Страница:
Однако видение и доказанное знание — это разные вещи; именно здесь проходит граница между искусством и наукой. Учёного, ищущего доказательств, великий ясновидец слишком легко считает «несчастнейшим из смертных» — и наоборот, использование непосредственного восприятия в качестве источника познания кажется учёному-аналитику в высшей степени подозрительным. В исследовании поведения существует даже школа — ортодоксальный американский бихевиоризм, — которая всерьёз пытается исключить из своей методики непосредственное наблюдение животных. Право же, стоит потрудиться ради того, чтобы доказать «незрячим», но разумным людям все то, что мы увидели; доказать так, чтобы им пришлось поверить, чтобы каждый поверил!
С другой стороны, статистический анализ может обратить наше внимание на несоответствия, до сих пор ускользавшие от нашего образного восприятия. Оно устроено так, что раскрывает закономерности и потому всегда все видит более красивым и правильным, чем на самом деле.
Решение проблемы, предлагаемое нам восприятием, часто носит характер хотя и очень «элегантной», но слишком уж упрощённой рабочей гипотезы. Как раз в случае исследования мотиваций рациональному анализу нередко удаётся придраться к образному восприятию и уличить его в ошибках.
В большей части всех проведённых до сих пор мотивационных анализов исследовались поведенческие акты, в которых принимают участие лишь два взаимно соперничающих инстинкта, причём, как правило, два из «большой четвёрки» (голод, любовь, бегство и агрессия). При изучении конфликтов между побуждениями, сознательный выбор простейших по возможности случаев вполне оправдывается нынешним скромным уровнем наших знаний. Точно так же правы были классики этологии, когда ограничивались лишь теми случаями, в которых животное находится под влиянием одного-единственного побуждения. Но мы должны ясно понимать, что поведение, определяемое только двумя компонентами побуждений, — это поистине редкость; оно встречается лишь немногим чаще, чем такое, которое вызывается только одним инстинктом, действующим без всяких помех.
Поэтому, при поисках подходящего объекта для образцово точного мотивационного анализа правильно поступает тот, кто выбирает поведение, о котором с некоторой достоверностью известно, что в нем принимают участие только два инстинкта одинакового веса. Иногда для этого можно использовать технический трюк, как это сделала моя сотрудница Хельга Фишер, проводя мотивационный анализ угрозы у серых гусей. Оказалось, что на родном озере наших гусей, Эсс-зее, взаимодействие агрессии и бегства в чистом виде изучать невозможно, так как в выразительных движениях птиц там «высказывается» слишком много других мотиваций, прежде всего сексуальных. Но несколько случайных наблюдений показали, что голос сексуальности почти совсем замолкает, если гуси находятся в незнакомом месте. Тогда они ведут себя примерно так же, как перелётная стая в пути: держатся гораздо теснее, становятся гораздо пугливее, и в своих социальных конфликтах позволяют наблюдать проявления обоих исследуемых инстинктов в более чистых формах. Учитывая все это, Фишер с помощью дрессировки кормом сумела научить наших гусей «по приказу» выходить на чужую для них местность, которую она выбирала за оградой Института, и пастись там. Затем из гусей, каждый из которых, разумеется, известен по сочетанию разноцветных колец, выбирался какой-то один — как правило, гусак, — и в течение долгого времени наблюдались его агрессивные столкновения с товарищами по стаду, причём регистрировались все замеченные выразительные движения угрозы. А поскольку из предыдущих многолетних наблюдений за этим стадом были во всех подробностях известны отношения между отдельными птицами в смысле иерархии и силы — особенно среди старых гусаков высоких рангов, — здесь представлялась особенно хорошая возможность точного анализа ситуаций. Анализ движений и регистрация последующего поведения происходили следующим образом. Хельга Фишер постоянно имела при себе приведённую здесь «таблицу образцов», которую составил художник нашего Института Герман Кахер на основании точно запротоколированных случаев угрозы, так что в каждом конкретном случае ей приходилось лишь продиктовать: «Макс сделал D Гермесу, который пасся и медленно приближался к нему; Гермес ответил Е, на что Макс ответил F''». Серия иллюстраций приводит настолько тонкие различия угрожающих жестов, что лишь в исключительных случаях приходилось обозначать замеченную позу как D-Е или К-L, если нужно было описать промежуточную форму.
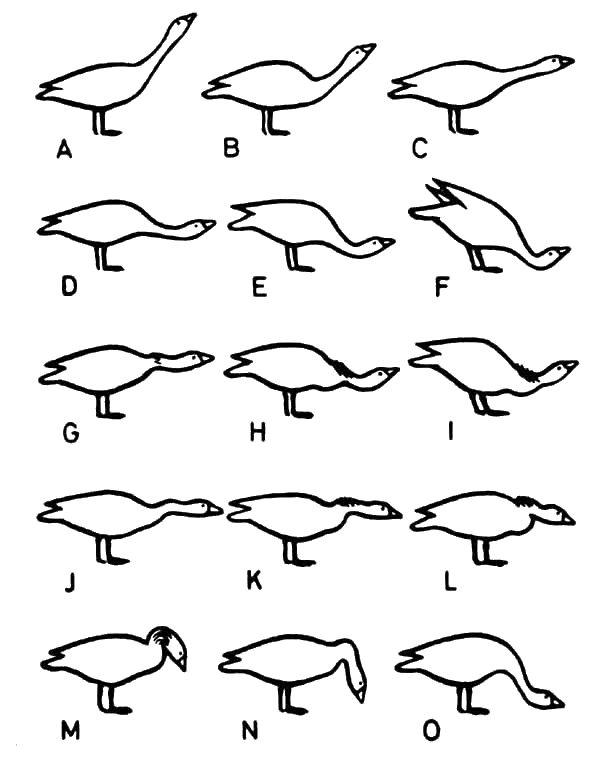
Даже при этих условиях, почти идеальных для «чистой культуры» двух мотиваций, иногда появлялись движения, которые нельзя было объяснить только взаимодействием этих двух побуждений. Про угрожающие движения А и В, когда шея вытянута вперёд и вверх, мы знаем, что на оба побуждения накладывается независимое третье — стремление к охранному наблюдению с поднятой головой. Различия между рядами А — С и D — F, в каждом из которых представлено возрастание слева направо социального страха на фоне примерно равной агрессивности, состоит, по-видимому, лишь в разной интенсивности обоих побуждений.
