Страница:
1960 год
Открытие РНК–полимеразы С. Б. Вейсом, Дж. Гурвицем и А. Сти–венсом.
И. А. Рапопорт сообщил об открытии «супермутагенов».
1961 год
В работах М. У. Ниренберга, Р. У. Холли и X. Г. Кораны начата расшифровка «языка жизни» – кода, которым в ДНК записана информация о структуре белковых молекул. В 1968 году все трое разделили Нобелевскую премию по физиологии и медицине, которая была присуждена им «за расшифровку генетического кода и его функционирования в синтезе белков».
Ф. Жакоб и Ж. Моно пришли к выводу о существовании двух групп генов – структурных, отвечающих за синтез специфических (ферментных) белков, и регуляторных, осуществляющих контроль за активностью структурных генов. В 1965 г. Нобелевская премия по физиологии и медицине присуждена А. М. Львову, Ф. Жакобу и Ж. Моно за открытие генетической регуляции синтеза ферментов и вирусов.
1962 год
Дж. Гёрдон осуществил первое клонирование животного организма (лягушка).
Дж. Кэндрью и М. Перутц были удостоены Нобелевской премии по химии за впервые осуществленную расшифровку трехмерной структуры белков миоглобина и гемоглобина.
1965 год
Р. Б. Хесин показал, что регуляция синтеза белка осуществляется путем включения и выключения транскрипции генов.
1966 год
Б. Вейс и С. Рихардсон открывают фермент ДНК–лигазу.
1969 год
Х. Г. Корана синтезировал химическим путем первый ген.
1970 год
Открытие обратной транскриптазы, фермента, синтезирующего ДНК с использованием комплементарной РНК в качестве матрицы. Это было сделано будущими Нобелевскими лауреатами по физиологии и медицине (1975) Г. Теминым и Д. Балтимором.
Выделена первая рестриктаза – фермент, разрезающий ДНК в строго определенных местах. За это открытие в 1978 году Нобелевская премия по физиологии и медицине была присуждена Д. Натансу, Х. Смиту и В. Арберу.
1972 год
В лаборатории Пола Берга получены первые рекомбинантные ДНК (Нобелевская премия по химии за 1980 г. вручена П. Бергу и Г. Бойеру). Заложены основы генной инженерии.
1973 год
С. Коэн и Г. Бойер разработали стратегию переноса генов в бактериальную клетку.
1974 год
С. Милстайн и Г. Келер создали технологию получения монокло–нальных антител. Ровно десять лет спустя они (вместе с Н. К. Ерне) получили за это Нобелевскую премию по физиологии и медицине.
Р. Д. Корнберг описывает структуру хроматина (нуклеосомы).
1975 год
С. Тонегава показал различное расположение генов, кодирующих вариабельную и константную часть иммуноглобулинов, в ДНК эмбриональных и миелоидных клеток, что дало основание для вывода о перегруппировках генов иммуноглобулинов при образовании клеток иммунной системы (Нобелевская премия по физиологии и медицине в 1987 г.).
Осуществлено первое клонирование кДНК.
Е. Саузерн описал метод переноса фрагментов ДНК на нитроцел–люлозные фильтры, метод получил название Саузерн–блот гибридизации.
1976 год
Открытие у животных (на примере дрозофилы) «прыгающих генов», сделанное Д. Хогнессом (США) и российскими учеными во главе с Г. П. Георгиевым и В. А. Гвоздевым.
Основана первая генно–инженерная компания (Genentech), использующая технологию рекомбинантных ДНК для производства различных ферментов и лекарственных средств.
Д. М. Бишоп и Г. Э. Вармус сообщили, что онкоген в вирусе представляет собою не истинный вирусный ген, а клеточный ген, который вирус «подхватил» когда–то давно в ходе репликации в клетках и теперь сохраняет в измененном мутациями виде. Было также показано, что его предшественник, клеточный протоонкоген, в здоровой клетке играет важнейшую роль—управляет ее ростом и делением. В 1989 г. оба этих ученых получили Нобелевскую премию по физиологии и медицине за фундаментальные исследование канцерогенных генов опухоли.
