными (рис. 5). Так стенотермные и эвритермные организмы – это виды соответственно неутойчивые и устойчивые к колебаниям температуры. К эвритермным относятся многие насекомые (муравьи, жуки-ксилофаги и др.). Пример стенотермности в животном мире – веслоногий рачок (Copilla mirabile), который не выдерживает изменений температуры за пределами 23–29°C (Шилов, 1998). Стенотермными являются деревья тропического леса, которые выдерживают колебания температуры в интервале 5–8°C. Классический пример эвритермности – Лиственница Гмелина (Larix gmelinii), которая в Якутии выдерживает колебания температуры от плюс 30 до минус 70 градусов. Большинство видов деревьев средней полосы Европы выдерживает колебания температуры в пределах 60 градусов. Эта эвритермность во многом связана со способностью деревьев переживать холодное время года в состоянии покоя.
Большой опыт изучения отношения видов к разным факторам среды накоплен ботаниками. Они определяли диапазоны распределения видов по отношению к увлажнению, богатству почвы элементами минерального питания, интенсивности пастбищной нагрузки и т.д. с целью последующей оценки характера экотопов по составу видов растительных сообществ.
На рис. 6 показаны экологические ареалы трех видов растений разной экологии, связанных с сухими местообитаниями – типчака (Festuca valesiaca, 1), условиями умеренного увлажнения – овсяницы луговой (Festuca pratensis, 2) и переувлажненными почвами – осоки лисьей (Carex vulpina, 3). Рисунок иллюстрирует общеизвестные экологические закономерности:
– при уменьшении обилия расширяется экологическая амплитуда вида;
– кривые распределения видов по градиентам факторов среды могут налагаться друг на друга при невысоком обилии, но оказываются разомкнутыми при высоком обилии.
Контрольные вопросы
1. На какие градации делится градиент фактора среды между пределами толерантности вида?2. Какие формы может иметь кривая распределения вида на градиенте экологического фактора, кроме колоколовидной?
3. Приведите примеры стенобионтных и эврибионтных видов.
3.2. Принцип индивидуальности экологии видов
Каждый вид индивидуально распределяется по экологическим факторам, кривые распределений разных видов перекрываются, но их оптимумы различаются.
По этой причине при изменении условий среды в пространстве (например,от сухой вершины холма к влажному логу) или во времени (при пересыхании озера, при усилении выпаса, при зарастании скал) состав экосистем изменяется постепенно. Известный российский эколог Л.Г. Раменский сформулировал этот закон образно: «Виды – это не рота солдат, марширующих в ногу».
Индивидуальность экологии видов – это то свойство, которое «крайне неудобно» для практического использования данных о связи видов и условий среды. По этой причине для удобства использования в практике (т.е. из прагматических соображений) знание об индивидуальных особенностях видов упрощается (редуцируется) до системы экологических групп. Такие группы объединяют не тождественные по экологии виды, а виды с близким отношением к одному или нескольким факторам среды.
Для выделения экологических групп градиент среды разбивается на некоторое число классов, и к одной экологической группе относятся виды, которые имеют зоны оптимума в этом классе. Количество классов зависит от задачи, которая была поставлена при выделении экологических групп, и полноты знаний о распределении видов по градиенту экологического фактора, и потому экологические группы видов могут быть выделены в более крупном и более мелком масштабе.
Так например,по отношению к фактору увлажнения почвы все виды растений можно разбить на три группы:
1. ксерофиты – виды сухих местообитаний;
2. мезофиты – виды нормально увлажненных местообитаний;
3. гидрофиты – виды переувлажненных местообитаний.
Однако этот ряд может быть разбит на 5 градаций за счет двух дополнительных переходных групп и тогда примет следующий вид:
1. ксерофиты,
2. ксеромезофиты,
3. мезофиты,
4. мезогидрофиты,
5. гидрофиты.
