Страница:
s-B-D-E-s
ws
wT-. Разведенцы далматинов вывели собак с более крупными, яркими, прокрашенными пятнами, случайно разбросанными по всему телу.
Экспрессия гена тиковой пятнистости Т варьирует от слегка крапчатые собак с несколькими крапивами только на конечностях и животе до сильно крапчатых животных, часто встречающихся у гончих пород (например, гасконская гончая и гриффон). В последнем случае собаку с таким окрасом можно назвать чалой. С другой стороны, варьирует не только количество крапа, но и качество его. Так что тиковость является весьма вариабельным признаком. Является ли это результатом отбора и закрепления в генотипе определенного количества полигенов, управляющих экспрессией гена Т, еще предстоит выяснить. Литтл заметил, что при скрещивании далматинов и собак с окрасом, похожим на далматинский, в первом поколении происходит возврат к обычной крапчатости. Возможно, что существует не один ген тиковости, каждый со своим характерным типом крапчатости. Однако, тот факт, что у собак наблюдается экстраординарная вариабельность экспрессии тиковости сам по себе не может служить этому доказательством.
Маски
Руфизм (рыжина)
Вариабельность чепрачности
Полигены затенения или соболиного окраса
Белые собаки
Окрас радужины глаза
Цвет мочки носа
КАЧЕСТВО ШЕРСТИ
СРАВНИТЕЛЬНАЯ СИМВОЛИКА ГЕНОВ
Заключение
Экспрессия гена тиковой пятнистости Т варьирует от слегка крапчатые собак с несколькими крапивами только на конечностях и животе до сильно крапчатых животных, часто встречающихся у гончих пород (например, гасконская гончая и гриффон). В последнем случае собаку с таким окрасом можно назвать чалой. С другой стороны, варьирует не только количество крапа, но и качество его. Так что тиковость является весьма вариабельным признаком. Является ли это результатом отбора и закрепления в генотипе определенного количества полигенов, управляющих экспрессией гена Т, еще предстоит выяснить. Литтл заметил, что при скрещивании далматинов и собак с окрасом, похожим на далматинский, в первом поколении происходит возврат к обычной крапчатости. Возможно, что существует не один ген тиковости, каждый со своим характерным типом крапчатости. Однако, тот факт, что у собак наблюдается экстраординарная вариабельность экспрессии тиковости сам по себе не может служить этому доказательством.
Маски
Маска — термин, употребляющийся для обозначения черного поля на морде, часто покрывающего переднюю часть «лица» и ушей. Наиболее четко маска видна у А
у-рыжих собак, например, у таких разных пород, как желто-коричневая (оленья) вариация гигантского мастифа и у миниатюрного мопса.
У чепрачных и черно-подпалых собак маска, если она есть, затемняет подпалы, ассоциированные с генами a saи a t. В этом случае получаются черномордые чепрачные и черно-подпалые собаки. У первых, ген маски трудно выявить, т.к.
наличие или отсутствие темной морды можно также рассматривать и как вариации чепрачности. Черно-подпалый окрас более стабилен и однообразен, поэтому маска у черноподпалых собак сразу же обнаруживается, благодаря отсутствию подпалов на морде. Естественно, что маску невозможно обнаружить у черных A s-собак, хотя в геноме соответствующий ген может присутствовать.
Маска может варьировать по степени развития. Так, она может покрывать всю «лицевую» часть головы и уши. Но это не обязательно означает влияние полигенов. О характере наследуемости маски известно очень мало. Литтл (1957) ссылается на случай рождения от рыжей таксы и мопса оленьего окраса с маской всех восьми щенков с черными масками. Это указывает на моногенность и доминантность маски как признака. Условно, ген маски можно обозначить как Ма.
Литтл считает, что существует несколько аллельных генов маски, но не дает четких доказательств. Тигровые животные могут быть как о маской так и без нее, что само по себе достаточно, чтобы отбросить всякие сомнения о моногенности признака. Геи маски, возможно, не экспрессируется у ee-животных, но и это не свидетельствует о наличии серии аллелей. Возможно, что ee-рыжие животные не могут вообще синтезировать черный пигмент в волосе. Отсутствие маски у ee-рыжих собак можно рассматривать как признак, отличающий их от А у-рыжих животных. Все рыжие собаки с маской имеют ген А у, тогда как рыжие без маски могут иметь как Ay так и ee, но, скорее всего последний.
У чепрачных и черно-подпалых собак маска, если она есть, затемняет подпалы, ассоциированные с генами a saи a t. В этом случае получаются черномордые чепрачные и черно-подпалые собаки. У первых, ген маски трудно выявить, т.к.
наличие или отсутствие темной морды можно также рассматривать и как вариации чепрачности. Черно-подпалый окрас более стабилен и однообразен, поэтому маска у черноподпалых собак сразу же обнаруживается, благодаря отсутствию подпалов на морде. Естественно, что маску невозможно обнаружить у черных A s-собак, хотя в геноме соответствующий ген может присутствовать.
