Обзаведясь фауной беспозвоночных, суша привлекла позвоночных хищников, которые прибыли в новую для себя экосреду в раннем верхнем девоне (век верхний фран, ок. 370 млн лет назад) [128а; 129, с. 420; 130, с. 508]; а также [246; 253; 274]. Вышедшие из пресноводных водоемов [834], первые земноводные были шестипалыми [345]. Они происходили от кистеперых рыб [334], а те – от рыб вроде Styloichthys changae (век поздний лочлов (жедин), нижний девон Китая), связывающих предков бесчелюстных рыбообразных с предками четвероногих позвоночных (земноводных, пресмыкающихся и млекопитающих) [835]. Рожденные межледниковьем, земноводные были маломобильными существами, а потому держались водоемов, привязанные к ним способом размножения (когда икра откладывается в воду, и там же развивается молодь, как у современных лягушек). Можно было бы предсказать, что, попади земноводные под давление отбора на высокомобильность, они изменили бы способ размножения на независимый от водной среды, принявшись прочесывать сушу и обзаведясь для этого непроницаемыми покровами тела взамен слизистой голой кожи, служащей для дыхания. Все это осуществили пресмыкающиеся.
По мере сближения материков климат девона портился, биопродуктивность среды падала, и маломобильные организмы погружались в кризис, что выразилось в позднедевонском вымирании (рубеж веков фран/ фамен, 367 млн лет назад), которое, помимо всего прочего, вызвало гибель тропических коралловых рифов и убыль чистой биомассы [184; 239]. Это прямо отражало снижение палеотемператур (чего не переносили прикрепленные маломобильные коралловые полипы) и оскудение среды.
Вслед за теплым девоном наступили каменноугольный (карбон, 354–286 млн лет назад) и пермский (286–251 млн лет назад) периоды. В это время континенты все более сближались, чтобы образовать сверхматерик Пангея (ок. 250–180 млн лет назад). Эти события вызвали пермо-карбоновое, или гондванское (по южной половине Пангеи – Гондване), оледенение (310–260 млн лет назад). Под давлением отбора на мобильность в соответствии с вышеописанным сценарием земноводные превратились в амниот (от греч. 'αμνίον – околозародышевый пузырь), т. е. начали откладывать яйца в плотной зародышевой оболочке, позволяющей им развиваться на суше. Защищенная чешуей кожа открывала недавним амфибиям сухопутные просторы. Появились пресмыкающиеся (век бригант (визей), нижний карбон, ок. 338 млн лет назад) [716].
На протяжении пермо-карбонового оледенения пресмыкающиеся сохраняли четвероногость, которая обеспечивала им максимальную мобильность. Ошибочно думать, будто ранние пресмыкающиеся не были в состоянии усвоить какую-либо другую локомоцию. Напротив, в их среде появилась интересная ящерицеобразная рептилия Eudibamus cursoris (из числа Bolosauridae, ранняя нижняя пермь, век ассель, 290 млн лет назад) [187]. Она напоминала ряд современных ящериц малопродуктивных, открытых пространств, бегающих рысью, переходящей в бег на двух задних ногах (такова, например, австралийская плащеносная ящерица), что отличается от энергосберегающей шагающей двуногой локомоции динозавро-человеческого типа. Отсюда следует, что E. сursoris был порожден оледенением и не имел ничего общего с двуногими динозаврами и людьми (в смыле конвергентной, внешне похожей эволюции), но подтверждал способность ранних рептилий разнообразить свою походку.
Здесь необходимо остановиться на способах локомоции (передвижения), присущих высшим наземным позвоночным. За вычетом промежуточных вариантов вроде бега E. cursoris или продвинутых кроличье-лягушачьих прыжков кенгуру, тушканчиков и им подобных животных существуют два основных типа локомоции: четвероногая, как у подавляющего большинства современных млекопитающих и «холодолюбивых» динозавров, и шагающая двуногая, как у «теплолюбивых» динозавров, людей и некоторых других существ.
Четвероногость весьма расточительна энергетически, сопряжена со значительным сцеплением животного с субстратом (из-за четырех конечностей), однако обеспечивает ему исключительную выносливость, проходимость, значительные скорости и дистанции передвижения. Подобная адаптация крайне полезна в прохладных условиях низкой биопродуктивности среды, когда скудную пищу приходится собирать с больших пространств.
В противоположность этому шагающая двуногость весьма выгодна, экономна энергетически, не сопряжена с большим сцеплением тела с субстратом (благодаря опоре на две конечности), однако не дает своему обладателю выносливости и не позволяет осваивать значительные территории. Такой способ передвижения выгоден в теплые эпохи высокой биопродуктивности среды, когда пища богата, скучена в пространстве и не требует больших путешествий для своей добычи. При этом двуногое существо очень выигрывает, экономя энергию на передвижении. С конкретными примерами такого рода мы познакомимся ниже.
Пермокарбоновое оледенение отмечено еще двумя перспективными достижениями биосферы. По молекулярно-генетическим данным, в это время возникли цветковые (покрытосеменные) растения. В конце нижнего карбона (век намюр, 319 ± 35 млн лет назад) они разделились на однодольные и двудольные, а в перми (276 ± 33 млн лет назад) двудольные растения претерпели радиацию [545]. Покрытосеменные растения были родственны растениям из рода Glossopteris, преобладавшему в перми и отличавшемуся простым способом размножения, как у цикадовых и гинкго [595]. Происходя из холодного ордовика, все наземные растения имели мобильные средства размножения (споры, семена) и совершенствовались в этой части. Возникнув в холодном карбоне, цветковые растения сделали здесь очередной шаг вперед: благодаря различным приспособлениям их семена распространялись на большие расстояния лучше, чем у голосеменных (например хвойных), хотя последние уступили преобладающее положение не сразу [88, с. 21–382].