При наличии более обширного материала о распределении видов вдоль градиентов среды этот ряд может быть увеличен до 7 групп за счет дополнительных переходных групп между мезофитами и гидрофитами. Кроме того, наряду с гидрофитами (типичными водными растениями), можно выделить группу гигрофитов – растений переувлажненных почв, которые покрыты водой лишь часть вегетационного сезона или не заливаются вообще, а увлажнены только за счет приближенных к поверхности грунтовых вод.
Задача выделения экологических групп имеет множество аналогий в практической жизни. К примеру, форма и размер стопы человека индивидуальны, тем не менее, из соображений удобства обувь шьется по системе размеров. Поскольку разделение любой совокупности индивидуальных объектов на группы противоречит их природе, периодически возникают ситуации, когда трудно определить принадлежность объекта к группе: один размер обуви еще мал, а следующий – уже велик. Условность отнесения видов к экологическим группам еще больше. Решить вопрос о том, к какой группе относится вид – ксеромезофитам или мезоксерофитам, гигромезофитам или мезогигрофитам – часто бывает очень сложно.
Важным следствием принципа индивидуальности экологии видов является постепенность изменения состава растительных сообществ и экосистем вдоль градиентов среды. Такие постепенные изменения называются континуумом (непрерывностью). По этой причине конкретные сообщества и экосистемы выделяются также условно, как и экологические группы видов.
Концепция континуума была сформулирована в начале ХХ в. независимо двумя учеными – россиянином Л.Г. Раменским и американцем Г. Глизоном (H. Gleason). Во второй половине ХХ в. наибольший вклад в ее развитие внесли Р. Уиттекер (R. Whittaker), Дж. Кертис (J. Curtis), Р. Макинтош (R. McIntosh) и М. Остин (M. Austin).
Различают континуумы двух типов: экоклин и экотон. Экоклин – это абсолютный континуум, внутри которого на градиенте не выделяется зон быстрого и медленного изменений видового состава сообществ. Этот тип континуума преобладает в тех случаях, когда изменение состава сообществ происходит без смены жизненной формы растений, т.е. меняется травяная или лесная растительность. Пример экоклина приведен на рис. 7, из него очевидно, что смена растительных сообществ на градиенте происходит постепенно и провести границы сообществ, соответствующих разным условиям засоления почвы, можно только условно.
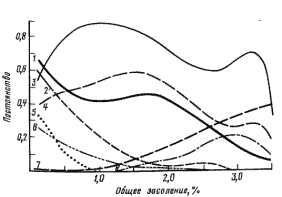 Рис. 7. Экоклин травяной растительности поймы нижней Волги по градиенту общего засоления (список видов сокращен, по Golub, Mirkin, 1986). 1 – Phalaroides arundinacea, 2 – Bolboschoenus borodinii, 6 – Argusia sibirica, 7 – Atriplex litoralis, 8 – Tripolium vulgare.
Рис. 7. Экоклин травяной растительности поймы нижней Волги по градиенту общего засоления (список видов сокращен, по Golub, Mirkin, 1986). 1 – Phalaroides arundinacea, 2 – Bolboschoenus borodinii, 6 – Argusia sibirica, 7 – Atriplex litoralis, 8 – Tripolium vulgare.
Экотон – это тип континуума, при котором на градиенте формируются более или менее однородные сообщества, связанные зоной быстрого и видимого на глаз перехода. Типичный пример экотона – растительность опушки (рис. 8), т.е. зоны контакта леса и злаковника (луга или степи).
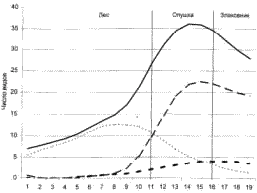 Рис. 8. Экотопный эффект в растительности лесной опушки (по Кучеровой, 2001). Число видов: 1 – общее, 2 – злаковников, 3 – лесных, 4 – опушечных.
Рис. 8. Экотопный эффект в растительности лесной опушки (по Кучеровой, 2001). Число видов: 1 – общее, 2 – злаковников, 3 – лесных, 4 – опушечных.
