Страница:
При синтезе ведущей нити основную роль играют ДНК-полимераза III Е. coli, ошибочность которой составляет 10-6, а у эукариот ДНК-полимеразы ε и δ (ошибочность 10-5). Различные автономные экзонуклеазы способны повышать точность этих процессивных полимераз в 5-10 раз.
При синтезе отстающей нити у Е. соli в объединении фрагментов Оказаки участвует ДНК-полимераза I. Точность этой умеренно процессивной полимеразы резко увеличивается в присутствии автономных экзонуклеаз.
Большинство систем репарации ДНК нуждаются в синтезе ДНК при исправлении ошибок. У Е. соli в репарации ДНК существенное участие принимает ДНК-полимераза I, точность работы которой явно увеличивается в присутствии автономных экзонуклеаз. В клетке человека за сутки образуется в среднем 105 спонтанных повреждений ДНК в результате ее дезаминирования, депуринизации, окисления и неправильного метилирования. Большинство этих повреждений репарируются при участии ДНК-полимеразы β (ошибочность 10-3), В результате этого в транскрибируемой ДНК каждый раунд репликации появляются приблизительно 3 ошибки, подлежащие коррекции. При работе другой системы репарации непроцессивные безнуклеазные сверхошибочные (ошибочность 10-2) ДНК-полимеразы ведут синтез ДНК в обход повреждений в ДНК-матрице. После завершения синтеза на поврежденной матрице они должны быть заменены, по-видимому, с помощью репликативного фактора С, на основную элонгирующую ДНК-полимеразу δ (ошибочность 10-5). Но в силу той же инерционности биохимических реакций замена не происходит мгновенно, что требует исправления ошибок (совершенных непроцессивными полимеразами напротив неповрежденной матрицы), в том числе с участием автономных экзонуклеаз. В экстрактах клеток человека недавно показана экзонуклеазная коррекция ошибок, допущенных сверхошибочной ДНК-полимеразой η (эта). Наконец, автономные экзонуклеазы участвуют в коррекции гетеродуплексов.
Все виды экзонуклеазной коррекции должны закончиться за время данной репликации. По-видимому, коррекция ДНК-полимеразных ошибок – весьма эффективный процесс, поскольку анализ генома человека показал, что дивергенция последовательностей в транскрибируемой ДНК составляет примерно 0,1 % при исследовании ДНК от 24 человек различных этнических групп.
2. Типы повреждений ДНК
3. Многообразие систем репарации ДНК
4. Прямая репарация ДНК
4.1. Фотореактивация
4.2. Репарация О6-алкилированного гуанина
4.3. Репарация однонитевых разрывов ДНК
4.4. Репарация АР-сайтов за счет прямой вставки пуринов
5. Эксцизионная репарация
5.1. Эксцизионная репарация оснований (base excision repair, BER)
При синтезе отстающей нити у Е. соli в объединении фрагментов Оказаки участвует ДНК-полимераза I. Точность этой умеренно процессивной полимеразы резко увеличивается в присутствии автономных экзонуклеаз.
Большинство систем репарации ДНК нуждаются в синтезе ДНК при исправлении ошибок. У Е. соli в репарации ДНК существенное участие принимает ДНК-полимераза I, точность работы которой явно увеличивается в присутствии автономных экзонуклеаз. В клетке человека за сутки образуется в среднем 105 спонтанных повреждений ДНК в результате ее дезаминирования, депуринизации, окисления и неправильного метилирования. Большинство этих повреждений репарируются при участии ДНК-полимеразы β (ошибочность 10-3), В результате этого в транскрибируемой ДНК каждый раунд репликации появляются приблизительно 3 ошибки, подлежащие коррекции. При работе другой системы репарации непроцессивные безнуклеазные сверхошибочные (ошибочность 10-2) ДНК-полимеразы ведут синтез ДНК в обход повреждений в ДНК-матрице. После завершения синтеза на поврежденной матрице они должны быть заменены, по-видимому, с помощью репликативного фактора С, на основную элонгирующую ДНК-полимеразу δ (ошибочность 10-5). Но в силу той же инерционности биохимических реакций замена не происходит мгновенно, что требует исправления ошибок (совершенных непроцессивными полимеразами напротив неповрежденной матрицы), в том числе с участием автономных экзонуклеаз. В экстрактах клеток человека недавно показана экзонуклеазная коррекция ошибок, допущенных сверхошибочной ДНК-полимеразой η (эта). Наконец, автономные экзонуклеазы участвуют в коррекции гетеродуплексов.
Все виды экзонуклеазной коррекции должны закончиться за время данной репликации. По-видимому, коррекция ДНК-полимеразных ошибок – весьма эффективный процесс, поскольку анализ генома человека показал, что дивергенция последовательностей в транскрибируемой ДНК составляет примерно 0,1 % при исследовании ДНК от 24 человек различных этнических групп.
2. Типы повреждений ДНК
Большинство повреждений ДНК не являются результатом только ошибок репликации. Множество повреждений возникает в любое время клеточного цикла под действием как экзогенных, так и эндогенных факторов. В табл. 1. схематично изображены основные типы повреждений ДНК, которые опознаются и устраняются различными системами репарации.