Маска может варьировать по степени развития. Так, она может покрывать всю «лицевую» часть головы и уши. Но это не обязательно означает влияние полигенов. О характере наследуемости маски известно очень мало. Литтл (1957) ссылается на случай рождения от рыжей таксы и мопса оленьего окраса с маской всех восьми щенков с черными масками. Это указывает на моногенность и доминантность маски как признака. Условно, ген маски можно обозначить как Ма.
Литтл считает, что существует несколько аллельных генов маски, но не дает четких доказательств. Тигровые животные могут быть как о маской так и без нее, что само по себе достаточно, чтобы отбросить всякие сомнения о моногенности признака. Геи маски, возможно, не экспрессируется у ee-животных, но и это не свидетельствует о наличии серии аллелей. Возможно, что ee-рыжие животные не могут вообще синтезировать черный пигмент в волосе. Отсутствие маски у ee-рыжих собак можно рассматривать как признак, отличающий их от А у-рыжих животных. Все рыжие собаки с маской имеют ген А у, тогда как рыжие без маски могут иметь как Ay так и ee, но, скорее всего последний.
Руфизм (рыжина)
Интенсивность желтого пигмента шерсти может варьировать от бледно-кремового до насыщенного рыжего. Некоторые из вариаций можно рассматривать как результат действия основных генов окраса, таких как d или, возможно, c
ch(шиншилла), но не все. Остается еще значительное количество вариаций, которые обязаны действию группы полигенов, известных как руфус-полигены. Причем, эти полигены воздействуют как на A
yтак и на ее. Интенсивность пигментации варьирует в зависимости от особи, линии или породы. Эти же модификаторы также определяют интенсивность пигментации подпалов у чепрачных и черно-подпалых собак, у которых они могут варьировать от бледно-желтых до насыщенно красных. Это поднимает статус данной группы генов о простых модификаторов генов А
уи е до фундаментальной группы генов, определяющих основной составной элемент окраса. Только сплошной черный или сплошной белый окрасы не подчинены влиянию руфус-полигенов.
Действие этих генов в основном направлено на усиление пигментации. Волк имеет тусклый, однообразный окрас и собаководы прошлого, очевидно, отбирали более яркоокрашенных особей, определяя таким образом внешний вид большинства пород. Ильин (1932, 1941) обнаружил, что более бледный окрас доминантен по отношению к более яркому и объяснил это существованием серии аллелей «интенсивности» (символы Int, int mи int). Аллель Int продуцирует наиболее бледные тона, a int — наиболее яркие, int mзанимает промежуточное положение. Вполне возможно, что интенсивность желтой пигментации действительно регулируется мажорными генами (а не полигенами), но, чтобы доказать это, у Ильина было слишком мало наблюдений. Скорее всего, в этот процесс вовлечены многочисленные руфус-полигены. Наблюдения Ильина интересны и, возможно, полезны, т.к. потерю яркого рыжего окраса у гибридов между яркими и осветленными собаками наблюдали Njebbes и Wriedt (1927). Бернс и Фразер (1966), а также Виллис (1976) заметили, что насыщенность рыжего окраса может быть легко потеряна и трудно затем восстанавливается. Это означает, что, если есть усиливающие и ослабляющие полигены, то последние должны быть доминантны по отношению к первым. Однако, для этой группы полигенов характерно, что те, которые доминантны в одних скрещиваниях, становятся рецессивны в других, в зависимости от родителей. Т.о. очень трудно выявить какие-либо закономерности, лежащие в основе взаимодействия этих генов.
Действие этих генов в основном направлено на усиление пигментации. Волк имеет тусклый, однообразный окрас и собаководы прошлого, очевидно, отбирали более яркоокрашенных особей, определяя таким образом внешний вид большинства пород. Ильин (1932, 1941) обнаружил, что более бледный окрас доминантен по отношению к более яркому и объяснил это существованием серии аллелей «интенсивности» (символы Int, int mи int). Аллель Int продуцирует наиболее бледные тона, a int — наиболее яркие, int mзанимает промежуточное положение. Вполне возможно, что интенсивность желтой пигментации действительно регулируется мажорными генами (а не полигенами), но, чтобы доказать это, у Ильина было слишком мало наблюдений. Скорее всего, в этот процесс вовлечены многочисленные руфус-полигены. Наблюдения Ильина интересны и, возможно, полезны, т.к. потерю яркого рыжего окраса у гибридов между яркими и осветленными собаками наблюдали Njebbes и Wriedt (1927). Бернс и Фразер (1966), а также Виллис (1976) заметили, что насыщенность рыжего окраса может быть легко потеряна и трудно затем восстанавливается. Это означает, что, если есть усиливающие и ослабляющие полигены, то последние должны быть доминантны по отношению к первым. Однако, для этой группы полигенов характерно, что те, которые доминантны в одних скрещиваниях, становятся рецессивны в других, в зависимости от родителей. Т.о. очень трудно выявить какие-либо закономерности, лежащие в основе взаимодействия этих генов.