Напротив, в отношении форм М-О совершенно ясно, что в них принимает участие ещё какая-то мотивация, природа которой пока не выяснена.
Как уже сказано, отыскивать в качестве объектов мотивационного анализа такие случаи, ще принимают участие только два источника побуждений, — это, безусловно, правильная стратегия исследований. Однако даже при таких благоприятных условиях необходимо внимательно и постоянно высматривать элементы движений, которые нельзя объяснить лишь соперничеством этих двух побуждений.
Перед началом любого такого анализа нужно ответить на первый и основной вопрос: сколько мотиваций принимают участие в данном действии и какие именно. Для решения этой задачи многие учёные, как например П. Випкема, в последнее время с успехом применяли точные методы факторного анализа.
Изящный пример мотивационного анализа, в котором с самого начала нужно было принимать в расчёт три главных компонента, представила в своей докторской диссертации моя ученица Беатриса Элерт. Предметом исследования было поведение некоторых цихлид при встрече двух незнакомых особей. Выбирались такие виды, у которых самцы и самки почти не отличаются внешне, и именно поэтому два незнакомца всегда реагируют друг на друга действиями, которые мотивируются одновременно бегством, агрессией и сексуальностью. У этих рыб движения, обусловленные каждым отдельным источником мотивации, различаются особенно легко, потому что даже при самой малой интенсивности их характеризует разная ориентация в пространстве. Все сексуально мотивированные действия — копание ямки под гнездо, очистка гнёзда, само выметывание икры и её осеменение — направлены в сторону дна; все движения бегства, даже малейшие намёки на них, направлены прочь от противника и, большей частью, одновременно к поверхности воды, а все движения агрессии — за исключением некоторых угрожающих движений, в какой-то степени «отягощённых бегством», — ориентированы в обратном направлении. Если знать эти общие правила и вдобавок специальную мотивацию некоторых ритуализованных выразительных движений, то у этих рыб можно очень точно установить соотношение, в котором находятся названные инстинкты, определяя их поведение в данный момент. Здесь помогает ещё и то, что многие из них в сексуальном, агрессивном или боязливом настроении наряжаются в разные характерные цвета.
Этот мотивационный анализ дал неожиданный побочный результат — Беатриса Элерт открыла механизм взаимного распознавания полов, который имеется, конечно, не только у этих рыб, но и у очень многих других позвоночных. У исследованных рыб самка и самец не только внешне похожи друг на друга; их движения, даже при половом акте — при выметывании икры и её осеменении — совпадают до мельчайших деталей. Поэтому до сих пор было совершенно загадочно, что же в поведении этих животных препятствует у них возникновению однополых пар. К важнейшим требованиям, какие предъявляются наблюдательности этолога, относится и то, что он должен заметить, если какое-либо широко распространённое действие у определённого животного, или группы животных, не встречается.
Например, у птиц и у рептилий отсутствует координация широкого открывания пасти с одновременным глубоким вдохом — то, что мы называем зевотой, (?) — и это таксономически важный факт, которого никто до Хейнрота не заметил. Можно привести и другие подобные примеры.
Поэтому открытие, что разнополые пары у цихлид возникают благодаря отсутствию одних элементов поведения у самцов и других у самок — это поистине шедевр точного наблюдения. У рыб, о которых идёт речь, сочетаемость трех главных инстинктов — агрессии, бегства и сексуальности — у самцов и у самок различна: у самцов не бывает смеси мотиваций бегства и сексуальности. Если самец хоть чуточку боится своего партнёра, то его сексуальность выключается полностью. У самок то же соотношение между сексуальностью и агрессивностью: если дама не настолько «уважает» своего партнёра, чтобы её агрессивность была полностью подавлена, она попросту не в состоянии проявить по отношению к нему сексуальную реакцию. Она превращается в Брунгильду и нападает на него тем яростнее, чем более готова была бы к сексуальной реакции, т.е. чем ближе она к икрометанию в смысле состояния овариев и уровня выделения гормонов. У самца, напротив, агрессия прекрасно уживается с сексуальностью: он может грубейшим образом нападать на свою невесту, гонять её по всему аквариуму, но при этом демонстрирует и чисто сексуальные движения, и все смешанные, какие только можно себе представить. Самка может очень бояться самца, но её сексуально мотивированных действий это не подавляет. Она может совершенно всерьёз удирать от самца, но при каждой передышке, какую даёт ей этот грубиян, будет выполнять сексуально-мотивированные брачные движения. Именно такие смешанные формы действий, обусловленные бегством и сексуальностью, превратились посредством ритуализации в те широко распространённые церемонии, которые принято называть «чопорным» поведением и которые имеют совершенно определённый смысл.
Из-за различных соотношений сочетаемости между тремя источниками побуждений у разных полов, самец может спариваться только с партнёром низшего ранга, которого он может запугать, а самка — наоборот — лишь с партнёром высшего ранга, который может запугать её; тем самым описанный механизм поведения обеспечивает создание разнополых пар. В различных вариантах, видоизменённый различными процессами ритуализации, этот способ распознавания пола играет важную роль у очень многих позвоночных, вплоть до человека. В то же время это впечатляющий пример того, какие задачи, необходимые для сохранения вида, может выполнять агрессия в гармоничном взаимодействии с другими мотивациями. В 3-й главе мы ещё не могли говорить об этом, поскольку недостаточно знали о парламентской борьбе инстинктов. Кроме того, мы видим на этом примере, насколько различны могут быть соотношения «главных» инстинктов даже у самца и самки одного и того же вида: два мотива, которые у одного пола практические не мешают друг другу и сочетаются в любых соотношениях, у другого взаимно выключаются по принципу триггера.