Аналогичные континуумы типов экоклин и экотон проявляются и в характере изменения гетеротрофной биоты, в первую очередь насекомых и почвенных животных.
2. Что такое экологические группы видов, с чем связана их условность?
3. Расскажите об истории развития и содержании концепции континуума.
4. Чем отличаются экоклин и экотон?
По этой причине при изменении условий среды в пространстве (например,от сухой вершины холма к влажному логу) или во времени (при пересыхании озера, при усилении выпаса, при зарастании скал) состав экосистем изменяется постепенно. Известный российский эколог Л.Г. Раменский сформулировал этот закон образно: «Виды – это не рота солдат, марширующих в ногу».
Индивидуальность экологии видов – это то свойство, которое «крайне неудобно» для практического использования данных о связи видов и условий среды. По этой причине для удобства использования в практике (т.е. из прагматических соображений) знание об индивидуальных особенностях видов упрощается (редуцируется) до системы экологических групп. Такие группы объединяют не тождественные по экологии виды, а виды с близким отношением к одному или нескольким факторам среды.
Для выделения экологических групп градиент среды разбивается на некоторое число классов, и к одной экологической группе относятся виды, которые имеют зоны оптимума в этом классе. Количество классов зависит от задачи, которая была поставлена при выделении экологических групп, и полноты знаний о распределении видов по градиенту экологического фактора, и потому экологические группы видов могут быть выделены в более крупном и более мелком масштабе.
Так например,по отношению к фактору увлажнения почвы все виды растений можно разбить на три группы:
1. ксерофиты – виды сухих местообитаний;
2. мезофиты – виды нормально увлажненных местообитаний;
3. гидрофиты – виды переувлажненных местообитаний.
Однако этот ряд может быть разбит на 5 градаций за счет двух дополнительных переходных групп и тогда примет следующий вид:
1. ксерофиты,
2. ксеромезофиты,
3. мезофиты,
4. мезогидрофиты,
5. гидрофиты.
При наличии более обширного материала о распределении видов вдоль градиентов среды этот ряд может быть увеличен до 7 групп за счет дополнительных переходных групп между мезофитами и гидрофитами. Кроме того, наряду с гидрофитами (типичными водными растениями), можно выделить группу гигрофитов – растений переувлажненных почв, которые покрыты водой лишь часть вегетационного сезона или не заливаются вообще, а увлажнены только за счет приближенных к поверхности грунтовых вод.
Задача выделения экологических групп имеет множество аналогий в практической жизни. К примеру, форма и размер стопы человека индивидуальны, тем не менее, из соображений удобства обувь шьется по системе размеров. Поскольку разделение любой совокупности индивидуальных объектов на группы противоречит их природе, периодически возникают ситуации, когда трудно определить принадлежность объекта к группе: один размер обуви еще мал, а следующий – уже велик. Условность отнесения видов к экологическим группам еще больше. Решить вопрос о том, к какой группе относится вид – ксеромезофитам или мезоксерофитам, гигромезофитам или мезогигрофитам – часто бывает очень сложно.
Важным следствием принципа индивидуальности экологии видов является постепенность изменения состава растительных сообществ и экосистем вдоль градиентов среды. Такие постепенные изменения называются континуумом (непрерывностью). По этой причине конкретные сообщества и экосистемы выделяются также условно, как и экологические группы видов.
Концепция континуума была сформулирована в начале ХХ в. независимо двумя учеными – россиянином Л.Г. Раменским и американцем Г. Глизоном (H. Gleason). Во второй половине ХХ в. наибольший вклад в ее развитие внесли Р. Уиттекер (R. Whittaker), Дж. Кертис (J. Curtis), Р. Макинтош (R. McIntosh) и М. Остин (M. Austin).