Вариабельность чепрачности
Экспрессия гена чепрачности настолько вариабельна, что можно предположить наличие более, чем одного аллеля, определяющего этот признак. Например, один аллель дает темно-чепрачный окрас, другой — светло-чепрачный. Вариации чепрачности составляют непрерывный ряд, что можно объяснить скорее влиянием модифицирующих полигенов. Чепрачный окрас часто можно спутать с соболиным, т.к. он имеет тенденцию с возрастом осветляться. Это говорит о генетической близости аллелей a
saи A
y. С другой стороны, когда чепрак распространяется на большую часть тела, окрас становится похожим на черно-подпалый.
В идеале, чепрачными принято называть собак с V-образным темным пятном на спине и боках туловища, простирающимся от хребта до живота. Однако, как распространение, так и интенсивность пигментации этой области может варьировать. При яркой, интенсивной пигментации чепрачный окрас легко идентифицировать, но при более светлой пигментации границы чепрака размыты и наличие его трудно распознать. Фактически, этот окрас стремится иметь сходство с диким волчеобразным. И это является основой для предположения, что ген чепрачности является модификацией исходного гена А. Сходство усиливается, если рыжий подпал осветлен. Строго говоря, вариабельность чепрачности определяется двумя независимыми системами модификаторов. С одной стороны — это модификаторы, управляющие вариабельностью чепрака, его интенсивностью, формой и протяженностью. С другой стороны, окрас подпалов определяется руфус-полигенами, как в случае с чисто рыжими собаками. Причем, они не влияют на экспрессию гена чепрачности.
Интересно было бы предположить, что некоторые фенотипические вариации чепрачного окраса обязаны полигенам, влияющим на экспрессию гена А, которые затеняют рыжий окрас превращая его в соболиный. Такие затеняющие полигены могли бы продуцировать некоторые вариации чепрачного окраса, модифицируя экспрессию гена А. К сожалению, никакими экспериментальными данными по этому вопросу мы не располагаем. Интенсивность пигментации подпалов может также варьировать, причем независимо от интенсивности окраса чепрака. Почти наверняка это происходит благодаря действию независимых руфус-полигенов.
В идеале, чепрачными принято называть собак с V-образным темным пятном на спине и боках туловища, простирающимся от хребта до живота. Однако, как распространение, так и интенсивность пигментации этой области может варьировать. При яркой, интенсивной пигментации чепрачный окрас легко идентифицировать, но при более светлой пигментации границы чепрака размыты и наличие его трудно распознать. Фактически, этот окрас стремится иметь сходство с диким волчеобразным. И это является основой для предположения, что ген чепрачности является модификацией исходного гена А. Сходство усиливается, если рыжий подпал осветлен. Строго говоря, вариабельность чепрачности определяется двумя независимыми системами модификаторов. С одной стороны — это модификаторы, управляющие вариабельностью чепрака, его интенсивностью, формой и протяженностью. С другой стороны, окрас подпалов определяется руфус-полигенами, как в случае с чисто рыжими собаками. Причем, они не влияют на экспрессию гена чепрачности.
Интересно было бы предположить, что некоторые фенотипические вариации чепрачного окраса обязаны полигенам, влияющим на экспрессию гена А, которые затеняют рыжий окрас превращая его в соболиный. Такие затеняющие полигены могли бы продуцировать некоторые вариации чепрачного окраса, модифицируя экспрессию гена А. К сожалению, никакими экспериментальными данными по этому вопросу мы не располагаем. Интенсивность пигментации подпалов может также варьировать, причем независимо от интенсивности окраса чепрака. Почти наверняка это происходит благодаря действию независимых руфус-полигенов.
Полигены затенения или соболиного окраса
Эти полигены, судя по названию, вызывает потемнение шерсти. Их наиболее очевидной экспрессией является измелен не обычного А
у-рыжего окраса до соболиного. Большинство, если не все А
у-рыжие особи имеют некоторое количество черно-типированных волосков, как правило, на голове и вдоль спины. Туловище щенка обычно покрыто темными волосами, придавая ему более темный окрас. При взрослении щенка окрас становится все более светлым, пока не будет достигнут окрас взрослого животного.
Когда количество темных волос многочисленно у взрослого животного рыжего окраса, получается соболиный окрас. Степень затененности значительно варьирует, особенно на голове, плечах и спине. Наиболее темные особи могут напоминать осветленных чепрачных. Особенно, если затенение распространяется на бока, напоминая по форме сглаженный чепрак.
Типичный соболиный окрас можно встретить у колли или шотландской овчарки. Показано, что только А у-рыжий окрас конвертируется в соболиный с помощью полигенов затенения. Предполагается, но еще не доказано, что эти полигены не оказывают сходного воздействия на ген е, который, очевидно, эпистатичен по отношению к данным модификаторам.
Когда количество темных волос многочисленно у взрослого животного рыжего окраса, получается соболиный окрас. Степень затененности значительно варьирует, особенно на голове, плечах и спине. Наиболее темные особи могут напоминать осветленных чепрачных. Особенно, если затенение распространяется на бока, напоминая по форме сглаженный чепрак.
Типичный соболиный окрас можно встретить у колли или шотландской овчарки. Показано, что только А у-рыжий окрас конвертируется в соболиный с помощью полигенов затенения. Предполагается, но еще не доказано, что эти полигены не оказывают сходного воздействия на ген е, который, очевидно, эпистатичен по отношению к данным модификаторам.