1977 год
Опубликованы быстрые методы определения (секвенирования) длинных нуклеотидных последовательностей ДНК (У. Гилберт и А. Максам; Ф. Сенгер с соавт.). Появилось реальное средство анализа структуры генов как основа для понимания их функций. В 1980 году У. Гильберт и Ф. Сенгер совместно с П. Бергом получили Нобелевскую премию по химии «за существенный вклад в установление первичной структуры ДНК; за фундаментальные исследования биохимических свойств нуклеиновых кислот, в том числе рекомбинант–ных ДНК».
Полностью секвенирован геном бактериофага φΧ174 (5386 п. н.).
Секвенирован первый ген человека – ген, кодирующий белок хорионный соматомаммотропин.
П. Шарп и Р. Робертс показали, что гены у аденовирусов (позднее выяснилось, что и у эукариотических организмов) имеют мозаичную экзон–интронную структуру, и открыли явление сплайсинга (Нобелевская премия по физиологии и медицине в 1993 г.).
К. Итакура с соавт. синтезируют химически ген соматостатина человека и осуществляет искусственный синтез гормона соматоста–тина в клетках кишечной палочки E. coli.
1978 год
Компания Genentech осуществила перенос эукариотического гена инсулина в бактериальную клетку, где на нем синтезирован белок – проинсулин.
Определена полная последовательность нуклеотидов ДНК вируса SV40 и фага fd.
1979 год
Показано, что химически трансформированные клетки содержат активированный онкоген BAS.
1980 год
Дж. Гордоном с соавт. получена первая трансгенная мышь. В про–нуклеус оплодотворенного одноклеточного эмбриона микроинъекцией введен ген тимидин–киназы вируса простого герпеса и показано, что этот ген работает во всех соматических клетках мыши. С тех пор трансгеноз стал основным подходом как для фундаментальных исследований, так и для решения практических задач сельского хозяйства и медицины.
1981 год
Определена полная нуклеотидная последовательность митохон–дриальной ДНК человека.
Несколько независимых исследовательских групп сообщили об открытии человеческих онкогенов.
1982 год
Определена полная нуклеотидная последовательность бактериофага λ (48502 п. н.).
Показано, что РНК может обладать каталитическими свойствами, как и белок.
1983 год
С помощью биоинформатики найдена гомология фактора роста PDGF с известным онкобелком, кодируемым онкогеном SIS.
Показано, что разные онкогены кооперируют при опухолевой трансформации клеток.
Ген болезни Хантигтона локализован на хромосоме 4 человека.
1984 год
У. Мак–Гиннис открыл гомеотические (Hox) регуляторные гены, ответственные за построение общего плана тела животных.
А. Джеффрис создает метод геномной дактилоскопии, в котором нуклеотидные последовательности ДНК используются для идентификации личности.
1985 год
Создание К. Б. Мюллисом революционизирующей технологии – полимеразной цепной реакции, ПЦР – наиболее чувствительного до сих пор метода детектирования ДНК. Эта технология получила широкое распространение (Нобелевская премия по химии за 1993 г.).
Клонирование и определение нуклеотидной последовательности ДНК, выделенной из древней египетской мумии.
1986 год
Клонирование гена RB– первого антионкогена – супрессора опухолей. Начало эпохи массированного клонирования генов опухоле–образования.
1987 год
Созданы первые дрожжевые искусственные хромосомы – YAC (Yeast Artificial Chromosomes). Они сыграют большую роль как векторы для клонирования больших фрагментов геномов.
1988 год
Создание международного проекта «Геном человека», поставившего своей целью полное секвенирование ДНК человека.
Под эгидой Комитета по науке и технике в СССР начала работу программа «Геном человека», которую возглавил Научный совет по геномной программе во главе с академиком А. А. Баевым.