Другое достижение касается холодолюбивой фауны, в рамках которой пресмыкающиеся котилозавры породили в верхнем карбоне звероподобных рептилий, господствовавших в перми, постепенно убывающих в триасе, вымирающих в средней юре, однако отмеченных в поздней юре и даже в позднем палеоцене (век ранний тиффан, ок. 60–57,3 млн лет назад) [25, с. 110–117; 87, с. 230–298; 250; 328; 655, с. 173–186]. От этих четвероногих рептилий в перми произошли зверозубые пресмыкающиеся – чрезвычайно прогрессивные существа: некоторые, по-видимому, стали теплокровными, покрылись шерстью и усвоили разнозубость. Зубы прочих пресмыкающихся обычно однородны (за вычетом динозавра гетеродонтозавра из позднего триаса или ранней юры (ок. 201 млн лет назад), у которого самцы имели клыки [37, с. 46–47]; гетеродонтность динозавра Eoraptor lunensis из верхнего триаса (ок. 230 млн лет назад) далека от разнообразия зверозубых рептилий и млекопитающих, cp. [689, с. 64, рис. 1]). У продвинутых же зверозубых пресмыкающихся зубы дифференцированы на резцы, клыки и коренные (как у их потомков – млекопитающих). Это требовалось для эффективного использования небогатой и разреженной пищи ледниковья. Казалось бы, перед прогрессивными зверозубыми рептилиями открыт весь мир. Между тем, их постепенно смело с эволюционной сцены.
С концом пермского похолодания грянуло позднепермское вымирание, неблагосклонное к мобильным организмам. Однако оно лишь потеснило цветковые растения и зверозубых рептилий. Это вымирание осуществлялось в две стадии: гваделупа, ок. 256 млн лет назад, истребившая 58% родов морских организмов, и татарий, ок. 251 млн лет назад, уничтоживший 67% родов морских организмов [723; 825 a]; cp. [35, с. 87, рис. 2; 184; 239; 311; 471; 639; 674, с. 437–439; 802].
Сухопутная биота понесла сходные потери, отвечающие эволюционно-экологической направленности естественного отбора, связанного с потеплением. Закономерно были утрачены небольшие подвижные рыбоеды и насекомоядные существа. Их участь разделили средние и крупные травоядные, привыкшие собирать скудные корма с обширных пространств. Высшие хищники потянулись следом. Даже спустя 15 млн лет этот урон среди пресмыкающихся еще не восполнился [185].
Легко понять, что избирательное двухступенчатое пермо-триасовое вымирание подчинялось определенной эволюционной логике, а не уничтожало живых существ без разбора, как свойственно стихийным бедствиям вроде падений астероидов или извержений вулканов. Как и в случае с мелтретичным вымиранием (см. далее), углубление исследований рисует эти биокатастрофы все менее окказионально (т. е. все менее случайно).
Мезозой (251-65 млн лет назад)
По мере сближения материков климат девона портился, биопродуктивность среды падала, и маломобильные организмы погружались в кризис, что выразилось в позднедевонском вымирании (рубеж веков фран/ фамен, 367 млн лет назад), которое, помимо всего прочего, вызвало гибель тропических коралловых рифов и убыль чистой биомассы [184; 239]. Это прямо отражало снижение палеотемператур (чего не переносили прикрепленные маломобильные коралловые полипы) и оскудение среды.
Вслед за теплым девоном наступили каменноугольный (карбон, 354–286 млн лет назад) и пермский (286–251 млн лет назад) периоды. В это время континенты все более сближались, чтобы образовать сверхматерик Пангея (ок. 250–180 млн лет назад). Эти события вызвали пермо-карбоновое, или гондванское (по южной половине Пангеи – Гондване), оледенение (310–260 млн лет назад). Под давлением отбора на мобильность в соответствии с вышеописанным сценарием земноводные превратились в амниот (от греч. 'αμνίον – околозародышевый пузырь), т. е. начали откладывать яйца в плотной зародышевой оболочке, позволяющей им развиваться на суше. Защищенная чешуей кожа открывала недавним амфибиям сухопутные просторы. Появились пресмыкающиеся (век бригант (визей), нижний карбон, ок. 338 млн лет назад) [716].
На протяжении пермо-карбонового оледенения пресмыкающиеся сохраняли четвероногость, которая обеспечивала им максимальную мобильность. Ошибочно думать, будто ранние пресмыкающиеся не были в состоянии усвоить какую-либо другую локомоцию. Напротив, в их среде появилась интересная ящерицеобразная рептилия Eudibamus cursoris (из числа Bolosauridae, ранняя нижняя пермь, век ассель, 290 млн лет назад) [187]. Она напоминала ряд современных ящериц малопродуктивных, открытых пространств, бегающих рысью, переходящей в бег на двух задних ногах (такова, например, австралийская плащеносная ящерица), что отличается от энергосберегающей шагающей двуногой локомоции динозавро-человеческого типа. Отсюда следует, что E. сursoris был порожден оледенением и не имел ничего общего с двуногими динозаврами и людьми (в смыле конвергентной, внешне похожей эволюции), но подтверждал способность ранних рептилий разнообразить свою походку.