Белые собаки
Фенотипически, полностью белые собаки могут быть основой для ряда пород или породных вариаций. В принципе, полностью белые собаки имеют различную генотипическую основу. У многих млекопитающих встречается альбинизм. Этот фенотип полностью лишен Пигмента не только в волосах, но и в радужине глаза, кожи, когтях и др. Такие белые особи красноглазы. Однако, когда собаководы говорят о белых собаках, они имеют в виду темноглазых животных. Таких собак нельзя назвать альбиносами, даже если они имеют голубые глаза.
Одно из наиболее ранних предположений о происхождении чисто белых собак состоит в том, что окрас таких особей есть следствие действия гена крайней степени пятнистости. Литтл постулировал существование аллеля s w, который и обуславливает чисто белый окрас. Животные с генотипом s ws wили полностью белые, или имеют незначительные отметины в области ушей и основании хвоста. Постоянная селекция и удаление особей с отметинами из разведения в прошлом привели к тому, что они составляют небольшую часть популяции. Однако, факт, что такие особи все-таки появляются, является хорошим подтверждением существования генотипа s ws w. В результате, Литтл смог предположить, что у бультерьера, большой пиренейской собаки, самоеда и силихэмтерьера имеет место быть генотип s ws w.
Можно также предположить, что чисто белый окрас возникает в результате действия гена шиншилла (c ch), который удаляет почти весь желтый пигмент волос, но не глаз. В результате могут получиться темноглазые белые собаки. Tjebbes и Wriedt (1927) предположили, что такими собаками могут быть самоеды. Возможно, что они ошибались, исходя из более поздней концепции Литтля. Однако, совсем не обязательно, что все самоеды несут s ws w, т.к. данные Литтля получены при исследовании американской популяции самоедов, а Tjebbes и Wriedt наблюдали скандинавскую группу.
Как известно, пигменты в пигментных гранулах могут быть только двух видов: желтый и черный. Синтез черного пигмента запрещают два гена — гены А уи е. Это первый шаг к возникновению белого окраса. Для получения белой собаки нужен еще один ген, который удалял бы уже желтый пигмент. Этим геном является c ch, т.к. именно он, главным образам, запрещает синтез желтого пигмента.
Хотя ген c chописан Литтлем (1957), существование его не бесспорно. На данный момент мы допускаем, что он есть и действует только в отношении желтого пигмента, оставляя черный. Тогда, комбинация A yс c chпродуцирует белый окрас, т.к. A yудаляет черный, a c chудаляет оставшийся желтый. Ген А уиногда не полностью удаляет черный пигмент. Поэтому, возможно получить затененный белый или белый с маской. Ген е более эффективно удаляет черный пигмент. Следовательно, более чистобелые собаки будут скорее c chc chee, чем A y-c chc ch.
Однако, c chможет недостаточно эффективно удалять желтый пигмент. Такие животные могут быть сходны с золотым ретривером бледно-кремового окраса. Поэтому, требуется дополнительная селекция для удаления остатков желтого пигмента и получения чисто-белых собак.
Осветление и фактическое исчезновение желтого пигмента может быть зависимо от осветляющих руфус-полигенов. Белые животные такой категории могут быть отличимы, если они дают собак с кремовым оттенком по всему корпусу, особенно вдоль позвоночника, или получаются от бледно-кремовых родителей. Литтл (1957) считает, что вестхайлендвайт-терьер может служить примером белого окраса, обусловленного генотипом A y-c chc c h.
Ген мраморного окраса М в гомозиготном состоянии дает белый или почти белый окрас с небольшими участками окрашенных волос на голове. Некоторые индивидуумы развиваются в прекрасных собак, но большая часть имеет один или несколько дефектов. Сюда можно отнести глухоту, редукцию глазного яблока, вплоть до полного его отсутствия. В целом, очевидно, что белые животные, имеющие ген мерль (М) не должны скрещиваться.
Ильин (1932) сообщает, что некоторые белые вариации собак обязаны доминантному гену W. Однако, Витни (1947) утверждает, что в его экспериментах на белых собаках ген W ведет себя как рецессивный. Вполне возможно, что они несли ген s w.
Одно из наиболее ранних предположений о происхождении чисто белых собак состоит в том, что окрас таких особей есть следствие действия гена крайней степени пятнистости. Литтл постулировал существование аллеля s w, который и обуславливает чисто белый окрас. Животные с генотипом s ws wили полностью белые, или имеют незначительные отметины в области ушей и основании хвоста. Постоянная селекция и удаление особей с отметинами из разведения в прошлом привели к тому, что они составляют небольшую часть популяции. Однако, факт, что такие особи все-таки появляются, является хорошим подтверждением существования генотипа s ws w. В результате, Литтл смог предположить, что у бультерьера, большой пиренейской собаки, самоеда и силихэмтерьера имеет место быть генотип s ws w.