Различают континуумы двух типов: экоклин и экотон. Экоклин – это абсолютный континуум, внутри которого на градиенте не выделяется зон быстрого и медленного изменений видового состава сообществ. Этот тип континуума преобладает в тех случаях, когда изменение состава сообществ происходит без смены жизненной формы растений, т.е. меняется травяная или лесная растительность. Пример экоклина приведен на рис. 7, из него очевидно, что смена растительных сообществ на градиенте происходит постепенно и провести границы сообществ, соответствующих разным условиям засоления почвы, можно только условно.
Экотон – это тип континуума, при котором на градиенте формируются более или менее однородные сообщества, связанные зоной быстрого и видимого на глаз перехода. Типичный пример экотона – растительность опушки (рис. 8), т.е. зоны контакта леса и злаковника (луга или степи).
Аналогичные континуумы типов экоклин и экотон проявляются и в характере изменения гетеротрофной биоты, в первую очередь насекомых и почвенных животных.
Контрольные вопросы
1. Объясните содержание принципа индивидуальности экологии вида.2. Что такое экологические группы видов, с чем связана их условность?
3. Расскажите об истории развития и содержании концепции континуума.
4. Чем отличаются экоклин и экотон?
3.3. Принцип лимитирующих факторов
Суть этого принципа, связанного с именем Ю. Либиха, заключается в том, что наиболее важным для распределения вида является тот фактор, значения которого находятся в минимуме или в максимуме.
Как уже отмечалось, наличие в составе комплексного градиента лимитирующего фактора делает этот градиент ведущим, т.е. в большей степени, чем другие, определяющим состав и продуктивность экосистем и состояние популяций формирующих их видов. Так в зоне тундры основу ведущего комплексного градиента составляет количество тепла, так как влаги там достаточно, а обеспеченность элементами минерального питания также зависит от тепла: чем теплее субстраты, тем активнее идет в них процесс минерализации органического вещества и меньше накапливается неразложившихся остатков растений.
В зоне тайги лимитирующим фактором является обеспеченность почв питательными элементами. Почвы, формирующиеся на карбонатных породах, которые богаты кальцием и другими минеральными элементами, позволяют формироваться очень продуктивным сообществам. Однако в условиях тайги, особенно в южной ее части, возрастает роль второго ведущего комплексного градиента, связанного с увлажнением.
В лесостепной и степной зонах ведущий комплексный градиент формируют экологические факторы, связанные с лимитирующим фактором увлажнения, так как почвы в этих зонах (черноземы) богаты питательными элементами. Особенно остро влияние лимитирующего фактора проявляется в годы засух. На этот комплексный градиент может накладываться влияние выпаса (как отмечалось, выпас – это также комплексный градиент), а в южной части степной зоны – комплексного градиента засоления почвы.
В водных экосистемах для большинства входящих в их состав организмов лимитирующими факторами являются содержание кислорода и фосфора, а для растений, кроме того, – свет.
Для многих животных в условиях умеренного климата лимитирующим фактором является глубина снежного покрова. Свободное перемещение по глубокому снегу свойственно сравнительно небольшому числу видов, имеющих «лыжи» (заяц-беляк, заяц-русак, белая куропатка) или отличающихся «длинноногостью» (лось). Глубокий снег является препятствием для перемещения волка и кабана. Этот фактор определяет границы коневодства с круглогодичным содержанием животных на пастбищах. При глубоком снеге (в период зимнего выпаса – тебеневки) лошади не могут использовать корм.
2. Приведите примеры лимитирующих факторов в наземных экосистемах разных природных зон.
3. Какие факторы являются лимитирующими в водных экосистемах?
2. Концепция континуума: история и значение для экологии.
3. Роль лимитирующих факторов в определении состава биоты экосистем.
Как уже отмечалось, наличие в составе комплексного градиента лимитирующего фактора делает этот градиент ведущим, т.е. в большей степени, чем другие, определяющим состав и продуктивность экосистем и состояние популяций формирующих их видов. Так в зоне тундры основу ведущего комплексного градиента составляет количество тепла, так как влаги там достаточно, а обеспеченность элементами минерального питания также зависит от тепла: чем теплее субстраты, тем активнее идет в них процесс минерализации органического вещества и меньше накапливается неразложившихся остатков растений.