Ультрафиолетовые лучи вызывают образование пиримидиновых димеров, 6,4-фотопродуктов, аддуктов, разрывов и прочие повреждения ДНК. Под действием химических агентов происходят разного рода модификвции нуклеотидов, возникают межнитевые сшивки, конформационные дефекты. Двунитевые разрывы могут приводить к перестройкам хромосом, что и является главной причиной летального действия ионизирующей радиации.
Таблица 1. Основные типы повреждений ДНК.
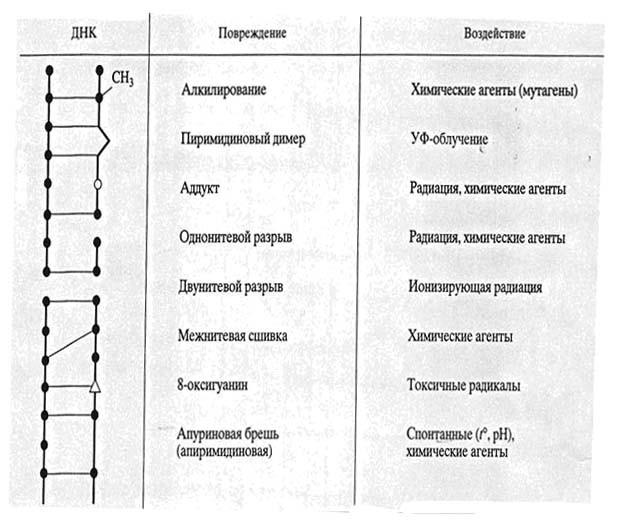
В результате внутриклеточных процессов в ДНК образуются многочисленные АP-сайты из-за спонтанной утраты пуринов/пиримидинов, окисленные участки (например, 8-оксигуанин), возникающие под действием токсических радикалов, постоянно генерируемых в процессах метаболизма.
Следует заметить, что все повреждения показаны только схематически, так как любое из них вызывает локальное изменение структуры ДНК вокруг, а разрывы ДНК почти всегда сопровождаются и модификацией прилежащих к ним оснований. Кроме обычно упоминаемых пиримидиновых димеров, под действием УФ облучения образуются и другие повреждения, например, 6,4-фотопродукты. В дальнейшем, описывая различные системы репарации ДНК, мы будем подробно останавливаться на тех типах повреждений, которые они способны исправлять.
Ультрафиолетовые лучи вызывают образование пиримидиновых димеров, 6,4-фотопродуктов, аддуктов, разрывов и прочие повреждения ДНК. Под действием химических агентов происходят разного рода модификвции нуклеотидов, возникают межнитевые сшивки, конформационные дефекты. Двунитевые разрывы могут приводить к перестройкам хромосом, что и является главной причиной летального действия ионизирующей радиации.
Таблица 1. Основные типы повреждений ДНК.
В результате внутриклеточных процессов в ДНК образуются многочисленные АP-сайты из-за спонтанной утраты пуринов/пиримидинов, окисленные участки (например, 8-оксигуанин), возникающие под действием токсических радикалов, постоянно генерируемых в процессах метаболизма.
Следует заметить, что все повреждения показаны только схематически, так как любое из них вызывает локальное изменение структуры ДНК вокруг, а разрывы ДНК почти всегда сопровождаются и модификацией прилежащих к ним оснований. Кроме обычно упоминаемых пиримидиновых димеров, под действием УФ облучения образуются и другие повреждения, например, 6,4-фотопродукты. В дальнейшем, описывая различные системы репарации ДНК, мы будем подробно останавливаться на тех типах повреждений, которые они способны исправлять.
3. Многообразие систем репарации ДНК
Системы репарации ДНК крайне разнообразны – от простых одноэтапных (фотореактивация, деалкилирование) до сложнейших, многоэтапных механизмов, контролируемых большим числом генов и, включающих соответственно, большое число белков. Этим репарация принципиально отличается от процессов репликации и рекомбинации, обходящихся существенно более ограниченным набором биохимических реакций. Множественность частично перекрывающихся и дополняющих друг друга систем репарации, даже некоторая их избыточность, повышает надежность защиты генома и расширяет возможности обеспечения его работы в онтогенезе и при различных физиологических условиях. Классификация процессов репарации ДНК строится на тех реакциях, которые являются их основой. Выделяются реакции прямой репарации, системы эксцизионной репарации ДНК, репарация с привлечением рекомбинации, пострепликативная репарация, репарация двунитевых разрывов, репаративный обход повреждения, причем иногда один и тот же тип репарации может иметь несколько разных названий. В результате исследований последних лет эта классификация менялась и уточнялась, что мы в дальнейшем будем учитывать.
4. Прямая репарация ДНК
Самый простой и эффективный путь восстановления поврежденных нуклеотидов в составе ДНК – это прямое обращение тех химических реакций, индуцированных химическими или физическими агентами, результатом которых стало данное повреждение. Такой способ устранения повреждений принято называть прямой репарацией. К сожалению, с помощью реакций этого типа может быть исправлено только ограниченное число повреждений. Реакциями прямой репарации являются фотореактивация, репарация с помощью метилтрансфераз (репарация алкилированного гуанина), прямая вставка пуринов инсертазой и прямое зашивание однонитевых разрывов полинуклеотидлигазой.