Можно также предположить, что чисто белый окрас возникает в результате действия гена шиншилла (c ch), который удаляет почти весь желтый пигмент волос, но не глаз. В результате могут получиться темноглазые белые собаки. Tjebbes и Wriedt (1927) предположили, что такими собаками могут быть самоеды. Возможно, что они ошибались, исходя из более поздней концепции Литтля. Однако, совсем не обязательно, что все самоеды несут s ws w, т.к. данные Литтля получены при исследовании американской популяции самоедов, а Tjebbes и Wriedt наблюдали скандинавскую группу.
Как известно, пигменты в пигментных гранулах могут быть только двух видов: желтый и черный. Синтез черного пигмента запрещают два гена — гены А уи е. Это первый шаг к возникновению белого окраса. Для получения белой собаки нужен еще один ген, который удалял бы уже желтый пигмент. Этим геном является c ch, т.к. именно он, главным образам, запрещает синтез желтого пигмента.
Хотя ген c chописан Литтлем (1957), существование его не бесспорно. На данный момент мы допускаем, что он есть и действует только в отношении желтого пигмента, оставляя черный. Тогда, комбинация A yс c chпродуцирует белый окрас, т.к. A yудаляет черный, a c chудаляет оставшийся желтый. Ген А уиногда не полностью удаляет черный пигмент. Поэтому, возможно получить затененный белый или белый с маской. Ген е более эффективно удаляет черный пигмент. Следовательно, более чистобелые собаки будут скорее c chc chee, чем A y-c chc ch.
Однако, c chможет недостаточно эффективно удалять желтый пигмент. Такие животные могут быть сходны с золотым ретривером бледно-кремового окраса. Поэтому, требуется дополнительная селекция для удаления остатков желтого пигмента и получения чисто-белых собак.
Осветление и фактическое исчезновение желтого пигмента может быть зависимо от осветляющих руфус-полигенов. Белые животные такой категории могут быть отличимы, если они дают собак с кремовым оттенком по всему корпусу, особенно вдоль позвоночника, или получаются от бледно-кремовых родителей. Литтл (1957) считает, что вестхайлендвайт-терьер может служить примером белого окраса, обусловленного генотипом A y-c chc c h.
Ген мраморного окраса М в гомозиготном состоянии дает белый или почти белый окрас с небольшими участками окрашенных волос на голове. Некоторые индивидуумы развиваются в прекрасных собак, но большая часть имеет один или несколько дефектов. Сюда можно отнести глухоту, редукцию глазного яблока, вплоть до полного его отсутствия. В целом, очевидно, что белые животные, имеющие ген мерль (М) не должны скрещиваться.
Ильин (1932) сообщает, что некоторые белые вариации собак обязаны доминантному гену W. Однако, Витни (1947) утверждает, что в его экспериментах на белых собаках ген W ведет себя как рецессивный. Вполне возможно, что они несли ген s w.
Окрас радужины глаза
Под цветом глаз обычно подразумевается цвет радужины, т.к. зрачок у большинства собак черный. Как правило, радужина у собак коричневая, интенсивность пигментации которой, варьирует. Глаза могут быть также ореховые или янтарные. Цвет радужины определяется двумя факторами: действием основного гена, который определяет основной цвет и действием модификаторов, которые создают дополнительное разнообразие цветовых вариаций радужины.
Кроме этого, ген коричневого окраса b и ослабитель d также влияют на цвет радужины. У bb-животных она светлее, чем у В-. Такой цвет называют ореховым. В свою очередь, интенсивность его варьирует под действием полигенов. Ослабитель d осветляет глаза до янтарного или даже желтого.
Бернс (1943) описывает радужину голубых собак как дымчатую или дымчато-желтую.
Он также отметил, что цвет глаз обычно окрашенных собак варьирует от темно- до светло-коричневых, и заметил, что собаки со светло-коричневыми глазами при скрещивании не дают щенков с темно-коричневыми глазами. Напротив, от собак с темными глазами можно получить светлоглазых щенков. Однако, утверждать о моногенности этого признака преждевременно. Более очевидно, что интенсивность пигментации детерминирована полигонами, увеличивающими или уменьшающими количество пигмента в радужине. Те же полигены, вероятно, модифицируют пигментацию глаз и у шоколадных и у голубых собак независимо от действия основных генов d и b, вызывая то разнообразие оттенков, которое мы наблюдаем у собак этих окрасов.
Вариации цвета радужины, описанные выше, присущи целому ряду обычно окрашенных собак. Однако, существуют случаи, о которых следует сказать особо, Некоторые, наиболее крайне выраженные альбинотические аллели, могут давать светло-голубые и даже белесые глаза благодаря резкому уменьшению пигмента в радужине. Происходит это в результате смешения цветов крови и остатка пигмента. В случае полного альбинизма радужина становится бесцветной и сквозь нее просвечивает красная кровь сосудов глаза.
Ген мерль (М) может давать голубую радужину, как одно из своих проявлений. Радужина может быть частично или полностью голубой. Редукция пигмента может также касаться как одного, так и обоих глаз. В таком случае собака вырастает разноглазой. Эффект редукции пигмента глаза находится в прямой связи с ослабляющим действием гена М на окрас шерсти, т.к. экспрессивность его варьирует.