В зоне тайги лимитирующим фактором является обеспеченность почв питательными элементами. Почвы, формирующиеся на карбонатных породах, которые богаты кальцием и другими минеральными элементами, позволяют формироваться очень продуктивным сообществам. Однако в условиях тайги, особенно в южной ее части, возрастает роль второго ведущего комплексного градиента, связанного с увлажнением.
В лесостепной и степной зонах ведущий комплексный градиент формируют экологические факторы, связанные с лимитирующим фактором увлажнения, так как почвы в этих зонах (черноземы) богаты питательными элементами. Особенно остро влияние лимитирующего фактора проявляется в годы засух. На этот комплексный градиент может накладываться влияние выпаса (как отмечалось, выпас – это также комплексный градиент), а в южной части степной зоны – комплексного градиента засоления почвы.
В водных экосистемах для большинства входящих в их состав организмов лимитирующими факторами являются содержание кислорода и фосфора, а для растений, кроме того, – свет.
Для многих животных в условиях умеренного климата лимитирующим фактором является глубина снежного покрова. Свободное перемещение по глубокому снегу свойственно сравнительно небольшому числу видов, имеющих «лыжи» (заяц-беляк, заяц-русак, белая куропатка) или отличающихся «длинноногостью» (лось). Глубокий снег является препятствием для перемещения волка и кабана. Этот фактор определяет границы коневодства с круглогодичным содержанием животных на пастбищах. При глубоком снеге (в период зимнего выпаса – тебеневки) лошади не могут использовать корм.
Контрольные вопросы
1. Кто является автором принципа лимитирующих факторов, какие факторы называются лимитирующими?2. Приведите примеры лимитирующих факторов в наземных экосистемах разных природных зон.
3. Какие факторы являются лимитирующими в водных экосистемах?
Темы докладов на семинарском занятии
1. Для чего и как выделяют экологические группы видов?2. Концепция континуума: история и значение для экологии.
3. Роль лимитирующих факторов в определении состава биоты экосистем.
Глава 4. Адаптации к абиотическим факторам
Единство организмов с условиями среды их обитания, которое достигается за счет адаптаций (приспособлений), является прописной истиной и составляет основу учения Ч. Дарвина. В этой главе будут рассмотрены общие вопросы природы адаптаций и несколько конкретных примеров адаптации разных организмов к разным условиям среды.
4.1. Определение понятия
Адаптация – это приспособление организма к определенным условиям среды, которое достигается за счет комплекса признаков – морфологических, физиологических, поведенческих. В результате адаптаций возникают организмы, приспособленные к различным условиям среды. Адаптациями объясняется различный состав экосистем разных экологических условий.
И.А. Шилов (1998) подчеркивает, что в эволюции крупных таксонов адаптация к лимитирующим факторам определяла наиболее крупные изменения морфологии и физиологии. Так выход позвоночных животных на сушу был невозможен без преодоления двух лимитирующих факторов – малой плотности среды и низкой влажности. В результате произошла замена «парящей» локомоции на развитие конечностей рычажного типа, способных обеспечить поступательное движение (возникновение «четвероногих») и активный полет птиц.
При выходе растений на сушу эти же лимитирующие факторы привели к возникновению высших растений. В условиях воздушной среды (исключая ту часть растений, которая сохранила связь с водной средой обитания) растения должны были решить комплекс адаптационных задач:
– не засохнуть (развить проводящую систему, корневую систему, покровные ткани с устьицами);
– не упасть (развить механические ткани, способные противостоять гравитационной силе и ветру);
– не умереть с голоду (сформировать мощную листовую поверхность для фотосинтеза и увеличивать всасывание корнями элементов минерального питания за счет симбиоза с грибами и бактериями);
– обеспечить условия для размножения – сформировать защищенные от высыхания органы бесполого и полового размножения – многоклеточные спорангии и гаметангии.