4.1. Фотореактивация
Фотореактивация – первый из описанных (еще до появления модели ДНК Уотсона и Крика) процессов репарации, поэтому на истории его открытия мы подробно остановимся. Намек на то, что живые клетки способны выживать после действия летальной дозы УФ облучения появляются в научном сообществе чуть раньше середины 1930-годов. Разразившаяся Вторая мировая война задержала открытия этого механизма ДНК-репарации повреждений, возникающих в результате УФ облучения до конца 1940-х, до независимых одновременных наблюдений Альберта Кельнера (эмигранта из Германии), работавшего в группе Милислава Демерека в лаборатории Колд-Спринг-Харбора, и Ренато Дальбекко из лаборатории Сальватора Лурии в университете Индианы. В 1949 году немецкий генетик Альберт Кельнер, бежавший из гитлеровской Германии в США, обнаружил, что в клетках бактерий и грибов, таких, как стрептомицеты и пенициллы, облученных ультрафиолетовым (УФ) светом, а затем перенесенных на видимый свет, частота мутаций падает, а выживаемость резко возрастает по сравнению с клетками, оставленными после облучения в темноте. Кельнер пришел к выводу, что на свету проходят реакции восстановления и какие-то поврежденные молекулы или части их возвращаются к норме. Нужно подчеркнуть, что в 1949 году большинство генетиков еще не понимали ведущей роли ДНК в наследственности, имели весьма смутные представления о структуре хромосом и даже не знали, что дрожжи и другие грибы являются эукариотами. Поэтому объяснение, данное Кельнером восстановлению повреждений на свету, было по-настоящему пионерским. Макс Дельбрюк, другой эмигрант из Германии, будущий Нобелевский лауреат, подсказал Кельнеру название для описанного им явления – фотореактивация. В том же 1949 году сходный процесс был найден независимо Р. Дальбекко у бактериофагов. Ни Кельнер, ни Дальбекко не занимались изучением повреждений ДНК или их репарацией. Они оба использовали УФ-облучение как экспериментальный инструмент и заметили неожиданно высокие уровни выживаемости, когда клетки или бактериофаги (вирусы бактерий) после УФ-облучения в результате лабораторной ошибки во время их исследований в соответствующих лабораториях оказались на свету. Их старания объяснить эти поразительные наблюдения привели к открытию феномена фотореактивациии, когда полученные при облучении УФ-светом ДНК-повреждения репарируются в реакции со светозависимым ферментом.
Кельнер считается первооткрывателем фотореактивации потому, что именно он пришел к выводу, что за процесс исправления ответственна простая реакция в наследственном аппарате. Этот вывод был сделан еще до постулирования существования ДНК в виде двойных спиралей и признания за ДНК функции наследственных молекул.
В этой истории есть еще одно забавное совпадение. Как раз в то самое время, когда Дальбекко наткнулся на фотореактивацию, Уотсон был аспирантом в той же самой лаборатории Лурии и сам в своем диссертационном исследовании занимался эффектами ионизирующей радиации. Удивительно, что всего через 4 года при открытии Уотсоном и Криком структуры ДНК, ни Дальбекко, ни сам Уотсон даже не подумали о репарации ДНК.
Процесс фотореактивации состоит в том, что тиминовые димеры, возникшие в результате УФ-облучения, разрушаются, и тимины возвращаются к своей исходной форме под действием видимого света. Что же такое представляет собой пиримилиновый димер? В 1960 году голландские ученые Р. Бьюкерс и У. Верендс изучили химию процесса повреждения нуклеиновых кислот УФ-светом и выделили продукт, специфический для данного повреждающего агента. Оказалось, что двойная связь между пятым и шестым атомами углерода в составе пиримидиновых оснований (тимине и питозине в ДНК и цитозине и урациле в РНК) под действием УФ-света может рваться. Атомы остаются связанными одиночной связью, а в результате разрыва другой связи образуются две свободные валентности.

Рисунок 2. Схема фотореактивации
Обычно ДНК в клетках находятся в так называемой В-форме, когда плоскости оснований параллельны друг другу и расстояние между плоскостями равно примерно 3,4 А. Это расстояние оказывается как раз таким, чтобы освободившиеся при УФ-облучении валентности между С5=С6 атомами пиримидпновых оснований, расположенных рядом в цепи ДНК, могли замкнуться друг на друга и сформировать более сложное, так называемое циклобутановое кольцо. Если димеризация произошла в РНК, то могут возникнуть димеры урацила и любого другого пиримидинового основания. Довольно часто употребляют и другой термин для их обозначении – пиримидиновые димеры. В зависимости от того, какие основания соединены в димер, их называют димерами тимина, димерами цитозина или тимин-цитозиновыми димерами.
Сущность процесса фотореактивации, изображенного на рис. 2, заключается в том, что фермент фотолиаза расщепляет вновь образовавшиеся связи между соседними основаниями и восстанавливает нативную структуру ДНК. В 1963 году фотолиаза E. coli была выделена и очищена. Этот белок кодируется геном phr, а линии с мутациями гена phr имеют дефекты световой репарации.