Показана возможность анализа митохондриальной ДНК из очень древних образцов при исследовании мозга человека давностью 7000 лет.
Предложен метод «нокаута» генов.
1989 год
Т. Р. Чех и С. Альтман получили Нобелевскую премию по химии за открытие каталитических свойств некоторых природных РНК (рибозимов).
1990 год
Создана Международная организация по изучению генома человека (HUGO), вице–президентом которой в течение нескольких лет был российский академик А. Д. Мирзабеков.
Ф. Коллинз и Л. – Ч. Тсуи идентифицировали первый ген человека ( CFTR), ответственный за наследственное заболевание (кистозный фиброз), который расположен на хромосоме 7.
В. Андерсоном осуществлено первое успешное применение генной терапии для лечения больной с наследственным иммунодефицитом.
Определена полная последовательность генома вируса оспо–вакцины (192 т. п. н.).
1992 год
Э. Кребсу и Э. Фишеру присуждена Нобелевская премия по физиологии и медицине за открытие обратимого фосфорилирования белков как важного регулирующего механизма клеточного метаболизма.
1995 год
Определена полная последовательность генома первого самостоятельно существующего организма – бактерии Haemophilus influenzae (1 830 137 п. н.). Становление геномики как самостоятельного раздела генетики.
1997 год
Определена полная последовательность нуклеотидов геномов кишечной палочки E. coli и дрожжей Saccharomyces cerevisiae.
Нобелевская премия по физиологии и медицине присуждена американцу С. Прузинеру за вклад в изучение болезнетворного агента белковой природы, приона, вызывающего губчатую энцефалопатию, или «коровье бешенство» у крупного рогатого скота.
Я. Вильмут с сотрудниками впервые клонировали млекопитающее – овцу Долли.
1998 год
Определена полная нуклеотидная последовательность первого высшего организма – нематоды Caenorhabditis elegans.
У нематоды C. elegans обнаружен механизм РНК–интерференции.
1999 год
Полностью секвенирована ДНК хромосомы 22 человека.
Роберт Фурчготт, Луис Игнарро и Ферид Мурад получили Нобелевскую премию за открытие роли оксида азота в качестве сигнальной молекулы (то есть, регулятора и переносчика сигналов) сердечно–сосудистой системы.
Клонировали мышь и корову.
2000 год
Вчерне завершено полное секвенирование генома человека и дрозофилы.
Нобелевская премия по физиологии и медицине присуждена А. Карлссону, П. Грингарду и Э. Кенделу за открытие, касающееся «передачи сигналов в нервной системе».
Клонировали свинью.
2001 год
Нобелевская премия по физиологии и медицине присуждена Л. Хартвеллу, Т. Ханту и П. Нерсу за открытие ключевых регуляторов клеточного цикла.
2002 год
Полностью секвенирован геном мыши.
Нобелевская премия по физиологии и медицине присуждена С. Бреннеру, Р. Хорвитцу и Дж. Салстону за их открытия в области генетического регулирования развития органов и запрограммированной клеточной смерти.
ГЕНЫ – ОСНОВНОЙ ТЕКСТ ГЕНОМА
«Жи, ши – пиши через и»
Знаки препинания (Генетическая пунктуация)
Гены построены из кусков
Открытие РНК–полимеразы С. Б. Вейсом, Дж. Гурвицем и А. Сти–венсом.
И. А. Рапопорт сообщил об открытии «супермутагенов».
1961 год
В работах М. У. Ниренберга, Р. У. Холли и X. Г. Кораны начата расшифровка «языка жизни» – кода, которым в ДНК записана информация о структуре белковых молекул. В 1968 году все трое разделили Нобелевскую премию по физиологии и медицине, которая была присуждена им «за расшифровку генетического кода и его функционирования в синтезе белков».
Ф. Жакоб и Ж. Моно пришли к выводу о существовании двух групп генов – структурных, отвечающих за синтез специфических (ферментных) белков, и регуляторных, осуществляющих контроль за активностью структурных генов. В 1965 г. Нобелевская премия по физиологии и медицине присуждена А. М. Львову, Ф. Жакобу и Ж. Моно за открытие генетической регуляции синтеза ферментов и вирусов.