Ильин (1932) приводит примеры голубоглазости, которые не зависят от основного окраса. Описанный им признак ведет себя рецессивно по отношению к коричневоглазости. Витни (1947) отмечает сходные случаи у пуделей. Он сообщает, что голубоглазые пуделя могут появляться и у темно- и у светло-коричневоглазых индивидуумов и подтверждает рецессивность этого признака.
Ильиным были описаны случаи, когда нормальное желто-зеленое свечение в темноте помещалось на красное, что являлось рецессивным по отношению к желто-зеленому.
Кроме этого, ген коричневого окраса b и ослабитель d также влияют на цвет радужины. У bb-животных она светлее, чем у В-. Такой цвет называют ореховым. В свою очередь, интенсивность его варьирует под действием полигенов. Ослабитель d осветляет глаза до янтарного или даже желтого.
Бернс (1943) описывает радужину голубых собак как дымчатую или дымчато-желтую.
Он также отметил, что цвет глаз обычно окрашенных собак варьирует от темно- до светло-коричневых, и заметил, что собаки со светло-коричневыми глазами при скрещивании не дают щенков с темно-коричневыми глазами. Напротив, от собак с темными глазами можно получить светлоглазых щенков. Однако, утверждать о моногенности этого признака преждевременно. Более очевидно, что интенсивность пигментации детерминирована полигонами, увеличивающими или уменьшающими количество пигмента в радужине. Те же полигены, вероятно, модифицируют пигментацию глаз и у шоколадных и у голубых собак независимо от действия основных генов d и b, вызывая то разнообразие оттенков, которое мы наблюдаем у собак этих окрасов.
Вариации цвета радужины, описанные выше, присущи целому ряду обычно окрашенных собак. Однако, существуют случаи, о которых следует сказать особо, Некоторые, наиболее крайне выраженные альбинотические аллели, могут давать светло-голубые и даже белесые глаза благодаря резкому уменьшению пигмента в радужине. Происходит это в результате смешения цветов крови и остатка пигмента. В случае полного альбинизма радужина становится бесцветной и сквозь нее просвечивает красная кровь сосудов глаза.
Ген мерль (М) может давать голубую радужину, как одно из своих проявлений. Радужина может быть частично или полностью голубой. Редукция пигмента может также касаться как одного, так и обоих глаз. В таком случае собака вырастает разноглазой. Эффект редукции пигмента глаза находится в прямой связи с ослабляющим действием гена М на окрас шерсти, т.к. экспрессивность его варьирует.
Ильин (1932) приводит примеры голубоглазости, которые не зависят от основного окраса. Описанный им признак ведет себя рецессивно по отношению к коричневоглазости. Витни (1947) отмечает сходные случаи у пуделей. Он сообщает, что голубоглазые пуделя могут появляться и у темно- и у светло-коричневоглазых индивидуумов и подтверждает рецессивность этого признака.
Ильиным были описаны случаи, когда нормальное желто-зеленое свечение в темноте помещалось на красное, что являлось рецессивным по отношению к желто-зеленому.
Цвет мочки носа
В противоположность цвету глаз, окрас мочки носа почти полностью предопределен основными мажорными генами окраса. Интенсивно прокрашенная собака с хорошим здоровьем имеет черную мочку носа. С другой стороны, коричневые животные (bb) имеют коричневую (печеночную) мочку носа, а ослабление по гену d дает асфальтово-серую. Можно наблюдать различные вариации в интенсивности прокрашивания, но, в целом, цвет мочки носа соответствует окрасу животного. Даже в случае красного или желтого окрасов В- или bb- животные могут быть легко различимы по цвету мочки носа.
КАЧЕСТВО ШЕРСТИ
У собак существует значительное количество различных типов шерсти. Однако, различия эти в некоторых случаях скорее видимы, чем реальны. Генов, определяющих качество шерсти может быть всего несколько, а вариации в их экспрессии определяются соответствующими модифицирующими генами. Проиллюстрируем это на примере такого признака, как длинная шерсть. Нет причин считать, что в проявлении этого признака участвует более, чем один ген. Несмотря на это, очевидно, что длинношерстные животные различаются от породы к породе. Эти вариации происходят благодаря полигенам модификации экспрессии гена длинношерстности. Различия в степени длинношерстности закреплены в породах в результате длительного периода их становления.
Для проведения корректного анализа наследуемости длины шерсти следовало бы изолировать различные компоненты качества шерсти и изучать наследственность каждого из них отдельно. К сожалению, в этом направлении сделано крайне мало. Даже у собак с обычным шерстным покровом существует удивительное разнообразие длины волоса, плотности шерсти (количество волос на единицу площади кожи), соотношения числа остевых волос и подшерстка, прямых и имеющих тенденцию к волнистости. Возможно, что перечислены еще не все слагаемые качества шерсти. Многие из них варьируют независимо друг от друга и комбинации их дают большинство типов шерсти.
Т.о., пока не доказано обратное, наследуемость качества шерсти следует считать полигенной.