Здесь необходимо остановиться на способах локомоции (передвижения), присущих высшим наземным позвоночным. За вычетом промежуточных вариантов вроде бега E. cursoris или продвинутых кроличье-лягушачьих прыжков кенгуру, тушканчиков и им подобных животных существуют два основных типа локомоции: четвероногая, как у подавляющего большинства современных млекопитающих и «холодолюбивых» динозавров, и шагающая двуногая, как у «теплолюбивых» динозавров, людей и некоторых других существ.
Четвероногость весьма расточительна энергетически, сопряжена со значительным сцеплением животного с субстратом (из-за четырех конечностей), однако обеспечивает ему исключительную выносливость, проходимость, значительные скорости и дистанции передвижения. Подобная адаптация крайне полезна в прохладных условиях низкой биопродуктивности среды, когда скудную пищу приходится собирать с больших пространств.
В противоположность этому шагающая двуногость весьма выгодна, экономна энергетически, не сопряжена с большим сцеплением тела с субстратом (благодаря опоре на две конечности), однако не дает своему обладателю выносливости и не позволяет осваивать значительные территории. Такой способ передвижения выгоден в теплые эпохи высокой биопродуктивности среды, когда пища богата, скучена в пространстве и не требует больших путешествий для своей добычи. При этом двуногое существо очень выигрывает, экономя энергию на передвижении. С конкретными примерами такого рода мы познакомимся ниже.
Пермокарбоновое оледенение отмечено еще двумя перспективными достижениями биосферы. По молекулярно-генетическим данным, в это время возникли цветковые (покрытосеменные) растения. В конце нижнего карбона (век намюр, 319 ± 35 млн лет назад) они разделились на однодольные и двудольные, а в перми (276 ± 33 млн лет назад) двудольные растения претерпели радиацию [545]. Покрытосеменные растения были родственны растениям из рода Glossopteris, преобладавшему в перми и отличавшемуся простым способом размножения, как у цикадовых и гинкго [595]. Происходя из холодного ордовика, все наземные растения имели мобильные средства размножения (споры, семена) и совершенствовались в этой части. Возникнув в холодном карбоне, цветковые растения сделали здесь очередной шаг вперед: благодаря различным приспособлениям их семена распространялись на большие расстояния лучше, чем у голосеменных (например хвойных), хотя последние уступили преобладающее положение не сразу [88, с. 21–382].
Другое достижение касается холодолюбивой фауны, в рамках которой пресмыкающиеся котилозавры породили в верхнем карбоне звероподобных рептилий, господствовавших в перми, постепенно убывающих в триасе, вымирающих в средней юре, однако отмеченных в поздней юре и даже в позднем палеоцене (век ранний тиффан, ок. 60–57,3 млн лет назад) [25, с. 110–117; 87, с. 230–298; 250; 328; 655, с. 173–186]. От этих четвероногих рептилий в перми произошли зверозубые пресмыкающиеся – чрезвычайно прогрессивные существа: некоторые, по-видимому, стали теплокровными, покрылись шерстью и усвоили разнозубость. Зубы прочих пресмыкающихся обычно однородны (за вычетом динозавра гетеродонтозавра из позднего триаса или ранней юры (ок. 201 млн лет назад), у которого самцы имели клыки [37, с. 46–47]; гетеродонтность динозавра Eoraptor lunensis из верхнего триаса (ок. 230 млн лет назад) далека от разнообразия зверозубых рептилий и млекопитающих, cp. [689, с. 64, рис. 1]). У продвинутых же зверозубых пресмыкающихся зубы дифференцированы на резцы, клыки и коренные (как у их потомков – млекопитающих). Это требовалось для эффективного использования небогатой и разреженной пищи ледниковья. Казалось бы, перед прогрессивными зверозубыми рептилиями открыт весь мир. Между тем, их постепенно смело с эволюционной сцены.
С концом пермского похолодания грянуло позднепермское вымирание, неблагосклонное к мобильным организмам. Однако оно лишь потеснило цветковые растения и зверозубых рептилий. Это вымирание осуществлялось в две стадии: гваделупа, ок. 256 млн лет назад, истребившая 58% родов морских организмов, и татарий, ок. 251 млн лет назад, уничтоживший 67% родов морских организмов [723; 825 a]; cp. [35, с. 87, рис. 2; 184; 239; 311; 471; 639; 674, с. 437–439; 802].
Сухопутная биота понесла сходные потери, отвечающие эволюционно-экологической направленности естественного отбора, связанного с потеплением. Закономерно были утрачены небольшие подвижные рыбоеды и насекомоядные существа. Их участь разделили средние и крупные травоядные, привыкшие собирать скудные корма с обширных пространств. Высшие хищники потянулись следом. Даже спустя 15 млн лет этот урон среди пресмыкающихся еще не восполнился [185].
Легко понять, что избирательное двухступенчатое пермо-триасовое вымирание подчинялось определенной эволюционной логике, а не уничтожало живых существ без разбора, как свойственно стихийным бедствиям вроде падений астероидов или извержений вулканов. Как и в случае с мелтретичным вымиранием (см. далее), углубление исследований рисует эти биокатастрофы все менее окказионально (т. е. все менее случайно).