Все описанные к настоящему времени участвующие в фотореактивации белки относятся к семейству фотолиаз\криптохромов. Фотолиазы были найдены у бактерий, дрожжей, дрозофилы, ксенопуса, иглокожих, но не у высших млекопитающих, включая человека. Эти белки являются мономерами с молекулярным весом 55–70 кд, имеющими два нековалентно связанных хроматофора: первый хроматофор это флавин, а второй – фолат (или дезафлавин). Фотолиаза светонезависимым образом (то есть в темноте) связывается с вызванным УФ-облучением повреждением ДНК. Сама же реакции репарации является светозависимой. Фотолиаза может восстанавливать как пиримидиновые димеры, так и (6–4)фотопродукты – другой тип повреждения ДНК, обычно возникающий под действием УФ-облучения. При образовании этого фотопродукта новые связи возникают между 6 и 4 атомами углерода соседних оснований, оно менее устойчиво, чем пиримидиновые димеры, и поэтому не является такой удобной экспериментальной моделью и несколько хуже изучено.
После расщепления связей в поврежденных основаниях и восстановления их формы фотолиаза отходит от ДНК. Прямое восстановление структуры ДНК на этом завершено.
Это единственная пока найденная ферментная реакция, в которой фактором активации служит не химическая энергия, а энергия видимого света. Все остальные типы репарации не требуют активации светом и потому первое время носили собирательное название "темновая репарация". Сейчас этот термин практически не встречается.
У человека белки – гомологи фотолиаз участвуют не в процессах репарации, а в регуляции циркадного ритма. Были клонированы гены hCry1, hCry2, mCry1, mCry2 (human/mouse circadian rythm). Эти гены экспрессируются в самых разных тканях – мозгу, печени, яичках, сетчатке и действуют, как циркадные проторецепторы. Вероятно, они являются антагонистами, так как у мышей дефектных по mCry1 циркадные периоды укорачиваются, а у дефектных по mCry2 – напротив, удлиняются. Гены Cry представляют собой очень интересный пример эволюционного изменения функции репарационных генов.
Кельнер считается первооткрывателем фотореактивации потому, что именно он пришел к выводу, что за процесс исправления ответственна простая реакция в наследственном аппарате. Этот вывод был сделан еще до постулирования существования ДНК в виде двойных спиралей и признания за ДНК функции наследственных молекул.
В этой истории есть еще одно забавное совпадение. Как раз в то самое время, когда Дальбекко наткнулся на фотореактивацию, Уотсон был аспирантом в той же самой лаборатории Лурии и сам в своем диссертационном исследовании занимался эффектами ионизирующей радиации. Удивительно, что всего через 4 года при открытии Уотсоном и Криком структуры ДНК, ни Дальбекко, ни сам Уотсон даже не подумали о репарации ДНК.
Процесс фотореактивации состоит в том, что тиминовые димеры, возникшие в результате УФ-облучения, разрушаются, и тимины возвращаются к своей исходной форме под действием видимого света. Что же такое представляет собой пиримилиновый димер? В 1960 году голландские ученые Р. Бьюкерс и У. Верендс изучили химию процесса повреждения нуклеиновых кислот УФ-светом и выделили продукт, специфический для данного повреждающего агента. Оказалось, что двойная связь между пятым и шестым атомами углерода в составе пиримидиновых оснований (тимине и питозине в ДНК и цитозине и урациле в РНК) под действием УФ-света может рваться. Атомы остаются связанными одиночной связью, а в результате разрыва другой связи образуются две свободные валентности.
Рисунок 2. Схема фотореактивации
Обычно ДНК в клетках находятся в так называемой В-форме, когда плоскости оснований параллельны друг другу и расстояние между плоскостями равно примерно 3,4 А. Это расстояние оказывается как раз таким, чтобы освободившиеся при УФ-облучении валентности между С5=С6 атомами пиримидпновых оснований, расположенных рядом в цепи ДНК, могли замкнуться друг на друга и сформировать более сложное, так называемое циклобутановое кольцо. Если димеризация произошла в РНК, то могут возникнуть димеры урацила и любого другого пиримидинового основания. Довольно часто употребляют и другой термин для их обозначении – пиримидиновые димеры. В зависимости от того, какие основания соединены в димер, их называют димерами тимина, димерами цитозина или тимин-цитозиновыми димерами.
Сущность процесса фотореактивации, изображенного на рис. 2, заключается в том, что фермент фотолиаза расщепляет вновь образовавшиеся связи между соседними основаниями и восстанавливает нативную структуру ДНК. В 1963 году фотолиаза E. coli была выделена и очищена. Этот белок кодируется геном phr, а линии с мутациями гена phr имеют дефекты световой репарации.
Все описанные к настоящему времени участвующие в фотореактивации белки относятся к семейству фотолиаз\криптохромов. Фотолиазы были найдены у бактерий, дрожжей, дрозофилы, ксенопуса, иглокожих, но не у высших млекопитающих, включая человека. Эти белки являются мономерами с молекулярным весом 55–70 кд, имеющими два нековалентно связанных хроматофора: первый хроматофор это флавин, а второй – фолат (или дезафлавин). Фотолиаза светонезависимым образом (то есть в темноте) связывается с вызванным УФ-облучением повреждением ДНК. Сама же реакции репарации является светозависимой. Фотолиаза может восстанавливать как пиримидиновые димеры, так и (6–4)фотопродукты – другой тип повреждения ДНК, обычно возникающий под действием УФ-облучения. При образовании этого фотопродукта новые связи возникают между 6 и 4 атомами углерода соседних оснований, оно менее устойчиво, чем пиримидиновые димеры, и поэтому не является такой удобной экспериментальной моделью и несколько хуже изучено.