Как уже пояснялось, «большая четвёрка» отнюдь не всегда поставляет главную мотивацию поведения животного, а тем более человека. И совершенно неправильно полагать, будто между одним из «главных», древних инстинктов и более специальным, эволюционно более молодым инстинктом всегда существует отношение доминирования, в том смысле, что второй выключается первым. Механизмы поведения, которые, вне всяких сомнений, возникли «совсем недавно» — например, социальные инстинкты у общественных животных, обеспечивающие постоянное сохранение стаи, — у многих видов подчиняют отдельную особь настолько, что при определённых обстоятельствах могут заглушить все остальные побуждения. Овцы, прыгающие в пропасть за вожаком-бараном, вошли в пословицу! Серый гусь, отставший от стаи, делает все возможное, чтобы вновь её обрести, и стадный инстинкт может даже пересилить стремление к бегству; дикие серые гуси неоднократно присоединялись к нашим приручённым — в непосредственной близости к людскому жилью — и оставались!. Кто знает, насколько пугливы дикие гуси, тому эти случаи дадут представление о силе их «стадного инстинкта». То же справедливо для очень многих общественных животных вплоть до шимпанзе, о которых Йеркс справедливо заметил: «Один шимпанзе — вообще не шимпанзе».
Даже те инстинкты, которые «только что» (с точки зрения филогенеза) приобрели самостоятельность через ритуализацию и, как я постарался показать в предыдущей главе, получили место и голос в Великом Парламенте Инстинктов в качестве самых молодых депутатов, — даже они при соответствующих обстоятельствах могут заглушить всех своих оппонентов точно так же, как Голод и Любовь. В триумфальном крике гусей мы увидим церемонию, которая управляет жизнью этих птиц больше, чем любой другой инстинкт. С другой стороны, разумеется, существует сколько угодно ритуализованных действий, которые ещё едва обособились от своего неритуализованного прототипа; их скромное влияние на общее поведение состоит лишь в том, что «желательная» для них координация движений — как мы видели в случае натравливания у огарей — становится в какой-то мере предпочтительной и используется чаще, чем другие, тоже возможные формы.
«Сильный» или «слабый» голос имеет ритуализованное действие в общем концерте инстинктов — оно во всех случаях чрезвычайно затрудняет любой мотивационный анализ, потому что может симулировать поведение, вытекающее из нескольких независимых побуждений. В предыдущей главе мы говорили, что ритуализованное действие, сплавленное в некоторую общность из различных компонентов, копирует форму последовательности движений, которая не является наследственно закреплённой и часто возникает из конфликта нескольких побуждений, как это видно на примере натравливания уток. А поскольку, как уже говорилось там же, копия и оригинал по большей части накладываются друг на друга в одном и том же движении, то чрезвычайно трудно разобраться, сколько же в нем от копии, а сколько от оригинала. Только когда один из первоначально независимых компонентов оказывается в противоречии с ритуально закреплённой координацией, — как направление на «врага», которому адресована угроза в случае натравливания, — тогда становится явным участие новых независимых переменных.
«Танец зигзага» у самцов колюшки, на котором Ян ван Йерсель провёл самый первый эксперимент мотивационного анализа, служит прекрасным примером того, как совсем «слабый» ритуал может вкрасться в конфликт двух «главных» инстинктов в качестве едва заметной третьей величины. Ван-Йерсель заметил, что замечательный танец зигзага, который половозрелые самцы, имеющие свой участок, исполняют перед каждой проплывающей мимо самкой, и который поэтому до тех пор считался просто «ухаживанием», — от случая к случаю выглядит совершенно по-разному. Оказалось, что иногда сильнее подчеркнут «зиг» в сторону самки, а иногда «заг» прочь от неё. Если это последнее движение очень явственно, то становится очевидным, что «заг» направлен в сторону гнёзда. В одном из предельных случаев самец при виде плывущей мимо самки быстро подплывает к ней, тормозит, разворачивается — особенно если самка тотчас поставит ему своё распухшее брюшко — и плывёт назад к входу в гнездо, которое затем показывает самке посредством определённой церемонии (ложась плоско на бок). В другом предельном случае, особенно частом если самка ещё не совсем готова к нересту, за первым «зигом» вообще не следует никакого «зага», а вместо того — нападение на самку.
Из этих наблюдений ван Йерсель правильно заключил, что «зиг» в сторону самки мотивируется агрессивным инстинктом, а «заг» в сторону гнёзда — сексуальным; и ему удалось экспериментально доказать правильность этого заключения. Он изобрёл методы, с помощью которых мог точно измерять силу агрессивного и сексуального инстинктов у каждого данного самца. Самцу предлагались макеты соперника стандартизованных размеров и регистрировалась интенсивность и продолжительность боевой реакции.
Сексуальный инстинкт измерялся с помощью макетов самки, которые внезапно убирались через определённое время.
В этих случаях самец «разряжает» внезапно заблокированный сексуальный инстинкт, совершая действия ухаживания за потомством, т.е. обмахивая плавниками как бы икру или мальков в гнезде; и продолжительность этого «заменяющего обмахивания» даёт надёжную меру сексуальной мотивации. Ван Йерсель научился предсказывать по результатам таких измерений, как будет выглядеть танец зигзага у данного самца, — и наоборот, по наблюдаемой форме танца заранее оценивать соотношения обоих инстинктов и результаты будущих измерений.
Но кроме обоих главных побуждений, определяющих движения самца колюшки в общих чертах, — на них оказывает влияние ещё какое-то третье, хоть и более слабое.
Это знаток ритуализованного поведения заподозрит сразу же, увидев ритмическую правильность смены «зигов» и «загов». Попеременное преобладание одного из двух противоречивых побуждений вряд ли может привести к столь регулярной смене направлений, если здесь не вступает в игру новая, ритуализованная координация. Без неё короткие рывки в разных направлениях следуют друг за другом с типичной случайностью, как это бывает у людей в состоянии крайней растерянности. Ритуализованное движение, напротив, всегда имеет тенденцию к ритмическому повторению в точности одинаковых элементов. Мы говорили об этом в связи с действенностью сигнала.