1962 год
Дж. Гёрдон осуществил первое клонирование животного организма (лягушка).
Дж. Кэндрью и М. Перутц были удостоены Нобелевской премии по химии за впервые осуществленную расшифровку трехмерной структуры белков миоглобина и гемоглобина.
1965 год
Р. Б. Хесин показал, что регуляция синтеза белка осуществляется путем включения и выключения транскрипции генов.
1966 год
Б. Вейс и С. Рихардсон открывают фермент ДНК–лигазу.
1969 год
Х. Г. Корана синтезировал химическим путем первый ген.
1970 год
Открытие обратной транскриптазы, фермента, синтезирующего ДНК с использованием комплементарной РНК в качестве матрицы. Это было сделано будущими Нобелевскими лауреатами по физиологии и медицине (1975) Г. Теминым и Д. Балтимором.
Выделена первая рестриктаза – фермент, разрезающий ДНК в строго определенных местах. За это открытие в 1978 году Нобелевская премия по физиологии и медицине была присуждена Д. Натансу, Х. Смиту и В. Арберу.
1972 год
В лаборатории Пола Берга получены первые рекомбинантные ДНК (Нобелевская премия по химии за 1980 г. вручена П. Бергу и Г. Бойеру). Заложены основы генной инженерии.
1973 год
С. Коэн и Г. Бойер разработали стратегию переноса генов в бактериальную клетку.
1974 год
С. Милстайн и Г. Келер создали технологию получения монокло–нальных антител. Ровно десять лет спустя они (вместе с Н. К. Ерне) получили за это Нобелевскую премию по физиологии и медицине.
Р. Д. Корнберг описывает структуру хроматина (нуклеосомы).
1975 год
С. Тонегава показал различное расположение генов, кодирующих вариабельную и константную часть иммуноглобулинов, в ДНК эмбриональных и миелоидных клеток, что дало основание для вывода о перегруппировках генов иммуноглобулинов при образовании клеток иммунной системы (Нобелевская премия по физиологии и медицине в 1987 г.).
Осуществлено первое клонирование кДНК.
Е. Саузерн описал метод переноса фрагментов ДНК на нитроцел–люлозные фильтры, метод получил название Саузерн–блот гибридизации.
1976 год
Открытие у животных (на примере дрозофилы) «прыгающих генов», сделанное Д. Хогнессом (США) и российскими учеными во главе с Г. П. Георгиевым и В. А. Гвоздевым.
Основана первая генно–инженерная компания (Genentech), использующая технологию рекомбинантных ДНК для производства различных ферментов и лекарственных средств.
Д. М. Бишоп и Г. Э. Вармус сообщили, что онкоген в вирусе представляет собою не истинный вирусный ген, а клеточный ген, который вирус «подхватил» когда–то давно в ходе репликации в клетках и теперь сохраняет в измененном мутациями виде. Было также показано, что его предшественник, клеточный протоонкоген, в здоровой клетке играет важнейшую роль—управляет ее ростом и делением. В 1989 г. оба этих ученых получили Нобелевскую премию по физиологии и медицине за фундаментальные исследование канцерогенных генов опухоли.
1977 год
Опубликованы быстрые методы определения (секвенирования) длинных нуклеотидных последовательностей ДНК (У. Гилберт и А. Максам; Ф. Сенгер с соавт.). Появилось реальное средство анализа структуры генов как основа для понимания их функций. В 1980 году У. Гильберт и Ф. Сенгер совместно с П. Бергом получили Нобелевскую премию по химии «за существенный вклад в установление первичной структуры ДНК; за фундаментальные исследования биохимических свойств нуклеиновых кислот, в том числе рекомбинант–ных ДНК».
Полностью секвенирован геном бактериофага φΧ174 (5386 п. н.).