После расщепления связей в поврежденных основаниях и восстановления их формы фотолиаза отходит от ДНК. Прямое восстановление структуры ДНК на этом завершено.
Это единственная пока найденная ферментная реакция, в которой фактором активации служит не химическая энергия, а энергия видимого света. Все остальные типы репарации не требуют активации светом и потому первое время носили собирательное название "темновая репарация". Сейчас этот термин практически не встречается.
У человека белки – гомологи фотолиаз участвуют не в процессах репарации, а в регуляции циркадного ритма. Были клонированы гены hCry1, hCry2, mCry1, mCry2 (human/mouse circadian rythm). Эти гены экспрессируются в самых разных тканях – мозгу, печени, яичках, сетчатке и действуют, как циркадные проторецепторы. Вероятно, они являются антагонистами, так как у мышей дефектных по mCry1 циркадные периоды укорачиваются, а у дефектных по mCry2 – напротив, удлиняются. Гены Cry представляют собой очень интересный пример эволюционного изменения функции репарационных генов.
4.2. Репарация О6-алкилированного гуанина
В 1944–1948 годах выдающийся советский генетик И.А. Рапопорт нашел новый класс химических мутагенов, способных добавлять к взаимодействующим с ними молекулам алкильные (метиловые, этиловые, пропиловые, бутиловые) боковые группы, и назвал их алкилирующими агентами. В конце 60-х годов стало ясно, что обработка клеток алкилирующими агентами вызывает, в зависимости от агента N– или О-алкилированние пуринов и пиримидинов, а также трифосфатов. Один из наиболее мощных алкилирующих мутагенов, метил-нитро-нитрозогуанидин, может алкилировать гуанин, присоединяя метальную группу к кислороду, связанному с шестым атомом кольца. Полученный продукт был назван О6-метил-гуанином. В 1978–1979 годах генетики и биохимики обнаружили, что метильная группа может отщепляться от гуанина и тогда происходит прямое восстановление структуры ДНК в этой точке. В 1982–1988 годах было установлено, что такой же механизм функционирует при репарации О4-алкилтимина.
Последующие исследования показали, что в клетках млекопитающих есть целый класс белков метилтрансфераз, которые могут захватывать метальные группы от модифицированного гуанина, переносить их с поврежденного основания на цистеин метилтрансферазы и благодаря этому восстанавливать исходную структуру ДНК. При этом сами метилтрансферазы инактивируются. Например, фермент, кодируемый геном ada (Об-метил-гуанин-трансфераза), распознает Об-метилгуанин в ДНК и удаляет метильную группу, возвращая основание в исходную форму. Репарацией О4-алкилтимина ведает О4-метил-тимин-ДНК-метилтрансфераза. Важно понять, что метилтрансфераза, захватив метильную группу, не может от нее освободиться. Тем самым в прямом смысле метилтрансферазы не являются ферментами, так как настоящие ферменты по определению не изменяются в ходе реакций. Если для каждого акта прямой репарации О6-метилгуанина или О4-алкилтимина нужна новая молекула белка, клетка вынуждена запускать синтез новых его порций. Обычно для обеспечения репарации внутри клетки их накапливается несколько тысяч, по одной молекуле уходит на каждое повреждение. Если процесс возникновения новых повреждений в ДНК идет медленнее, чем синтез новых порций метилтрансфераз, то последних хватает на захват всех метильных групп в гуанинах, и мутации не возникают. Если же скорость внесения новых повреждений превышает скорость синтеза метилтрансфераз, последние перестают справляться со всеми повреждениями, и в клетках накапливаются метилированные основания и предшественники.
В минуту в клетке Е. coli может синтезироваться порядка 100 молекул метилтрансфераз. Следовательно, мутации не возникнут, если скорость возникновения повреждений будет меньше 100 в минуту. Для сравнения: кишечные палочки делятся каждые 30 минут и, таким образом, клетка за один клеточный цикл может накопить не более 3000 метилтрансфераз. У Е. coli устойчивость к алкилирующим агентам связана с 4 генами, ada, alkA, alkB, aidB, но только ген ada вовлечен в репарацию О6-метилгуанина.
Необходимо отметить, что О6-алкилгуанины являются одними из самых значимых повреждений, несмотря на то, что их общее количество среди всех повреждений ДНК, вызванных алкилированием меньше 8 %. В основном это О6-метилгуанин и О6-этилгуанин, основания, способные к неправильному спариванию и являющиеся основной причиной возникновения транзиций GC-AT. К тому же именно эти повреждения не удаляются другими системами репарации, например, эксцизионной репарацией оснований (base excision repair, BER), а при действии другой эксцизионной системы репарации – репарации неспаренных оснований(missmath repair, MMR), приводят к возникновению двунитевых разрывов. Другим столь же серьезным предмутационным алкилированным основанием является появляющийся крайне редко О4-метилтимин, – всего менее 0.4 % среди всех вызванных алкилированием повреждений. То есть в случае двух данных повреждений при инактивации или подавлении их прямого восстановления с участием метилтрансфераз, другие системы репарации окажутся бесссильны.