Как высшая форма адаптации к жизни в условиях наземной среды возникли семенные растения с «маленьким шедевром эволюции» – семенем (Тахтаджян, 1978) и объединением спорофита и гаметофита в единый организм.
Адаптации животных к переживанию неблагоприятных условий в целом более разнообразны, чем у растений. Их можно свести к трем основным направлениям:
– уход от неблагоприятных условий (миграция птиц, кочевка оленей и других копытных в поисках корма, зарывание в песок, почву или снег и др.);
– переход в состояние анабиоза – резкого снижения активности процессов жизнедеятельности (покоящиеся стадии у беспозвоночных животных, прекращение активности рептилий при низких температурах, зимняя спячка млекопитающих и др.);
– развитие приспособлений для жизни в неблагоприятных условиях (шерстный покров и подкожный жир у животных в условиях холодного климата, экономное использование воды у пустынных животных и т.д.).
Растения «заякорены», т.е. ведут прикрепленный образ жизни, и потому у них возможны лишь два варианта адаптаций: снижение интенсивности процессов жизнедеятельности в неблагоприятные периоды (сбрасывание листьев, перезимовывание в стадии погребенных в почву органов – луковиц, корневищ, клубней, а также семян и спор, полный криптобиоз у моховидных) или повышение устойчивости к неблагоприятным факторам за счет специальных физиологических механизмов.
Разнообразие физиологических механизмов переживания неблагоприятных абиотических (засуха, засоление, дефицит света, холод) и биотических (влияние фитофагов, конкуренция с другими растениями) факторов компенсирует отсутствие у растений способности к миграциям в более благоприятные экологические условия.
Впрочем, наблюдаемые изменения в поведении организмов часто связаны не с адаптациями, а с преадаптациями – «скрытыми» приспособлениями, которые проявились и оказались полезными при появлении нового фактора. Преадаптациями объясняется устойчивость некоторых экотипов сорных растений к действию гербицидов и некоторых видов деревьев к загрязнению атмосферы промышленностью и транспортом.
Экологи используют также понятие «экзаптация» (Gould, Vrba, 1982; Гиляров, 2003), которое обозначает приспособление, первоначально возникшее для одной цели, но затем оказавшееся полезным для решения другой задачи (например, перья у птиц возникли как адаптация для защиты от холода, но в дальнейшем оказались полезными для полета).
Экзаптациями объясняются некоторые «странные» явления природы, переток элементов питания из растения одного вида в другой по микоризе. Микоризным грибам «выгодно» сотрудничать с несколькими видами деревьев, и переток элементов питания является побочным следствием этой адаптации.
2. Расскажите об адаптации растений к наземному образу жизни.
3. Расскажите об адаптации животных к наземно-воздушной среде жизни.
4. Чем различаются адаптации к неблагоприятным условиям среды у растений и животных?
5. Что такое преадаптация и экзаптация?
И.А. Шилов (1998) подчеркивает, что в эволюции крупных таксонов адаптация к лимитирующим факторам определяла наиболее крупные изменения морфологии и физиологии. Так выход позвоночных животных на сушу был невозможен без преодоления двух лимитирующих факторов – малой плотности среды и низкой влажности. В результате произошла замена «парящей» локомоции на развитие конечностей рычажного типа, способных обеспечить поступательное движение (возникновение «четвероногих») и активный полет птиц.
При выходе растений на сушу эти же лимитирующие факторы привели к возникновению высших растений. В условиях воздушной среды (исключая ту часть растений, которая сохранила связь с водной средой обитания) растения должны были решить комплекс адаптационных задач:
– не засохнуть (развить проводящую систему, корневую систему, покровные ткани с устьицами);
– не упасть (развить механические ткани, способные противостоять гравитационной силе и ветру);
– не умереть с голоду (сформировать мощную листовую поверхность для фотосинтеза и увеличивать всасывание корнями элементов минерального питания за счет симбиоза с грибами и бактериями);
– обеспечить условия для размножения – сформировать защищенные от высыхания органы бесполого и полового размножения – многоклеточные спорангии и гаметангии.