Мезозой (251-65 млн лет назад)
Наступил триасовый период (251–201 млн лет назад), начало которого ознаменовалось «метановым потеплением» (вследствие парникового эффекта от вулканических газов, включая метан) [742] и морской трансгрессией (подъемом уровня Мирового океана) [74, с. 219, рис. 7.1; 401]. Естественный отбор принялся поощрять маломобильных животных. Еще в верхней перми от рептилий-эозухий, близких прогрессивным котилозаврам-капторинам и низшим зверообразным пеликозаврам (заложившим черты молекулярно-генетического родства своих потомков-млекопитающих и людей с потомками текодонтов, т. е. птицами), произошли пресмыкающиеся текодонты (псевдозухии). Под влиянием потепления и связанного с ним роста биопродуктивности среды, они усвоили экономичную шагающую двуногость человеческого типа, длительное время процветали, однако не перенесли похолодания (падения биопродуктивности среды) в конце триасового периода и вымерли. Однако до того, в середине триаса, морская трансгрессия и потепление обострили давление отбора в пользу маломобильных животных. В результате от текодонтов монофилетично (из одного корня) произошли двуногие динозавры двух отрядов (ящеротазовые и птицетазовые), 227,8 ± 0,3 млн лет назад (см. рис. 3) [87, с. 494, 525; 271; 324; 329; 653; 655, с. 138–141; 688–690].
Судя по своим поздним потомкам мелового периода и подобно ближайшим родичам, птицам и крокодилам, динозавры имели четырехкамерное сердце. Аналогично птицам они были покрыты перьями [148; 441; 747; 827–829], возможно, отличались постоянной температурой тела (гомеотермией) [173]; cp. [660] и даже спали в птичьей позе [826]. Они были сходны с птицами в особенностях роста челюсти [172], но расходились с ними в скоростях общего роста [310], что можно объяснить повышенным метаболизмом (обменом веществ) у последних вследствие летающего образа жизни. Кроме того, двуногим динозаврам, даже их верхнемеловому венцу, хищному колоссу тираннозавру, были присущи маломобильность [430] и равнозубость, ориентированная на расточительное потребление богатой и скученной пищи, – то и другое гармонировало с высокой биопродуктивностью среды.
Длительное триасовое потепление раздвинуло существующие экониши и вовлекло в них маломобильных существ, что привело пресмыкающихся в водную среду и дало известных ихтиозавров (средний триас – поздний мел), плезиозавров (тогда же) и других водных ящеров [25, с. 18, 22, 33]. Похолодание позднего триаса (норий-рэтийская морская регрессия, падение уровня Мирового океана) привело к упадку и частичному вымиранию маломобильных ихтиозавров. Выжили те их формы, которые лучше освоили живорождение, являющееся вариантом неявной мобильности.
То же похолодание отпочковало от динозавров подвижных летающих птиц, засвидетельствованных спорными остатками Protoavis texensis («первоптицы из Техаса», поздний триас, ок. 225 млн лет назад [141]; cp. [240; 326]), ископаемыми птичьими следами (поздний триас, века норий-рэт, св. 212,5 ± 7 млн лет назад [565]) и синхронным молекулярно-генетическим расхождением птиц с крокодилами (222 ± 52 млн лет назад [479]), что, впрочем, не исключает обособления предков птиц от крокодилов еще в составе динозавров (т. е. сначала от крокодилов отделились динозавры, а впоследствии, много позже, от динозавров обособились птицы вроде археоптерикса, которые положили начало современной авиафауне, что представляется нам сомнительным, см. ниже).
 Рис. 3. Ставрикозавр (Staurikosaurus pricei, «ящер созвездия Южного Креста») из семейства ставрикозаврид подотряда теропод (звероногов) отряда заурисхий (ящеротазовых динозавров) был ловким двуногим хищником и охотился в среднем триасе, не подозревая, что в конце этого периода его отсталые родичи прозауроподы встанут на четвереньки и дадут начало крупнейшим наземным животным планеты, зауроподам. [37, с. 30–31]
Рис. 3. Ставрикозавр (Staurikosaurus pricei, «ящер созвездия Южного Креста») из семейства ставрикозаврид подотряда теропод (звероногов) отряда заурисхий (ящеротазовых динозавров) был ловким двуногим хищником и охотился в среднем триасе, не подозревая, что в конце этого периода его отсталые родичи прозауроподы встанут на четвереньки и дадут начало крупнейшим наземным животным планеты, зауроподам. [37, с. 30–31]
Позднетриасовое вымирание (210 млн лет назад [184; 239; 518]) затронуло малоподвижных животных, но оживило четвероногих, что выразилось в оформлении млекопитающих как класса позвоночных животных (век норий, св. 200 млн лет назад [330]). Однако затем рэтийская регрессия ненадолго сменилась парниковым потеплением на 3–4° вследствие роста вулканического диоксида углерода 205,7 ± 4 млн лет назад [560], что спровоцировало вымирание мегафлоры, ориентированной на падение биопродуктивности среды.
Наступивший юрский период (201–144,2 млн лет назад) открылся очередной регрессией (похолоданием) и потряс двуногих динозавров – произошло их ограниченное нижнеюрское вымирание [239, с. 49]. Если целурозавры и карнозавры всегда оставались удачливыми, а потому эволюционно негибкими двуногами, то неуклюжие двуногие прозауроподы были готовы ко всему новому (о причинах чего см. далее) и охотно перешли к четвероногой локомоции, превратившись в гигантских зауропод (нижняя юра, век поздний лейас – верхний мел) (см. рис. 4, 5).
Отметим впечатляющую парадоксальность эволюционного процесса: предки динозавров (текодонты) сначала «встают» с четырех ног на две, порождают успешных двуногих динозавров, а затем вновь «опускаются» на четыре конечности (как произошло в случае с зауроподами и некоторыми другими динозаврами, см. далее). В рамках нашей версии эволюционной направленности этот нонсенс находит естественное объяснение – климато-экологическое. Возможно, обсуждаемое нижнеюрское похолодание развило у гетеродонтозавров разнозубость, являющуюся адаптацией к низкой биопродуктивности среды, свойственной похолоданиям.