Подозрение, что здесь замешана ритуализация, превращается в уверенность, когда мы видим, как танцующий самец при своих «загах» временами, кажется, совершенно забывает, что они сексуально мотивированы и должны указывать точно на гнездо. Вместо этого он рисует вокруг самки очень красивый и правильный зубчатый венец, в котором каждый «зиг» направлен точно в сторону самки, а каждый «заг» — точно от неё. Как ни очевидна относительная слабость новой координации движений, стремящейся превратить «зиги» и «заги» в ритмический «зигзаг», — она может, однако, решающим образом определить регулярность последовательных проявлений обеих главных мотиваций.
Вторая важная функция, которую ритуализованная координация может, очевидно, выполнять, даже будучи очень слабой в других отношениях, — это изменение направления неритуализованных движений, лежавших в основе ритуала и происходивших из других побуждений. Примеры этого мы уже видели при обсуждении классического образца ритуала, а именно — при натравливании селезня уткой.
В 5-й главе, где речь шла о процессе ритуализации, я старался показать, как этот процесс, причины которого все ещё весьма загадочны, создаёт совершенно новые инстинкты, диктующие организму свои собственные «Ты должен…» так же категорично, как и любой из, казалось бы, единовластных «больших» инстинктов голода, страха или любви. В предыдущей 6-й главе я пытался решить ещё более трудную задачу: коротко и доступно показать, как происходит взаимодействие между различными автономными инстинктами, каким общим правилам подчиняются эти взаимодействия, и какими способами можно — несмотря на все сложности — получить некоторое представление о структуре взаимодействий в таком поведении, которое определяется несколькими соперничающими побуждениями.
Я тёшу себя надеждой, быть может обманчивой, что решить предыдущие задачи мне удалось, и что я могу не только обобщить сказанное в двух последних главах, но и применить полученные в них результаты к вопросу, которым мы займёмся теперь: каким образом ритуал выполняет поистине невыполнимую задачу — удерживает внутривидовую агрессию от всех проявлений, которые могли бы серьёзно повредить сохранению вида, но при этом не выключает её функций, необходимых для сохранения вида! Часть предыдущей фразы, выделенная курсивом, уже отвечает на вопрос, — он кажется очевидным, но вытекает из совершеннейшего непонимания сущности агрессии, — почему у тех животных, для которых тесная совместная жизнь является преимуществом, агрессия попросту не запрещена? Именно потому, что её функции, рассмотренные нами в 3-й главе, необходимы!
Решение проблем, возникающих таким образом перед обоими конструкторами эволюции, достигается всегда одним и тем же способом. Полезный, необходимый инстинкт — вообще остаётся неизменным; но для особых случаев, где его проявление было бы вредно, вводится специально созданный механизм торможения. И здесь снова культурно-историческое развитие народов происходит аналогичным образом; именно потому важнейшие требования Моисеевых и всех прочих скрижалей — это не предтисания, а запреты. Нам ещё придётся подробнее говоьрить о том, о чем здесь лишь предварительно упомянем:
передаваемые традицией и привычно выполняемые табу имеют какое-то отношение к разумной морали — в понимании Иммануила Канта — разве что у вдохновенного законодателя, но никак не у его верующих последователей.
Как врождённые механизмы и ритуалы, препятствующие асоциальному поведению животных, так и человеческие табу определяют поведение, аналогичное истинно моральному лишь с функциональной точки зрения; во всем остальном оно так же далеко от морали, как животное от человека! Но даже постигая сущность этих движущих мотивов, нельзя не восхищаться снова и снова при виде работы физиологических механизмов, которые побуждают животных к самоотверженному поведению, направленному на благо сообщества, как это предписывают нам, людям, законы морали.
Впечатляющий пример такого поведения, аналогичного человеческой морали, являют так называемые турнирные бои. Вся их организация направлена на то, чтобы выполнить важнейшую задачу поединка — определить, кто сильнее, — не причинив серьёзного вреда более слабому. Поскольку рыцарский турнир или спортивное состязание имеют ту же цель, то все турнирные бои неизбежно производят даже на знающих людей впечатление «рыцарственности», или «спортивного благородства». Среди цихлид есть вид, Cichlasoma biocellatum, который именно из-за этого приобрёл своё название, широко распространённое у американских любителей: у них эта рыбка называется «Джек Дэмпси» по имени боксёра, чемпиона мира, который прославился своим безупречным поведением на ринге.
О турнирных боях рыб и, в частности, о процессах ритуализации, которые привели к ним от первоначальных подлинных боев, мы знаем довольно много. Почти у всех костистых рыб настоящей схватке предшествуют угрожающие позы, которые, как уже говорилось, всегда вытекают из конфликта между стремлениями напасть и бежать.
Среди этих поз особенно заметна как специальный ритуал так называемая демонстрация развёрнутого бока, которая первоначально наверняка возникла за счёт того, что рыба под влиянием испуга отворачивается от противника и одновременно, готовясь к бегству, разворачивает вертикальные плавники. Но поскольку при этих движениях противнику предъявляется контур тела максимально возможных размеров, то из них — путём мимического утрирования при добавочных изменениях морфологии плавников — смогла развиться та впечатляющая демонстрация развёрнутого бока, которую знают все аквариумисты, да и не только они, по сиамским бойцовым рыбкам и по другим популярным породам рыб.
В тесной связи с угрозой развёрнутым боком у костистых рыб возник очень широко распространённый запугивающий жест — так называемый удар хвостом. Из позиции развёрнутого бока рыба, напрягая все тело и далеко оттопыривая хвостовой плавник, производит сильный удар хвостом в сторону противника. Хотя сам удар до противника не доходит, но рецепторы давления на его боковой линии воспринимают волну, сила которой, очевидно, сообщает ему о величине и боеспособности его соперника, так же как и размеры контура, видимого при демонстрации развёрнутого бока.
Другая форма угрозы возникла у многих окуневых и у других костистых рыб из заторможенного страхом фронтального удара. В исходной позиции для броска вперёд оба противника изгибают свои тела, словно напряжённые Sобразные пружины, и медленно плывут друг другу навстречу, как можно сильнее топорща жаберные крышки.