Рисунок 3. AGT человека (А – схема всего белка, Б – схема его активного центра)
В человеческих клетках О6-алкилированные повреждения могут быть восстановлены в прямой реакции О6-метилгуанин-ДНК– метилтрансферазой (MGMT, ATase, AGT,AGAT), которая является гомологом продукта гена ada (ADA) E.coli, структура которой изображена на рис. 3.
С-концевой домен (остатки 86-207) содержит консервативный активный сайт с цистеиновым мотивом (IPCHRV), канал для связывания 06-алкилгуанина и ДНК-распознающий мотив спираль-поворот-спираль (Н6), работающий благодаря его внедрению в глубокую бороздку ДНК.
Гуанин «втаскивается» внутрь активного канала вдоль распознающей спирали. Гуанин-специфические водородные связи устанавливаются между белком и ДНК.
Человеческий MGMT ген картирован на 10q26 и кодирует белок с молекулярным весом в 24 кД, состоящий из 207 аминокислотных остатков. Этот белок переносит метил или хлорэтил из поврежденного гуанина на остаток цистенина в своем активном центре. Активность MGMT была измерена в различных тканях человека, как нормальных, так и опухолевых и было показано, что его экспрессия, особенно в опухолевых тканях, может быть различной. Клетки, дефектные по этому гену, не способны к репарации О6-алкилгуанина, в них резко повышен уровень мутаций, вызываемых алкилирующими агентами, сестринских хроматидных обменов и хромосомных аберраций. Мыши, нокаутные по MGMT жизнеспособны, но у них резко повышено количество спонтанно возникающих опухолей и они чувствительны к действию алкилирующих агентов. Напротив, у мышей с оверэкспрессией MGMT спонтанные опухоли развиваются значительно реже, чем у контрольных. MGMT был первым геном млекопитающих, у которого была показана экспрессия, индуцированная действием генотоксического стресса и глюкокортикоидов, приводящая к адаптивному ответу клетки на мутагенное и токсическое действие простых алкилирующих агентов.
Экспрессия MGMT регулируется метилированием как самого гена, так и его промотора, причем метилирование промотора приводит к ее ингибированию, а метилирование самого гена к повышению экспрессии MGMT. С метилированием MGMT также связана повышенная устойчивость клеток меланомы к действию хлорэтиловых антиопухолевых препаратов.
Последующие исследования показали, что в клетках млекопитающих есть целый класс белков метилтрансфераз, которые могут захватывать метальные группы от модифицированного гуанина, переносить их с поврежденного основания на цистеин метилтрансферазы и благодаря этому восстанавливать исходную структуру ДНК. При этом сами метилтрансферазы инактивируются. Например, фермент, кодируемый геном ada (Об-метил-гуанин-трансфераза), распознает Об-метилгуанин в ДНК и удаляет метильную группу, возвращая основание в исходную форму. Репарацией О4-алкилтимина ведает О4-метил-тимин-ДНК-метилтрансфераза. Важно понять, что метилтрансфераза, захватив метильную группу, не может от нее освободиться. Тем самым в прямом смысле метилтрансферазы не являются ферментами, так как настоящие ферменты по определению не изменяются в ходе реакций. Если для каждого акта прямой репарации О6-метилгуанина или О4-алкилтимина нужна новая молекула белка, клетка вынуждена запускать синтез новых его порций. Обычно для обеспечения репарации внутри клетки их накапливается несколько тысяч, по одной молекуле уходит на каждое повреждение. Если процесс возникновения новых повреждений в ДНК идет медленнее, чем синтез новых порций метилтрансфераз, то последних хватает на захват всех метильных групп в гуанинах, и мутации не возникают. Если же скорость внесения новых повреждений превышает скорость синтеза метилтрансфераз, последние перестают справляться со всеми повреждениями, и в клетках накапливаются метилированные основания и предшественники.
В минуту в клетке Е. coli может синтезироваться порядка 100 молекул метилтрансфераз. Следовательно, мутации не возникнут, если скорость возникновения повреждений будет меньше 100 в минуту. Для сравнения: кишечные палочки делятся каждые 30 минут и, таким образом, клетка за один клеточный цикл может накопить не более 3000 метилтрансфераз. У Е. coli устойчивость к алкилирующим агентам связана с 4 генами, ada, alkA, alkB, aidB, но только ген ada вовлечен в репарацию О6-метилгуанина.
Необходимо отметить, что О6-алкилгуанины являются одними из самых значимых повреждений, несмотря на то, что их общее количество среди всех повреждений ДНК, вызванных алкилированием меньше 8 %. В основном это О6-метилгуанин и О6-этилгуанин, основания, способные к неправильному спариванию и являющиеся основной причиной возникновения транзиций GC-AT. К тому же именно эти повреждения не удаляются другими системами репарации, например, эксцизионной репарацией оснований (base excision repair, BER), а при действии другой эксцизионной системы репарации – репарации неспаренных оснований(missmath repair, MMR), приводят к возникновению двунитевых разрывов. Другим столь же серьезным предмутационным алкилированным основанием является появляющийся крайне редко О4-метилтимин, – всего менее 0.4 % среди всех вызванных алкилированием повреждений. То есть в случае двух данных повреждений при инактивации или подавлении их прямого восстановления с участием метилтрансфераз, другие системы репарации окажутся бесссильны.