После метанового потепления (ок.183 млн лет назад [742]) в середине средней юры (век поздний байос) палеотемпературы упали с наступлением антарктического микрооледенения, о чем свидетельствуют тиллиты (ископаемые морены, ледниковые отложения) юрского времени в Антарктиде [42, с. 33; 74, с. 220, 222], однако затем тепло стало нарастать к максимуму трансгрессии в середине верхней юры (конец века кимериджа). Средне-верхнеюрское потепление пагубно сказалось на четвероногих звероподобных рептилиях, которые почти полностью вымерли, оставив по себе молодых отпрысков, млекопитающих. То же самое произошло с четвероногими зауроподами, испытавшими с потеплением первый эпизод вымирания [25, с. 56]. Их судьбу повторили четвероногие птицетазовые динозавры сцелидозавры из ранней юры, когда их четвероногость была закономерна. Во второй половине юрского периода они дали четвероногих же стегозавров (середина юры – ранний мел), а через сирмозавров (конец раннего мела) – панцирных анкилозавров (ранний – поздний мел) [25, с. 79]. Иными словами, эволюция этих существ шла как бы «ракетным способом», когда в моменты потеплений отбрасываются «отработанные» (устаревшие и вымирающие) ступени.
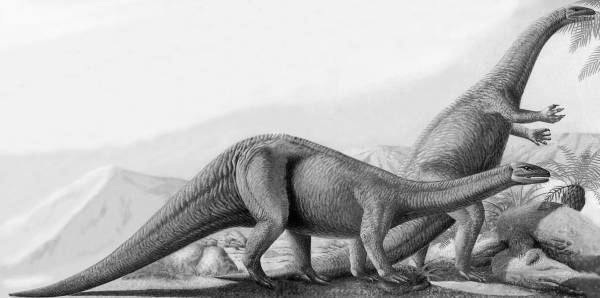 Рис. 4. Платеозавр (Plateosaurus engelhardti, плоский ящер Энгельгардта) из семейства платеозаврид инфраотряда прозауропод (проящероногов) из подотряда теропод группы зауроподоморфов (ящероногоподобных) отряда заурисхий. Это неуклюжее двуногое животное питалось растительностью, но происходило от совершенных двуногих динозавров среднего триасового периода, хищных теропод. В позднем триасе под влиянием похолодания и падения биопродуктивности среды прозауроподы стали возвращаться к четвероногому способу передвижения, свойственному далеким «холодолюбивым» предкам двуногих текодонтов и динозавров [37, с. 34–35]
Рис. 4. Платеозавр (Plateosaurus engelhardti, плоский ящер Энгельгардта) из семейства платеозаврид инфраотряда прозауропод (проящероногов) из подотряда теропод группы зауроподоморфов (ящероногоподобных) отряда заурисхий. Это неуклюжее двуногое животное питалось растительностью, но происходило от совершенных двуногих динозавров среднего триасового периода, хищных теропод. В позднем триасе под влиянием похолодания и падения биопродуктивности среды прозауроподы стали возвращаться к четвероногому способу передвижения, свойственному далеким «холодолюбивым» предкам двуногих текодонтов и динозавров [37, с. 34–35]
Нижнеюрское похолодание с его давлением в пользу мобильности выщепило из среды архозавров (родственников динозавров) летающих ящеров птерозавров, покрытых шерстью. Они просуществовали до верхнего мела и разделили судьбу динозавровой биоты [25, с. 106–109]. Их вымирание объясняется тем, что, в отличие от сноровистых, высокоподвижных птиц современного типа, они были планирующими, энергосберегающими созданиями, а не активными летунами, т. е. характеризовались ограниченной мобильностью [25, с. 106; 87, с. 593], в то время как маастрихтское (конца мела) и палеоценовое (после мела) похолодания требовали от животных более сильно выраженной подвижности.
 Рис. 5. Барапазавр (Barapasaurus tagorsi, большеногий ящер) из подотряда зауропод (ящероногов) группы зауроподоморфов отряда заурисхий. Под влиянием ухудшения климата и падения биопродуктивности среды неуклюжие двуногие прозауроподы породили в раннем юрском периоде совершенных четвероногих зауропод. Они процветали и достигли размеров, рекордных для наземных животных [37, с. 42–43]
Рис. 5. Барапазавр (Barapasaurus tagorsi, большеногий ящер) из подотряда зауропод (ящероногов) группы зауроподоморфов отряда заурисхий. Под влиянием ухудшения климата и падения биопродуктивности среды неуклюжие двуногие прозауроподы породили в раннем юрском периоде совершенных четвероногих зауропод. Они процветали и достигли размеров, рекордных для наземных животных [37, с. 42–43]
В поздней юре (150,7–144,2 млн лет назад) максимум трансгрессии сменился регрессией и похолоданием, что возобновило селекцию на подвижность и побудило динозавров вторично породить птицеподобных существ – в частности, известного археоптерикса [240, с. 349], который строением зрительной и слуховой коры головного мозга действительно напоминал современных птиц [136], что явилось результатом конвергенции, параллельной эволюции. Будучи предком энантиорнитинов, или «противоположных птиц» (от последовательности совмещения костей стопы в ходе развития, противоположной современным птицам), археоптерикс напоминал миниатюрного велоцираптора или дромеозавра [697], тоже пернатых динозавров, и отличался от протоависа, предка орнитуринов, давших современных птиц. Динозавров, археоптерикса и энантиорнитинов объединяли примитивные признаки эктотермности, т. е. непостоянной температуры тела [660].