Это соответствует разворачиванию плавников при угрозе боком, поскольку увеличивает контур тела, видимый противником. Из фронтальной угрозы у очень многих рыб иногда получается, что оба противника одновременно хватают друг друга за пасть, но — в соответствии с конфликтной ситуацией, из которой возникла сама фронтальная угроза, — они всегда делают это не резко, не ударом, а словно колеблясь, заторможенно. Из этой формы борьбы у некоторых — и у лабиринтовых рыб, лишь отдалённо примыкающих к большой группе окуневых, и у цихлид, типичных представителей окуневых, — возникла интереснейшая ритуализованная борьба, при которой оба соперника в самом буквальном смысле слова «меряются силами», не причиняя друг другу вреда. Они хватают друг друга за челюсть — а у всех видов, для которых характерен этот способ турнирного боя, челюсть покрыта толстым, трудноуязвимым слоем кожи — и тянут изо всех сил. Так возникает состязание, очень похожее на старую борьбу на поясах у швейцарских крестьян, которое может продолжаться по нескольку часов, если встречаются равные противники. У двух в точности равных по силе самцов красивого синего вида широколобых окуньков мы запротоколировали однажды такой поединок, длившийся с 8.30 утра до 2.30 пополудни.
С другой стороны, статистический анализ может обратить наше внимание на несоответствия, до сих пор ускользавшие от нашего образного восприятия. Оно устроено так, что раскрывает закономерности и потому всегда все видит более красивым и правильным, чем на самом деле.
Решение проблемы, предлагаемое нам восприятием, часто носит характер хотя и очень «элегантной», но слишком уж упрощённой рабочей гипотезы. Как раз в случае исследования мотиваций рациональному анализу нередко удаётся придраться к образному восприятию и уличить его в ошибках.
В большей части всех проведённых до сих пор мотивационных анализов исследовались поведенческие акты, в которых принимают участие лишь два взаимно соперничающих инстинкта, причём, как правило, два из «большой четвёрки» (голод, любовь, бегство и агрессия). При изучении конфликтов между побуждениями, сознательный выбор простейших по возможности случаев вполне оправдывается нынешним скромным уровнем наших знаний. Точно так же правы были классики этологии, когда ограничивались лишь теми случаями, в которых животное находится под влиянием одного-единственного побуждения. Но мы должны ясно понимать, что поведение, определяемое только двумя компонентами побуждений, — это поистине редкость; оно встречается лишь немногим чаще, чем такое, которое вызывается только одним инстинктом, действующим без всяких помех.
Поэтому, при поисках подходящего объекта для образцово точного мотивационного анализа правильно поступает тот, кто выбирает поведение, о котором с некоторой достоверностью известно, что в нем принимают участие только два инстинкта одинакового веса. Иногда для этого можно использовать технический трюк, как это сделала моя сотрудница Хельга Фишер, проводя мотивационный анализ угрозы у серых гусей. Оказалось, что на родном озере наших гусей, Эсс-зее, взаимодействие агрессии и бегства в чистом виде изучать невозможно, так как в выразительных движениях птиц там «высказывается» слишком много других мотиваций, прежде всего сексуальных. Но несколько случайных наблюдений показали, что голос сексуальности почти совсем замолкает, если гуси находятся в незнакомом месте. Тогда они ведут себя примерно так же, как перелётная стая в пути: держатся гораздо теснее, становятся гораздо пугливее, и в своих социальных конфликтах позволяют наблюдать проявления обоих исследуемых инстинктов в более чистых формах. Учитывая все это, Фишер с помощью дрессировки кормом сумела научить наших гусей «по приказу» выходить на чужую для них местность, которую она выбирала за оградой Института, и пастись там. Затем из гусей, каждый из которых, разумеется, известен по сочетанию разноцветных колец, выбирался какой-то один — как правило, гусак, — и в течение долгого времени наблюдались его агрессивные столкновения с товарищами по стаду, причём регистрировались все замеченные выразительные движения угрозы. А поскольку из предыдущих многолетних наблюдений за этим стадом были во всех подробностях известны отношения между отдельными птицами в смысле иерархии и силы — особенно среди старых гусаков высоких рангов, — здесь представлялась особенно хорошая возможность точного анализа ситуаций. Анализ движений и регистрация последующего поведения происходили следующим образом. Хельга Фишер постоянно имела при себе приведённую здесь «таблицу образцов», которую составил художник нашего Института Герман Кахер на основании точно запротоколированных случаев угрозы, так что в каждом конкретном случае ей приходилось лишь продиктовать: «Макс сделал D Гермесу, который пасся и медленно приближался к нему; Гермес ответил Е, на что Макс ответил F''». Серия иллюстраций приводит настолько тонкие различия угрожающих жестов, что лишь в исключительных случаях приходилось обозначать замеченную позу как D-Е или К-L, если нужно было описать промежуточную форму.
Даже при этих условиях, почти идеальных для «чистой культуры» двух мотиваций, иногда появлялись движения, которые нельзя было объяснить только взаимодействием этих двух побуждений. Про угрожающие движения А и В, когда шея вытянута вперёд и вверх, мы знаем, что на оба побуждения накладывается независимое третье — стремление к охранному наблюдению с поднятой головой. Различия между рядами А — С и D — F, в каждом из которых представлено возрастание слева направо социального страха на фоне примерно равной агрессивности, состоит, по-видимому, лишь в разной интенсивности обоих побуждений.
Напротив, в отношении форм М-О совершенно ясно, что в них принимает участие ещё какая-то мотивация, природа которой пока не выяснена.
Как уже сказано, отыскивать в качестве объектов мотивационного анализа такие случаи, ще принимают участие только два источника побуждений, — это, безусловно, правильная стратегия исследований. Однако даже при таких благоприятных условиях необходимо внимательно и постоянно высматривать элементы движений, которые нельзя объяснить лишь соперничеством этих двух побуждений.
Перед началом любого такого анализа нужно ответить на первый и основной вопрос: сколько мотиваций принимают участие в данном действии и какие именно. Для решения этой задачи многие учёные, как например П. Випкема, в последнее время с успехом применяли точные методы факторного анализа.