Секвенирован первый ген человека – ген, кодирующий белок хорионный соматомаммотропин.
П. Шарп и Р. Робертс показали, что гены у аденовирусов (позднее выяснилось, что и у эукариотических организмов) имеют мозаичную экзон–интронную структуру, и открыли явление сплайсинга (Нобелевская премия по физиологии и медицине в 1993 г.).
К. Итакура с соавт. синтезируют химически ген соматостатина человека и осуществляет искусственный синтез гормона соматоста–тина в клетках кишечной палочки E. coli.
1978 год
Компания Genentech осуществила перенос эукариотического гена инсулина в бактериальную клетку, где на нем синтезирован белок – проинсулин.
Определена полная последовательность нуклеотидов ДНК вируса SV40 и фага fd.
1979 год
Показано, что химически трансформированные клетки содержат активированный онкоген BAS.
1980 год
Дж. Гордоном с соавт. получена первая трансгенная мышь. В про–нуклеус оплодотворенного одноклеточного эмбриона микроинъекцией введен ген тимидин–киназы вируса простого герпеса и показано, что этот ген работает во всех соматических клетках мыши. С тех пор трансгеноз стал основным подходом как для фундаментальных исследований, так и для решения практических задач сельского хозяйства и медицины.
1981 год
Определена полная нуклеотидная последовательность митохон–дриальной ДНК человека.
Несколько независимых исследовательских групп сообщили об открытии человеческих онкогенов.
1982 год
Определена полная нуклеотидная последовательность бактериофага λ (48502 п. н.).
Показано, что РНК может обладать каталитическими свойствами, как и белок.
1983 год
С помощью биоинформатики найдена гомология фактора роста PDGF с известным онкобелком, кодируемым онкогеном SIS.
Показано, что разные онкогены кооперируют при опухолевой трансформации клеток.
Ген болезни Хантигтона локализован на хромосоме 4 человека.
1984 год
У. Мак–Гиннис открыл гомеотические (Hox) регуляторные гены, ответственные за построение общего плана тела животных.
А. Джеффрис создает метод геномной дактилоскопии, в котором нуклеотидные последовательности ДНК используются для идентификации личности.
1985 год
Создание К. Б. Мюллисом революционизирующей технологии – полимеразной цепной реакции, ПЦР – наиболее чувствительного до сих пор метода детектирования ДНК. Эта технология получила широкое распространение (Нобелевская премия по химии за 1993 г.).
Клонирование и определение нуклеотидной последовательности ДНК, выделенной из древней египетской мумии.
1986 год
Клонирование гена RB– первого антионкогена – супрессора опухолей. Начало эпохи массированного клонирования генов опухоле–образования.
1987 год
Созданы первые дрожжевые искусственные хромосомы – YAC (Yeast Artificial Chromosomes). Они сыграют большую роль как векторы для клонирования больших фрагментов геномов.
1988 год
Создание международного проекта «Геном человека», поставившего своей целью полное секвенирование ДНК человека.
Под эгидой Комитета по науке и технике в СССР начала работу программа «Геном человека», которую возглавил Научный совет по геномной программе во главе с академиком А. А. Баевым.
Показана возможность анализа митохондриальной ДНК из очень древних образцов при исследовании мозга человека давностью 7000 лет.
Предложен метод «нокаута» генов.
1989 год
Т. Р. Чех и С. Альтман получили Нобелевскую премию по химии за открытие каталитических свойств некоторых природных РНК (рибозимов).
1990 год
Создана Международная организация по изучению генома человека (HUGO), вице–президентом которой в течение нескольких лет был российский академик А. Д. Мирзабеков.
Ф. Коллинз и Л. – Ч. Тсуи идентифицировали первый ген человека ( CFTR), ответственный за наследственное заболевание (кистозный фиброз), который расположен на хромосоме 7.
В. Андерсоном осуществлено первое успешное применение генной терапии для лечения больной с наследственным иммунодефицитом.