Меловой период (144,2–65 млн лет назад) ознаменовался нижнемеловой регрессией (век валанжин, 130 млн лет назад) и продолжением отбора на мобильность. В это время развивались упомянутые тупиковые энантиорнитины и перспективные орнитурины, в среде которых началась дивергенция современных птиц (средний нижний мел, век апт, 122,9 ± 0,3, 121,2 ± 0,3–121,1 ± 0,2 млн лет назад) [260; 747]. За тот же временной интервал пернатые динозавры породили еще одно птицеобразное создание – четырехкрылого Microraptor gui из Китая [829], все четыре конечности которого годились для планирования, повышая подвижность. Среди современников этого маленького дромеозавра – китайских птиц-энантиорнитинов – также встречался четырехкрылый вид [833], так что данная эволюционная тенденция проявлялась неоднократно.
Если зауроподы откликнулись на падение биопродуктивности среды мобильной четвероногостью, то орнитоподы, ящеротазовые четвероноги (игуанодоны) и двуноги (утконосые динозавры-гадрозавры, верхняя юра – верхний мел) отреагировали иначе: они освоили умение жевать, как млекопитающие [596], что повышало эффективность потребления редкой пищи соответственно похолоданию. В этом отношении названные представители динозаврового племени принципиально отличались от прочих архозавров, т. е. других динозавров, а также крокодилов. Последние, кстати, пережили роковой меловой период в силу того обстоятельства, что являлись ловкими наземными бегунами-четвероногами, т. е. мобильными животными, что открывало им путь в холодный кайнозой, где они, впрочем, вскоре «замерзли» и перешли к водному образу жизни в тропических и субтропических реках, не выдержав конкуренции с крепнущими наземными четвероногими млекопитающими.
Рубеж нижнего-верхнего мела (105 млн лет назад) прошел с трансгрессией, а века турон – коньяк (92–86 млн лет назад) подверглись вулканическому диоксид-углеродному потеплению [750]. В конце мела, в маастрихте (71–66 млн лет назад) регрессия сопровождалась заметным похолоданием [526], когда тропические температуры в океанах упали ниже современных [288]. К этому времени изнеженные теплым и обильным мезозоем (251–65 млн лет назад) динозавры находились на вершине своего расцвета, о чем свидетельствует сам факт существования крупнейшего наземного хищника – королевского тираннозавра (76–65 млн лет назад) (см. рис. 6). Сопряженная с ним мегафауна стала совершенной и высокоспециализированной, а потому могла лишь углубляться в аппетитах, но никак не ограничивать себя, чего требовала оскудевающая обстановка.
Вымирание динозавров происходило постепенно и пропорционально прогрессу млекопитающих (по модели цветковых растений и костистых рыб, см. далее). В веке кампан (76–73 млн лет назад) на пик расцвета динозавров приходилось 30 их родов. С приближением маастрихтского похолодания (71–66 млн лет назад) гордые животные ощутили эволюционно-экологическое затруднение: они любили много потреблять и мало двигаться, а теперь среда благоволила к сноровистым аскетам типа четвероногих млекопитающих.
 Рис. 6. Тираннозавр (Tyrannosaurus rex, королевский ящер-тиран) из семейства тираннозаврид инфраотряда карнозавров (хищных ящеров) подотряда теропод отряда заурисхий. Это был крупнейший наземный хищник всех времен, чуть уступавший лишь хищному гигантозавру начала верхнего мела (ок. 100 млн лет назад), который, будучи южноамериканцем, закономерно превосходил массой североамериканского собрата, как свойственно рептилиям с приближением к экватору (у млекопитающих – наоборот). Тираннозавр был совершенным хищником (и падальщиком, как современные львы), узко приспособленным к среде, богатой пищей. С ухудшением климата и падением биопродуктивности среды королевский ящер угас и вымер вместе с подавляющим большинством остальных динозавров. По современным данным, он был оперенным существом [37, с. 98–99]
Рис. 6. Тираннозавр (Tyrannosaurus rex, королевский ящер-тиран) из семейства тираннозаврид инфраотряда карнозавров (хищных ящеров) подотряда теропод отряда заурисхий. Это был крупнейший наземный хищник всех времен, чуть уступавший лишь хищному гигантозавру начала верхнего мела (ок. 100 млн лет назад), который, будучи южноамериканцем, закономерно превосходил массой североамериканского собрата, как свойственно рептилиям с приближением к экватору (у млекопитающих – наоборот). Тираннозавр был совершенным хищником (и падальщиком, как современные львы), узко приспособленным к среде, богатой пищей. С ухудшением климата и падением биопродуктивности среды королевский ящер угас и вымер вместе с подавляющим большинством остальных динозавров. По современным данным, он был оперенным существом [37, с. 98–99]
За весь маастрихт и две трети дания (72–65,3 млн лет назад) число родов динозавров упало до 12. Затем вымирание ускорилось, и за период 65,3–64, 96 млн лет назад прошло пять убывающих стадий: 12–12–11–10–7 родов уходящих динозавров. О том, что биосфера отреагировала на коварный удар иридиевого астероида 65 млн лет назад, не может быть и речи. Параллельно со ступенчатым падением динозавров шел уверенный прогресс копытных млекопитающих, представленных в тех же временных точках 0–1 – 3–5 – 8 родами. Так что за 40 тыс. лет, истекших с момента падения астероида, млекопитающие зримо окрепли, а динозавры еще не вымерли окончательно, являя редкий случай послемеловых, палеоценовых динозавров, останки которых обнаружены выше мелтретичной иридиевой аномалии, оставленной упавшим болидом [714; 713]; cp. [222; 642; 696].