Описаны случаи, когда бесшерстность наследовалась рецессивно. Поэтому, не стоит считать, что этот признак всегда кодируется доминантным геном Hr.
Бесшерстные мутанты время от времени встречаются среди нормальных собак и некоторые из них могут выживать в более теплом климате или специально отбираться. Если такие животные имеют древнее происхождение, то могут приобрести ауру респектабельности и быть признанными как порода. Насколько это оправдано является спорным вопросом.
Для проведения корректного анализа наследуемости длины шерсти следовало бы изолировать различные компоненты качества шерсти и изучать наследственность каждого из них отдельно. К сожалению, в этом направлении сделано крайне мало. Даже у собак с обычным шерстным покровом существует удивительное разнообразие длины волоса, плотности шерсти (количество волос на единицу площади кожи), соотношения числа остевых волос и подшерстка, прямых и имеющих тенденцию к волнистости. Возможно, что перечислены еще не все слагаемые качества шерсти. Многие из них варьируют независимо друг от друга и комбинации их дают большинство типов шерсти.
Т.о., пока не доказано обратное, наследуемость качества шерсти следует считать полигенной.
Длинная шерсть
Длинная шерсть определяется мутантным геном l, возможно, что этот ген продлевает рост волоса, и он становится в несколько раз длиннее, чем обычный. Экспрессия l модифицирована по ряду показателей. У некоторых шерсть не кажется очень длинной и плотно прилегает к телу. На экспрессию гена l указывает в этом случае длинный украшающий рост волос на конечностях и хвосте, как у английского сеттера, у других шерсть особенно длинная, прямая и шелковистая как у йоркширского терьера. У третьих, она обильная и очень плотная, как у чау-чау. Наконец, у последних она длинная, свалявшаяся и косматая, как у бобтейла. У спаниелей длинная шерсть часто имеет волнистую структуру и сильно развита на конечностях. Особенности наследуемости украшающего волоса будут обсуждаться ниже.Жесткая шерсть
Жесткая шерсть, характерна для эрдельтерьера, таксы, фокстерьера, брюссельского гриффона и дратхаара. Она образуется в результате действия доминантного гена Wh (Whitney, 1947; Winge, 1950). Считается, что у всех жесткошерстных пород работает один и тот же ген, хотя предположение о том, что их больше, можно принять как рабочую гипотезу. Есть наблюдения, что ген Wh задерживает рост шерсти, тогда генотип llWh- дает волосы длиннее, чем LLWh-, но короче, чем llwhwh.Скрученная шерсть
Витни предположил, что курчавая или скрученная шерсть ирландского водяного спаниеля наследуется рецессивно по отношению к прямой. Условно, ген обозначен k.Курчавая шерсть
Считалось, что курчавая шерсть ретривера является короткошерстной вариацией скрученной шерсти ирландского водяного спаниеля. Бернс и Фразер (1966) сообщают, что при вязке курчавого ретривера с дратхааром щенки имели такую же курчавую шерсть, как у ретривера. Следовательно, курчавая шерсть, возможно, определяется разными генами у водяного спаниеля и ретривера.Волнистая шерсть
Витни описал особенности некоторых скрещиваний, которые указывают на то, что волнистая шерсть кокер-спаниелей и ряда других пород, возможно, наследуется рецессивно по отношению к прямой. Однако, это не более, чем предположение. Сведений для прямых доказательств недостаточно, поэтому, предполагаемый ген может быть предварительно обозначен, как wa.Шерсть с завитком
Пиллингом была описана линия кокер-спаниеля с необычными кольцами волос на плечах и на боках. Некоторые собаки имели, лишь одно кольцо, которое могло располагаться на голове или шее. Этот признак наследовался рецессивно и ген, его определяющий, обозначен как wo.«Складчатая» шерсть
Витни описал необычную аномалию кожи (в виде складок) или направления роста волос у щенков бладхаунда. В возрасте 1-7 дней у них появлялись длинные, регулярно расположенные волосы, растущие от шеи до задних конечностей. В семь дней они исчезают, и шерсть по всему телу становится нормальной. Результаты скрещиваний позволяют предположить, что этот признак наследуется рецессивно. Ген, кодирующий его обозначен как rp.Украшающая шерсть
Украшающая шерсть или кайма из длинных волос на конечностях и нижней поверхности хвоста является характерной чертой длинношерстных собак. Степень развития этого признака сильно варьирует. Битнер (1947) сообщает, что кроссы между ирландским водяным спаниелем (с незначительным развитием украшающего волоса на хвосте) и кокером или ирландским сеттером (с обильной оброслостью) давали потомство с обильной оброслостью. Это указывает на доминантность признака, но недостаточно для того, чтобы ответить на вопрос, кодируется ли этот признак одним независимым геном или развивается в результате воздействия модифицирующих полигенов на ген длинношерстности. Последнее наиболее вероятно и может служить примером того, как экспрессия гена длинношерстности может быть модифицирована другими генами.Особенность роста волос разного цвета
Витни (1947) отмечает, что у длинношерстных, пятнистых собак черные и белые волосы разной длины. У кокера и английского сеттера черные волосы длиннее, чем белые. Черные волосы также длиннее, чем рыжие. Бернс и Фразер (1966) наблюдали подобное и добавляют, что белые волосы в отдельных случаях могут быть той же длины, что и черные, но мягче по структуре. Рыжие волосы также мягче и шелковистее, чем черные.Бесшерстность
Бесшерстность — это явная аномалия. Однако этот признак следует рассмотреть в настоящей работе, т.к. он является определяющим для таких пород, как китайская или мексиканская голые собаки. Эти животные не всегда полностью бесшерстны, а имеют варьирующее количество волос на голове и конечностях. Летард (1930) показал, что эти породы образовались в результате действия доминантного гена Hr, который в гомозиготном состоянии летален. Следовательно, все бесшерстные особи этих пород гетерозиготны по Hr (Hr hr).Описаны случаи, когда бесшерстность наследовалась рецессивно. Поэтому, не стоит считать, что этот признак всегда кодируется доминантным геном Hr.