Изящный пример мотивационного анализа, в котором с самого начала нужно было принимать в расчёт три главных компонента, представила в своей докторской диссертации моя ученица Беатриса Элерт. Предметом исследования было поведение некоторых цихлид при встрече двух незнакомых особей. Выбирались такие виды, у которых самцы и самки почти не отличаются внешне, и именно поэтому два незнакомца всегда реагируют друг на друга действиями, которые мотивируются одновременно бегством, агрессией и сексуальностью. У этих рыб движения, обусловленные каждым отдельным источником мотивации, различаются особенно легко, потому что даже при самой малой интенсивности их характеризует разная ориентация в пространстве. Все сексуально мотивированные действия — копание ямки под гнездо, очистка гнёзда, само выметывание икры и её осеменение — направлены в сторону дна; все движения бегства, даже малейшие намёки на них, направлены прочь от противника и, большей частью, одновременно к поверхности воды, а все движения агрессии — за исключением некоторых угрожающих движений, в какой-то степени «отягощённых бегством», — ориентированы в обратном направлении. Если знать эти общие правила и вдобавок специальную мотивацию некоторых ритуализованных выразительных движений, то у этих рыб можно очень точно установить соотношение, в котором находятся названные инстинкты, определяя их поведение в данный момент. Здесь помогает ещё и то, что многие из них в сексуальном, агрессивном или боязливом настроении наряжаются в разные характерные цвета.
Этот мотивационный анализ дал неожиданный побочный результат — Беатриса Элерт открыла механизм взаимного распознавания полов, который имеется, конечно, не только у этих рыб, но и у очень многих других позвоночных. У исследованных рыб самка и самец не только внешне похожи друг на друга; их движения, даже при половом акте — при выметывании икры и её осеменении — совпадают до мельчайших деталей. Поэтому до сих пор было совершенно загадочно, что же в поведении этих животных препятствует у них возникновению однополых пар. К важнейшим требованиям, какие предъявляются наблюдательности этолога, относится и то, что он должен заметить, если какое-либо широко распространённое действие у определённого животного, или группы животных, не встречается.
Например, у птиц и у рептилий отсутствует координация широкого открывания пасти с одновременным глубоким вдохом — то, что мы называем зевотой, (?) — и это таксономически важный факт, которого никто до Хейнрота не заметил. Можно привести и другие подобные примеры.
Поэтому открытие, что разнополые пары у цихлид возникают благодаря отсутствию одних элементов поведения у самцов и других у самок — это поистине шедевр точного наблюдения. У рыб, о которых идёт речь, сочетаемость трех главных инстинктов — агрессии, бегства и сексуальности — у самцов и у самок различна: у самцов не бывает смеси мотиваций бегства и сексуальности. Если самец хоть чуточку боится своего партнёра, то его сексуальность выключается полностью. У самок то же соотношение между сексуальностью и агрессивностью: если дама не настолько «уважает» своего партнёра, чтобы её агрессивность была полностью подавлена, она попросту не в состоянии проявить по отношению к нему сексуальную реакцию. Она превращается в Брунгильду и нападает на него тем яростнее, чем более готова была бы к сексуальной реакции, т.е. чем ближе она к икрометанию в смысле состояния овариев и уровня выделения гормонов. У самца, напротив, агрессия прекрасно уживается с сексуальностью: он может грубейшим образом нападать на свою невесту, гонять её по всему аквариуму, но при этом демонстрирует и чисто сексуальные движения, и все смешанные, какие только можно себе представить. Самка может очень бояться самца, но её сексуально мотивированных действий это не подавляет. Она может совершенно всерьёз удирать от самца, но при каждой передышке, какую даёт ей этот грубиян, будет выполнять сексуально-мотивированные брачные движения. Именно такие смешанные формы действий, обусловленные бегством и сексуальностью, превратились посредством ритуализации в те широко распространённые церемонии, которые принято называть «чопорным» поведением и которые имеют совершенно определённый смысл.
Из-за различных соотношений сочетаемости между тремя источниками побуждений у разных полов, самец может спариваться только с партнёром низшего ранга, которого он может запугать, а самка — наоборот — лишь с партнёром высшего ранга, который может запугать её; тем самым описанный механизм поведения обеспечивает создание разнополых пар. В различных вариантах, видоизменённый различными процессами ритуализации, этот способ распознавания пола играет важную роль у очень многих позвоночных, вплоть до человека. В то же время это впечатляющий пример того, какие задачи, необходимые для сохранения вида, может выполнять агрессия в гармоничном взаимодействии с другими мотивациями. В 3-й главе мы ещё не могли говорить об этом, поскольку недостаточно знали о парламентской борьбе инстинктов. Кроме того, мы видим на этом примере, насколько различны могут быть соотношения «главных» инстинктов даже у самца и самки одного и того же вида: два мотива, которые у одного пола практические не мешают друг другу и сочетаются в любых соотношениях, у другого взаимно выключаются по принципу триггера.
Как уже пояснялось, «большая четвёрка» отнюдь не всегда поставляет главную мотивацию поведения животного, а тем более человека. И совершенно неправильно полагать, будто между одним из «главных», древних инстинктов и более специальным, эволюционно более молодым инстинктом всегда существует отношение доминирования, в том смысле, что второй выключается первым. Механизмы поведения, которые, вне всяких сомнений, возникли «совсем недавно» — например, социальные инстинкты у общественных животных, обеспечивающие постоянное сохранение стаи, — у многих видов подчиняют отдельную особь настолько, что при определённых обстоятельствах могут заглушить все остальные побуждения. Овцы, прыгающие в пропасть за вожаком-бараном, вошли в пословицу! Серый гусь, отставший от стаи, делает все возможное, чтобы вновь её обрести, и стадный инстинкт может даже пересилить стремление к бегству; дикие серые гуси неоднократно присоединялись к нашим приручённым — в непосредственной близости к людскому жилью — и оставались!. Кто знает, насколько пугливы дикие гуси, тому эти случаи дадут представление о силе их «стадного инстинкта». То же справедливо для очень многих общественных животных вплоть до шимпанзе, о которых Йеркс справедливо заметил: «Один шимпанзе — вообще не шимпанзе».