Судя по своим поздним потомкам мелового периода и подобно ближайшим родичам, птицам и крокодилам, динозавры имели четырехкамерное сердце. Аналогично птицам они были покрыты перьями [148; 441; 747; 827–829], возможно, отличались постоянной температурой тела (гомеотермией) [173]; cp. [660] и даже спали в птичьей позе [826]. Они были сходны с птицами в особенностях роста челюсти [172], но расходились с ними в скоростях общего роста [310], что можно объяснить повышенным метаболизмом (обменом веществ) у последних вследствие летающего образа жизни. Кроме того, двуногим динозаврам, даже их верхнемеловому венцу, хищному колоссу тираннозавру, были присущи маломобильность [430] и равнозубость, ориентированная на расточительное потребление богатой и скученной пищи, – то и другое гармонировало с высокой биопродуктивностью среды.
Длительное триасовое потепление раздвинуло существующие экониши и вовлекло в них маломобильных существ, что привело пресмыкающихся в водную среду и дало известных ихтиозавров (средний триас – поздний мел), плезиозавров (тогда же) и других водных ящеров [25, с. 18, 22, 33]. Похолодание позднего триаса (норий-рэтийская морская регрессия, падение уровня Мирового океана) привело к упадку и частичному вымиранию маломобильных ихтиозавров. Выжили те их формы, которые лучше освоили живорождение, являющееся вариантом неявной мобильности.
То же похолодание отпочковало от динозавров подвижных летающих птиц, засвидетельствованных спорными остатками Protoavis texensis («первоптицы из Техаса», поздний триас, ок. 225 млн лет назад [141]; cp. [240; 326]), ископаемыми птичьими следами (поздний триас, века норий-рэт, св. 212,5 ± 7 млн лет назад [565]) и синхронным молекулярно-генетическим расхождением птиц с крокодилами (222 ± 52 млн лет назад [479]), что, впрочем, не исключает обособления предков птиц от крокодилов еще в составе динозавров (т. е. сначала от крокодилов отделились динозавры, а впоследствии, много позже, от динозавров обособились птицы вроде археоптерикса, которые положили начало современной авиафауне, что представляется нам сомнительным, см. ниже).
Позднетриасовое вымирание (210 млн лет назад [184; 239; 518]) затронуло малоподвижных животных, но оживило четвероногих, что выразилось в оформлении млекопитающих как класса позвоночных животных (век норий, св. 200 млн лет назад [330]). Однако затем рэтийская регрессия ненадолго сменилась парниковым потеплением на 3–4° вследствие роста вулканического диоксида углерода 205,7 ± 4 млн лет назад [560], что спровоцировало вымирание мегафлоры, ориентированной на падение биопродуктивности среды.
Наступивший юрский период (201–144,2 млн лет назад) открылся очередной регрессией (похолоданием) и потряс двуногих динозавров – произошло их ограниченное нижнеюрское вымирание [239, с. 49]. Если целурозавры и карнозавры всегда оставались удачливыми, а потому эволюционно негибкими двуногами, то неуклюжие двуногие прозауроподы были готовы ко всему новому (о причинах чего см. далее) и охотно перешли к четвероногой локомоции, превратившись в гигантских зауропод (нижняя юра, век поздний лейас – верхний мел) (см. рис. 4, 5).
Отметим впечатляющую парадоксальность эволюционного процесса: предки динозавров (текодонты) сначала «встают» с четырех ног на две, порождают успешных двуногих динозавров, а затем вновь «опускаются» на четыре конечности (как произошло в случае с зауроподами и некоторыми другими динозаврами, см. далее). В рамках нашей версии эволюционной направленности этот нонсенс находит естественное объяснение – климато-экологическое. Возможно, обсуждаемое нижнеюрское похолодание развило у гетеродонтозавров разнозубость, являющуюся адаптацией к низкой биопродуктивности среды, свойственной похолоданиям.
После метанового потепления (ок.183 млн лет назад [742]) в середине средней юры (век поздний байос) палеотемпературы упали с наступлением антарктического микрооледенения, о чем свидетельствуют тиллиты (ископаемые морены, ледниковые отложения) юрского времени в Антарктиде [42, с. 33; 74, с. 220, 222], однако затем тепло стало нарастать к максимуму трансгрессии в середине верхней юры (конец века кимериджа). Средне-верхнеюрское потепление пагубно сказалось на четвероногих звероподобных рептилиях, которые почти полностью вымерли, оставив по себе молодых отпрысков, млекопитающих. То же самое произошло с четвероногими зауроподами, испытавшими с потеплением первый эпизод вымирания [25, с. 56]. Их судьбу повторили четвероногие птицетазовые динозавры сцелидозавры из ранней юры, когда их четвероногость была закономерна. Во второй половине юрского периода они дали четвероногих же стегозавров (середина юры – ранний мел), а через сирмозавров (конец раннего мела) – панцирных анкилозавров (ранний – поздний мел) [25, с. 79]. Иными словами, эволюция этих существ шла как бы «ракетным способом», когда в моменты потеплений отбрасываются «отработанные» (устаревшие и вымирающие) ступени.