Рисунок 3. AGT человека (А – схема всего белка, Б – схема его активного центра)
В человеческих клетках О6-алкилированные повреждения могут быть восстановлены в прямой реакции О6-метилгуанин-ДНК– метилтрансферазой (MGMT, ATase, AGT,AGAT), которая является гомологом продукта гена ada (ADA) E.coli, структура которой изображена на рис. 3.
С-концевой домен (остатки 86-207) содержит консервативный активный сайт с цистеиновым мотивом (IPCHRV), канал для связывания 06-алкилгуанина и ДНК-распознающий мотив спираль-поворот-спираль (Н6), работающий благодаря его внедрению в глубокую бороздку ДНК.
Гуанин «втаскивается» внутрь активного канала вдоль распознающей спирали. Гуанин-специфические водородные связи устанавливаются между белком и ДНК.
Человеческий MGMT ген картирован на 10q26 и кодирует белок с молекулярным весом в 24 кД, состоящий из 207 аминокислотных остатков. Этот белок переносит метил или хлорэтил из поврежденного гуанина на остаток цистенина в своем активном центре. Активность MGMT была измерена в различных тканях человека, как нормальных, так и опухолевых и было показано, что его экспрессия, особенно в опухолевых тканях, может быть различной. Клетки, дефектные по этому гену, не способны к репарации О6-алкилгуанина, в них резко повышен уровень мутаций, вызываемых алкилирующими агентами, сестринских хроматидных обменов и хромосомных аберраций. Мыши, нокаутные по MGMT жизнеспособны, но у них резко повышено количество спонтанно возникающих опухолей и они чувствительны к действию алкилирующих агентов. Напротив, у мышей с оверэкспрессией MGMT спонтанные опухоли развиваются значительно реже, чем у контрольных. MGMT был первым геном млекопитающих, у которого была показана экспрессия, индуцированная действием генотоксического стресса и глюкокортикоидов, приводящая к адаптивному ответу клетки на мутагенное и токсическое действие простых алкилирующих агентов.
Экспрессия MGMT регулируется метилированием как самого гена, так и его промотора, причем метилирование промотора приводит к ее ингибированию, а метилирование самого гена к повышению экспрессии MGMT. С метилированием MGMT также связана повышенная устойчивость клеток меланомы к действию хлорэтиловых антиопухолевых препаратов.
4.3. Репарация однонитевых разрывов ДНК
Еще один тип реакций прямой репарации был обнаружен для однонитевых разрывов ДНК, индуцируемых, например, ионизирующим излучением. При этом с помощью фермента ДНК полинуклеотидлигазы (от англ. ligase – соединять, связывать) происходит прямое воссоединение разорванных концов в молекуле ДНК. Здесь у нас есть возможность немного подробнее остановиться на роли лигаз в клетке. ДНК лигазы являются важными ферментами метаболизма ДНК. Они катализируют реакцию объединения концов ДНК, необходимую для репликации ДНК и для тех путей репарации, при которых есть репаративный синтез. У большинства организмов лигазы используют энергию АТФ, и лишь у эубактерий – энергию НАД(+). Интересно, что несмотря на различия в аминокислотных последовательностях и биохимических реакциях между этими двумя классами лигаз, структура аденилирующего домена у них совершенно одинакова.
Полинуклеотидлигаза является основным ферментом у E.coli, а высшие организмы производят несколько различных лигаз, имеющих специфические мишени и функции. Лигаза I необходима для сшивания фрагментов Оказаки и некоторых репарационных реакций, лигаза II не является отдельным самостоятельным ферментом, а появляется как продукт деградации лигазы III, ДНК лигаза III имеет несколько изоформ, вовлеченных в процессы репарации и рекомбинации, а ДНК лигаза IV необходима для V(D)J рекомбинации и негомологичного соединения концов ДНК при воссоединении двунитевых разрывов. Изучение аминокислотной последовательности и структурный анализ показали, что все лигазы построены вокруг общего каталитического кора, подобного таковому у лигазы бактериофага Т-7, структура которого до сих пор недостаточно ясна.
Полинуклеотидлигаза является основным ферментом у E.coli, а высшие организмы производят несколько различных лигаз, имеющих специфические мишени и функции. Лигаза I необходима для сшивания фрагментов Оказаки и некоторых репарационных реакций, лигаза II не является отдельным самостоятельным ферментом, а появляется как продукт деградации лигазы III, ДНК лигаза III имеет несколько изоформ, вовлеченных в процессы репарации и рекомбинации, а ДНК лигаза IV необходима для V(D)J рекомбинации и негомологичного соединения концов ДНК при воссоединении двунитевых разрывов. Изучение аминокислотной последовательности и структурный анализ показали, что все лигазы построены вокруг общего каталитического кора, подобного таковому у лигазы бактериофага Т-7, структура которого до сих пор недостаточно ясна.