Даже те инстинкты, которые «только что» (с точки зрения филогенеза) приобрели самостоятельность через ритуализацию и, как я постарался показать в предыдущей главе, получили место и голос в Великом Парламенте Инстинктов в качестве самых молодых депутатов, — даже они при соответствующих обстоятельствах могут заглушить всех своих оппонентов точно так же, как Голод и Любовь. В триумфальном крике гусей мы увидим церемонию, которая управляет жизнью этих птиц больше, чем любой другой инстинкт. С другой стороны, разумеется, существует сколько угодно ритуализованных действий, которые ещё едва обособились от своего неритуализованного прототипа; их скромное влияние на общее поведение состоит лишь в том, что «желательная» для них координация движений — как мы видели в случае натравливания у огарей — становится в какой-то мере предпочтительной и используется чаще, чем другие, тоже возможные формы.
«Сильный» или «слабый» голос имеет ритуализованное действие в общем концерте инстинктов — оно во всех случаях чрезвычайно затрудняет любой мотивационный анализ, потому что может симулировать поведение, вытекающее из нескольких независимых побуждений. В предыдущей главе мы говорили, что ритуализованное действие, сплавленное в некоторую общность из различных компонентов, копирует форму последовательности движений, которая не является наследственно закреплённой и часто возникает из конфликта нескольких побуждений, как это видно на примере натравливания уток. А поскольку, как уже говорилось там же, копия и оригинал по большей части накладываются друг на друга в одном и том же движении, то чрезвычайно трудно разобраться, сколько же в нем от копии, а сколько от оригинала. Только когда один из первоначально независимых компонентов оказывается в противоречии с ритуально закреплённой координацией, — как направление на «врага», которому адресована угроза в случае натравливания, — тогда становится явным участие новых независимых переменных.
«Танец зигзага» у самцов колюшки, на котором Ян ван Йерсель провёл самый первый эксперимент мотивационного анализа, служит прекрасным примером того, как совсем «слабый» ритуал может вкрасться в конфликт двух «главных» инстинктов в качестве едва заметной третьей величины. Ван-Йерсель заметил, что замечательный танец зигзага, который половозрелые самцы, имеющие свой участок, исполняют перед каждой проплывающей мимо самкой, и который поэтому до тех пор считался просто «ухаживанием», — от случая к случаю выглядит совершенно по-разному. Оказалось, что иногда сильнее подчеркнут «зиг» в сторону самки, а иногда «заг» прочь от неё. Если это последнее движение очень явственно, то становится очевидным, что «заг» направлен в сторону гнёзда. В одном из предельных случаев самец при виде плывущей мимо самки быстро подплывает к ней, тормозит, разворачивается — особенно если самка тотчас поставит ему своё распухшее брюшко — и плывёт назад к входу в гнездо, которое затем показывает самке посредством определённой церемонии (ложась плоско на бок). В другом предельном случае, особенно частом если самка ещё не совсем готова к нересту, за первым «зигом» вообще не следует никакого «зага», а вместо того — нападение на самку.
Из этих наблюдений ван Йерсель правильно заключил, что «зиг» в сторону самки мотивируется агрессивным инстинктом, а «заг» в сторону гнёзда — сексуальным; и ему удалось экспериментально доказать правильность этого заключения. Он изобрёл методы, с помощью которых мог точно измерять силу агрессивного и сексуального инстинктов у каждого данного самца. Самцу предлагались макеты соперника стандартизованных размеров и регистрировалась интенсивность и продолжительность боевой реакции.
Сексуальный инстинкт измерялся с помощью макетов самки, которые внезапно убирались через определённое время.
В этих случаях самец «разряжает» внезапно заблокированный сексуальный инстинкт, совершая действия ухаживания за потомством, т.е. обмахивая плавниками как бы икру или мальков в гнезде; и продолжительность этого «заменяющего обмахивания» даёт надёжную меру сексуальной мотивации. Ван Йерсель научился предсказывать по результатам таких измерений, как будет выглядеть танец зигзага у данного самца, — и наоборот, по наблюдаемой форме танца заранее оценивать соотношения обоих инстинктов и результаты будущих измерений.
Но кроме обоих главных побуждений, определяющих движения самца колюшки в общих чертах, — на них оказывает влияние ещё какое-то третье, хоть и более слабое.
Это знаток ритуализованного поведения заподозрит сразу же, увидев ритмическую правильность смены «зигов» и «загов». Попеременное преобладание одного из двух противоречивых побуждений вряд ли может привести к столь регулярной смене направлений, если здесь не вступает в игру новая, ритуализованная координация. Без неё короткие рывки в разных направлениях следуют друг за другом с типичной случайностью, как это бывает у людей в состоянии крайней растерянности. Ритуализованное движение, напротив, всегда имеет тенденцию к ритмическому повторению в точности одинаковых элементов. Мы говорили об этом в связи с действенностью сигнала.
Подозрение, что здесь замешана ритуализация, превращается в уверенность, когда мы видим, как танцующий самец при своих «загах» временами, кажется, совершенно забывает, что они сексуально мотивированы и должны указывать точно на гнездо. Вместо этого он рисует вокруг самки очень красивый и правильный зубчатый венец, в котором каждый «зиг» направлен точно в сторону самки, а каждый «заг» — точно от неё. Как ни очевидна относительная слабость новой координации движений, стремящейся превратить «зиги» и «заги» в ритмический «зигзаг», — она может, однако, решающим образом определить регулярность последовательных проявлений обеих главных мотиваций.
Вторая важная функция, которую ритуализованная координация может, очевидно, выполнять, даже будучи очень слабой в других отношениях, — это изменение направления неритуализованных движений, лежавших в основе ритуала и происходивших из других побуждений. Примеры этого мы уже видели при обсуждении классического образца ритуала, а именно — при натравливании селезня уткой.
7. ПОВЕДЕНЧЕСКИЕ АНАЛОГИИ МОРАЛИ
Не убий.