Как высшая форма адаптации к жизни в условиях наземной среды возникли семенные растения с «маленьким шедевром эволюции» – семенем (Тахтаджян, 1978) и объединением спорофита и гаметофита в единый организм.
Адаптации животных к переживанию неблагоприятных условий в целом более разнообразны, чем у растений. Их можно свести к трем основным направлениям:
– уход от неблагоприятных условий (миграция птиц, кочевка оленей и других копытных в поисках корма, зарывание в песок, почву или снег и др.);
– переход в состояние анабиоза – резкого снижения активности процессов жизнедеятельности (покоящиеся стадии у беспозвоночных животных, прекращение активности рептилий при низких температурах, зимняя спячка млекопитающих и др.);
– развитие приспособлений для жизни в неблагоприятных условиях (шерстный покров и подкожный жир у животных в условиях холодного климата, экономное использование воды у пустынных животных и т.д.).
Растения «заякорены», т.е. ведут прикрепленный образ жизни, и потому у них возможны лишь два варианта адаптаций: снижение интенсивности процессов жизнедеятельности в неблагоприятные периоды (сбрасывание листьев, перезимовывание в стадии погребенных в почву органов – луковиц, корневищ, клубней, а также семян и спор, полный криптобиоз у моховидных) или повышение устойчивости к неблагоприятным факторам за счет специальных физиологических механизмов.
Разнообразие физиологических механизмов переживания неблагоприятных абиотических (засуха, засоление, дефицит света, холод) и биотических (влияние фитофагов, конкуренция с другими растениями) факторов компенсирует отсутствие у растений способности к миграциям в более благоприятные экологические условия.
Впрочем, наблюдаемые изменения в поведении организмов часто связаны не с адаптациями, а с преадаптациями – «скрытыми» приспособлениями, которые проявились и оказались полезными при появлении нового фактора. Преадаптациями объясняется устойчивость некоторых экотипов сорных растений к действию гербицидов и некоторых видов деревьев к загрязнению атмосферы промышленностью и транспортом.
Экологи используют также понятие «экзаптация» (Gould, Vrba, 1982; Гиляров, 2003), которое обозначает приспособление, первоначально возникшее для одной цели, но затем оказавшееся полезным для решения другой задачи (например, перья у птиц возникли как адаптация для защиты от холода, но в дальнейшем оказались полезными для полета).
Экзаптациями объясняются некоторые «странные» явления природы, переток элементов питания из растения одного вида в другой по микоризе. Микоризным грибам «выгодно» сотрудничать с несколькими видами деревьев, и переток элементов питания является побочным следствием этой адаптации.
Контрольные вопросы
1. Что такое адаптация?2. Расскажите об адаптации растений к наземному образу жизни.
3. Расскажите об адаптации животных к наземно-воздушной среде жизни.
4. Чем различаются адаптации к неблагоприятным условиям среды у растений и животных?
5. Что такое преадаптация и экзаптация?
4.2. Адаптивные комплексы
Как уже отмечалось, адаптация любого организма к условиям среды достигается за счет комплекса признаков, при этом набор адаптивных признаков бывает достаточно разнообразным. Поэтому разные организмы используют лишь часть потенциально возможных адаптивных признаков. Так переживание животными экстремальных условий в пустыне (высокие температуры, дефицит воды) возможно как за счет физиологических (запас жира с получением воды путем его расщепления), так и поведенческих адаптаций (зарывание в песок). Для переживания холода животные могут сжигать некоторое количество жира, сохранять тепло за счет густого шерстного или перьевого покрова, снижать активность жизнедеятельности (впадать в спячку), зарываться в снег (временно, или зимовать под снегом).
Конец бесплатного ознакомительного фрагмента