4.4. Репарация АР-сайтов за счет прямой вставки пуринов
Голландский ученый Т. Линдал в 1979 году нашел, что при некоторых типах повреждений пуриновых оснований ковалентная связь между основанием и сахаром (гликозидная связь) может рваться. Тогда в молекуле ДНК на месте этих оснований образуется брешь, названная АР-сайтом. Термин приложим также к случаям, когда из ДНК выпадают пиримидиновые основания (термин АР-сайт, таким образом, объединяет все случаи выщепления оснований с образованием и апуриновых и апиримидиновых сайтов). Описаны ферменты, названные инсертазами (insert – вставлять), которые могут вставлять в брешь такое же основание, какое было до повреждения, и соединять его с сахаром. Например, ДНК-пурин-инсертаза, которая катализирует образование N-гликозидной связи между 1-атомом дезоксирибозы апурин/апиримидинового сайта (АР-сайта), образовавшегося напротив пиримидина, и комплементарным ему основанием. Структура ДНК приобретает исходный неповрежденный вид. Однако только ограниченное число АР-сайтов может быть исправлено с помощью этого типа реакции.
5. Эксцизионная репарация
Существуют более сложные реакции восстановления, напоминающие хирургические вмешательства в структуру ДНК, когда поврежденные участки вырезаются из цепи ДНК (отсюда происходит и термин "эксцизионная репарация", от excision – вырезание), а затем образовавшиеся бреши заполняются неповрежденным материалом. Все типы эксцизионной репарации имеют общие этапы:
1. Распознавание повреждения.
2. Надрезание нити ДНК (сахарофосфатного остова).
3. Эксцизия участка, содержащего повреждение.
4. Репаративный синтез на неповрежденной матрице и лигирование.
Три основных типа эксцизионной репарации получили свои названия в зависимости от того, какие именно повреждения будут исправляться. Эти типы репарации, несмотря на лежащий в их основе общий процесс вырезания участка ДНК с повреждением, принципиально различаются между собой.
1. Распознавание повреждения.
2. Надрезание нити ДНК (сахарофосфатного остова).
3. Эксцизия участка, содержащего повреждение.
4. Репаративный синтез на неповрежденной матрице и лигирование.
Три основных типа эксцизионной репарации получили свои названия в зависимости от того, какие именно повреждения будут исправляться. Эти типы репарации, несмотря на лежащий в их основе общий процесс вырезания участка ДНК с повреждением, принципиально различаются между собой.
5.1. Эксцизионная репарация оснований (base excision repair, BER)
Объектом BER служат неправильно спаренные, алкилированные, окисленные и тому подобное, основания. Как мы уже упоминали ранее, за сутки в каждой клетке человека происходит не менее 105 нуждающихся в коррекции модификаций оснований. Этот тип повреждений не вызывает серьезных изменений в структуре двойной спирали ДНК, приводящих к нарушению репликации ДНК и остановке клеточного цикла, но служит источником мутаций. Механизм BER, возможно, является наиболее древним и важным. Первый этап распознавания поврежденных оснований в этой системе репарации осуществляют специальные белки – гоикозилазы. Это высокоспецифические ферменты, гидролизующие N-гликозидную связь между сахарофосфатным остовом и поврежденным основанием.
ДНК-гликозилазы различаются по своей субстратной специфичности, то есть они способны распознавать только определенные поврежденные основания, а не все подряд. У некоторых из них спектр распознаваемых повреждений достаточно широк, а у некоторых – крайне узок. На рис. 4 приведены различные типы повреждений оснований, которые могут распознаваться и репарироваться системой BER. Обычно определенные повреждения репарируются определенными гликозилазами.
У E.coli к настоящему времени выделено и описано 8, а у человека – 11 различных гликозилаз. В табл. 2 приведены названия 11 описанных к настоящему времени гликозилаз человека и указана специфичность распознавания ими модифицированных или неправильно спаренных оснований. Обратите внимание, что ни одна из гликозилаз не распознает О6-МеG.
Гликозилазы бывают двух типов – 1 типа убирают измененное основание, оставляя в цепи АР-сайт, а 2 типа сразу же надрезают после удаления основания АР-сайт с помощью эндогенной 3’-эндонуклеазы, оставляя после себя однонитевой разрыв. В таком случае принято указывать, что ДНК-гликозилазы 2 типа обладают не только гликозилазной, но и АР-лиазной активностью.
ДНК-гликозилазы различаются по своей субстратной специфичности, то есть они способны распознавать только определенные поврежденные основания, а не все подряд. У некоторых из них спектр распознаваемых повреждений достаточно широк, а у некоторых – крайне узок. На рис. 4 приведены различные типы повреждений оснований, которые могут распознаваться и репарироваться системой BER. Обычно определенные повреждения репарируются определенными гликозилазами.
У E.coli к настоящему времени выделено и описано 8, а у человека – 11 различных гликозилаз. В табл. 2 приведены названия 11 описанных к настоящему времени гликозилаз человека и указана специфичность распознавания ими модифицированных или неправильно спаренных оснований. Обратите внимание, что ни одна из гликозилаз не распознает О6-МеG.
Гликозилазы бывают двух типов – 1 типа убирают измененное основание, оставляя в цепи АР-сайт, а 2 типа сразу же надрезают после удаления основания АР-сайт с помощью эндогенной 3’-эндонуклеазы, оставляя после себя однонитевой разрыв. В таком случае принято указывать, что ДНК-гликозилазы 2 типа обладают не только гликозилазной, но и АР-лиазной активностью.