Нижнеюрское похолодание с его давлением в пользу мобильности выщепило из среды архозавров (родственников динозавров) летающих ящеров птерозавров, покрытых шерстью. Они просуществовали до верхнего мела и разделили судьбу динозавровой биоты [25, с. 106–109]. Их вымирание объясняется тем, что, в отличие от сноровистых, высокоподвижных птиц современного типа, они были планирующими, энергосберегающими созданиями, а не активными летунами, т. е. характеризовались ограниченной мобильностью [25, с. 106; 87, с. 593], в то время как маастрихтское (конца мела) и палеоценовое (после мела) похолодания требовали от животных более сильно выраженной подвижности.
В поздней юре (150,7–144,2 млн лет назад) максимум трансгрессии сменился регрессией и похолоданием, что возобновило селекцию на подвижность и побудило динозавров вторично породить птицеподобных существ – в частности, известного археоптерикса [240, с. 349], который строением зрительной и слуховой коры головного мозга действительно напоминал современных птиц [136], что явилось результатом конвергенции, параллельной эволюции. Будучи предком энантиорнитинов, или «противоположных птиц» (от последовательности совмещения костей стопы в ходе развития, противоположной современным птицам), археоптерикс напоминал миниатюрного велоцираптора или дромеозавра [697], тоже пернатых динозавров, и отличался от протоависа, предка орнитуринов, давших современных птиц. Динозавров, археоптерикса и энантиорнитинов объединяли примитивные признаки эктотермности, т. е. непостоянной температуры тела [660].
Меловой период (144,2–65 млн лет назад) ознаменовался нижнемеловой регрессией (век валанжин, 130 млн лет назад) и продолжением отбора на мобильность. В это время развивались упомянутые тупиковые энантиорнитины и перспективные орнитурины, в среде которых началась дивергенция современных птиц (средний нижний мел, век апт, 122,9 ± 0,3, 121,2 ± 0,3–121,1 ± 0,2 млн лет назад) [260; 747]. За тот же временной интервал пернатые динозавры породили еще одно птицеобразное создание – четырехкрылого Microraptor gui из Китая [829], все четыре конечности которого годились для планирования, повышая подвижность. Среди современников этого маленького дромеозавра – китайских птиц-энантиорнитинов – также встречался четырехкрылый вид [833], так что данная эволюционная тенденция проявлялась неоднократно.
Если зауроподы откликнулись на падение биопродуктивности среды мобильной четвероногостью, то орнитоподы, ящеротазовые четвероноги (игуанодоны) и двуноги (утконосые динозавры-гадрозавры, верхняя юра – верхний мел) отреагировали иначе: они освоили умение жевать, как млекопитающие [596], что повышало эффективность потребления редкой пищи соответственно похолоданию. В этом отношении названные представители динозаврового племени принципиально отличались от прочих архозавров, т. е. других динозавров, а также крокодилов. Последние, кстати, пережили роковой меловой период в силу того обстоятельства, что являлись ловкими наземными бегунами-четвероногами, т. е. мобильными животными, что открывало им путь в холодный кайнозой, где они, впрочем, вскоре «замерзли» и перешли к водному образу жизни в тропических и субтропических реках, не выдержав конкуренции с крепнущими наземными четвероногими млекопитающими.
Рубеж нижнего-верхнего мела (105 млн лет назад) прошел с трансгрессией, а века турон – коньяк (92–86 млн лет назад) подверглись вулканическому диоксид-углеродному потеплению [750]. В конце мела, в маастрихте (71–66 млн лет назад) регрессия сопровождалась заметным похолоданием [526], когда тропические температуры в океанах упали ниже современных [288]. К этому времени изнеженные теплым и обильным мезозоем (251–65 млн лет назад) динозавры находились на вершине своего расцвета, о чем свидетельствует сам факт существования крупнейшего наземного хищника – королевского тираннозавра (76–65 млн лет назад) (см. рис. 6). Сопряженная с ним мегафауна стала совершенной и высокоспециализированной, а потому могла лишь углубляться в аппетитах, но никак не ограничивать себя, чего требовала оскудевающая обстановка.
Вымирание динозавров происходило постепенно и пропорционально прогрессу млекопитающих (по модели цветковых растений и костистых рыб, см. далее). В веке кампан (76–73 млн лет назад) на пик расцвета динозавров приходилось 30 их родов. С приближением маастрихтского похолодания (71–66 млн лет назад) гордые животные ощутили эволюционно-экологическое затруднение: они любили много потреблять и мало двигаться, а теперь среда благоволила к сноровистым аскетам типа четвероногих млекопитающих.
За весь маастрихт и две трети дания (72–65,3 млн лет назад) число родов динозавров упало до 12. Затем вымирание ускорилось, и за период 65,3–64, 96 млн лет назад прошло пять убывающих стадий: 12–12–11–10–7 родов уходящих динозавров. О том, что биосфера отреагировала на коварный удар иридиевого астероида 65 млн лет назад, не может быть и речи. Параллельно со ступенчатым падением динозавров шел уверенный прогресс копытных млекопитающих, представленных в тех же временных точках 0–1 – 3–5 – 8 родами. Так что за 40 тыс. лет, истекших с момента падения астероида, млекопитающие зримо окрепли, а динозавры еще не вымерли окончательно, являя редкий случай послемеловых, палеоценовых динозавров, останки которых обнаружены выше мелтретичной иридиевой аномалии, оставленной упавшим болидом [714; 713]; cp. [222; 642; 696].
Конец бесплатного ознакомительного фрагмента