Пятая заповедь
В 5-й главе, где речь шла о процессе ритуализации, я старался показать, как этот процесс, причины которого все ещё весьма загадочны, создаёт совершенно новые инстинкты, диктующие организму свои собственные «Ты должен…» так же категорично, как и любой из, казалось бы, единовластных «больших» инстинктов голода, страха или любви. В предыдущей 6-й главе я пытался решить ещё более трудную задачу: коротко и доступно показать, как происходит взаимодействие между различными автономными инстинктами, каким общим правилам подчиняются эти взаимодействия, и какими способами можно — несмотря на все сложности — получить некоторое представление о структуре взаимодействий в таком поведении, которое определяется несколькими соперничающими побуждениями.
Я тёшу себя надеждой, быть может обманчивой, что решить предыдущие задачи мне удалось, и что я могу не только обобщить сказанное в двух последних главах, но и применить полученные в них результаты к вопросу, которым мы займёмся теперь: каким образом ритуал выполняет поистине невыполнимую задачу — удерживает внутривидовую агрессию от всех проявлений, которые могли бы серьёзно повредить сохранению вида, но при этом не выключает её функций, необходимых для сохранения вида! Часть предыдущей фразы, выделенная курсивом, уже отвечает на вопрос, — он кажется очевидным, но вытекает из совершеннейшего непонимания сущности агрессии, — почему у тех животных, для которых тесная совместная жизнь является преимуществом, агрессия попросту не запрещена? Именно потому, что её функции, рассмотренные нами в 3-й главе, необходимы!
Решение проблем, возникающих таким образом перед обоими конструкторами эволюции, достигается всегда одним и тем же способом. Полезный, необходимый инстинкт — вообще остаётся неизменным; но для особых случаев, где его проявление было бы вредно, вводится специально созданный механизм торможения. И здесь снова культурно-историческое развитие народов происходит аналогичным образом; именно потому важнейшие требования Моисеевых и всех прочих скрижалей — это не предтисания, а запреты. Нам ещё придётся подробнее говоьрить о том, о чем здесь лишь предварительно упомянем:
передаваемые традицией и привычно выполняемые табу имеют какое-то отношение к разумной морали — в понимании Иммануила Канта — разве что у вдохновенного законодателя, но никак не у его верующих последователей.
Как врождённые механизмы и ритуалы, препятствующие асоциальному поведению животных, так и человеческие табу определяют поведение, аналогичное истинно моральному лишь с функциональной точки зрения; во всем остальном оно так же далеко от морали, как животное от человека! Но даже постигая сущность этих движущих мотивов, нельзя не восхищаться снова и снова при виде работы физиологических механизмов, которые побуждают животных к самоотверженному поведению, направленному на благо сообщества, как это предписывают нам, людям, законы морали.
Впечатляющий пример такого поведения, аналогичного человеческой морали, являют так называемые турнирные бои. Вся их организация направлена на то, чтобы выполнить важнейшую задачу поединка — определить, кто сильнее, — не причинив серьёзного вреда более слабому. Поскольку рыцарский турнир или спортивное состязание имеют ту же цель, то все турнирные бои неизбежно производят даже на знающих людей впечатление «рыцарственности», или «спортивного благородства». Среди цихлид есть вид, Cichlasoma biocellatum, который именно из-за этого приобрёл своё название, широко распространённое у американских любителей: у них эта рыбка называется «Джек Дэмпси» по имени боксёра, чемпиона мира, который прославился своим безупречным поведением на ринге.
О турнирных боях рыб и, в частности, о процессах ритуализации, которые привели к ним от первоначальных подлинных боев, мы знаем довольно много. Почти у всех костистых рыб настоящей схватке предшествуют угрожающие позы, которые, как уже говорилось, всегда вытекают из конфликта между стремлениями напасть и бежать.
Среди этих поз особенно заметна как специальный ритуал так называемая демонстрация развёрнутого бока, которая первоначально наверняка возникла за счёт того, что рыба под влиянием испуга отворачивается от противника и одновременно, готовясь к бегству, разворачивает вертикальные плавники. Но поскольку при этих движениях противнику предъявляется контур тела максимально возможных размеров, то из них — путём мимического утрирования при добавочных изменениях морфологии плавников — смогла развиться та впечатляющая демонстрация развёрнутого бока, которую знают все аквариумисты, да и не только они, по сиамским бойцовым рыбкам и по другим популярным породам рыб.
В тесной связи с угрозой развёрнутым боком у костистых рыб возник очень широко распространённый запугивающий жест — так называемый удар хвостом. Из позиции развёрнутого бока рыба, напрягая все тело и далеко оттопыривая хвостовой плавник, производит сильный удар хвостом в сторону противника. Хотя сам удар до противника не доходит, но рецепторы давления на его боковой линии воспринимают волну, сила которой, очевидно, сообщает ему о величине и боеспособности его соперника, так же как и размеры контура, видимого при демонстрации развёрнутого бока.
Другая форма угрозы возникла у многих окуневых и у других костистых рыб из заторможенного страхом фронтального удара. В исходной позиции для броска вперёд оба противника изгибают свои тела, словно напряжённые Sобразные пружины, и медленно плывут друг другу навстречу, как можно сильнее топорща жаберные крышки.
Это соответствует разворачиванию плавников при угрозе боком, поскольку увеличивает контур тела, видимый противником. Из фронтальной угрозы у очень многих рыб иногда получается, что оба противника одновременно хватают друг друга за пасть, но — в соответствии с конфликтной ситуацией, из которой возникла сама фронтальная угроза, — они всегда делают это не резко, не ударом, а словно колеблясь, заторможенно. Из этой формы борьбы у некоторых — и у лабиринтовых рыб, лишь отдалённо примыкающих к большой группе окуневых, и у цихлид, типичных представителей окуневых, — возникла интереснейшая ритуализованная борьба, при которой оба соперника в самом буквальном смысле слова «меряются силами», не причиняя друг другу вреда. Они хватают друг друга за челюсть — а у всех видов, для которых характерен этот способ турнирного боя, челюсть покрыта толстым, трудноуязвимым слоем кожи — и тянут изо всех сил. Так возникает состязание, очень похожее на старую борьбу на поясах у швейцарских крестьян, которое может продолжаться по нескольку часов, если встречаются равные противники. У двух в точности равных по силе самцов красивого синего вида широколобых окуньков мы запротоколировали однажды такой поединок, длившийся с 8.30 утра до 2.30 пополудни.