Бесшерстные мутанты время от времени встречаются среди нормальных собак и некоторые из них могут выживать в более теплом климате или специально отбираться. Если такие животные имеют древнее происхождение, то могут приобрести ауру респектабельности и быть признанными как порода. Насколько это оправдано является спорным вопросом.
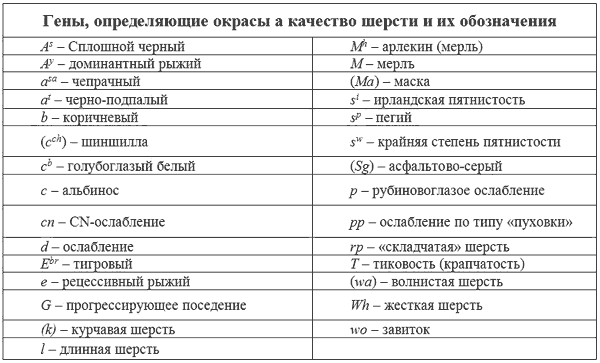
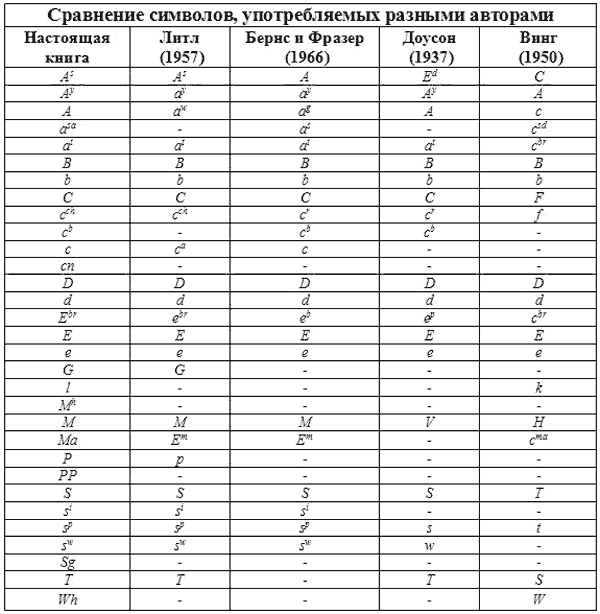
СРАВНИТЕЛЬНАЯ СИМВОЛИКА ГЕНОВ
Читатели, которые интересуются литературой по генетике, рано или поздно сталкиваются с проблемой путаницы в обозначениях генов. Дело в том, что различные авторы пользуются различными символами для обозначения одного и того же гена. Это прискорбно, но неизбежно. Исследователи сразу же не согласовали стандартную номенклатуру генов и называли новые описанные гены как им заблагорассудится. Настоящий раздел книги является попыткой свести накопленное в единую систему.
Впервые на это обратил внимание Доусон (1937), который попытался привести в соответствие предыдущие достижения. Собранный им материал выдержал проверку временем. Последующий вклад Литтля (1957), а также Бернса и Фразера (1966) укладывался в основное русло генетики млекопитающих. Увы, это не относится к работам Винга (1951). Его система полностью противоречит предшествующим и достаточно запутана. Он предлагает ряд генов, в которых нет необходимости. Некоторые гены Винг разместил не в тех аллельных сериях, где следовало бы.
Впервые на это обратил внимание Доусон (1937), который попытался привести в соответствие предыдущие достижения. Собранный им материал выдержал проверку временем. Последующий вклад Литтля (1957), а также Бернса и Фразера (1966) укладывался в основное русло генетики млекопитающих. Увы, это не относится к работам Винга (1951). Его система полностью противоречит предшествующим и достаточно запутана. Он предлагает ряд генов, в которых нет необходимости. Некоторые гены Винг разместил не в тех аллельных сериях, где следовало бы.
Таблица 11
Таблица 12
Заключение
Если сравнить окрасы собак, обусловленные генами Ay и е, можно заметить, что в первом случае собаки более интенсивно пигментированы. Однако, не стоит считать, что все красные собаки несут Ay, а все более бледные — ее. Там где возникают сомнения по поводу происхождения окраса можно допустить, что он скорее обусловлен А