Определена полная последовательность генома вируса оспо–вакцины (192 т. п. н.).
1992 год
Э. Кребсу и Э. Фишеру присуждена Нобелевская премия по физиологии и медицине за открытие обратимого фосфорилирования белков как важного регулирующего механизма клеточного метаболизма.
1995 год
Определена полная последовательность генома первого самостоятельно существующего организма – бактерии Haemophilus influenzae (1 830 137 п. н.). Становление геномики как самостоятельного раздела генетики.
1997 год
Определена полная последовательность нуклеотидов геномов кишечной палочки E. coli и дрожжей Saccharomyces cerevisiae.
Нобелевская премия по физиологии и медицине присуждена американцу С. Прузинеру за вклад в изучение болезнетворного агента белковой природы, приона, вызывающего губчатую энцефалопатию, или «коровье бешенство» у крупного рогатого скота.
Я. Вильмут с сотрудниками впервые клонировали млекопитающее – овцу Долли.
1998 год
Определена полная нуклеотидная последовательность первого высшего организма – нематоды Caenorhabditis elegans.
У нематоды C. elegans обнаружен механизм РНК–интерференции.
1999 год
Полностью секвенирована ДНК хромосомы 22 человека.
Роберт Фурчготт, Луис Игнарро и Ферид Мурад получили Нобелевскую премию за открытие роли оксида азота в качестве сигнальной молекулы (то есть, регулятора и переносчика сигналов) сердечно–сосудистой системы.
Клонировали мышь и корову.
2000 год
Вчерне завершено полное секвенирование генома человека и дрозофилы.
Нобелевская премия по физиологии и медицине присуждена А. Карлссону, П. Грингарду и Э. Кенделу за открытие, касающееся «передачи сигналов в нервной системе».
Клонировали свинью.
2001 год
Нобелевская премия по физиологии и медицине присуждена Л. Хартвеллу, Т. Ханту и П. Нерсу за открытие ключевых регуляторов клеточного цикла.
2002 год
Полностью секвенирован геном мыши.
Нобелевская премия по физиологии и медицине присуждена С. Бреннеру, Р. Хорвитцу и Дж. Салстону за их открытия в области генетического регулирования развития органов и запрограммированной клеточной смерти.
ГЕНЫ – ОСНОВНОЙ ТЕКСТ ГЕНОМА
Гены – это атомы наследственности.
С. Бензер
Орлы летают одиноко, бараны пасутся стадами.
Ф. Сидни
Необходимо отметить, что термин «ген» многие, даже неспециалисты, воспринимают легко без всякого специального пояснения. Слишком много и давно об этом говорят и довольно часто используют этот термин в разных ситуациях. И всем вроде бы все ясно: ген – это участок ДНК, который кодирует определенный белок. Согласно центральной догме молекулярной биологии, информация, записанная в ДНК с помощью четырехбуквенного нуклеотидного алфавита, переводится простым механическим способом на другой, аминокислотный, алфавит из 20 букв, которым записывается строение белковых молекул. «Один ген – один белок» – вот каким был многие годы основополагающий тезис молекулярных генетиков. Для того, чтобы ген правильно работал, необходимы специальные знаки, обозначающие начало и конец инструкции для синтеза сначала мРНК, а потом белка. Для того, чтобы началась считывание (транскрипция) гена, в первую очередь необходим структурный элемент, получивший название промотора (старт–сигнал транскрипции). Для окончания транскрипции существует другой специальный сигнал. Обязательными элементами гена являются также сигналы, определяющие начало и конец трансляции мРНК на рибосомах. Все эти старт–и стоп–сигналы записаны в ДНК с помощью того же самого четырехбуквенного алфавита, что и информация для построения молекулы белка. Между двумя такими знаками препинания записаны инструкции для синтеза белка. В таком упрощенном виде ген определяют как последовательность нуклеотидов, расположенных между старт–сигналом и стоп–сигналом и кодирующих одну белковую цепь (полипептид). Однако на самом деле все обстоит намного сложнее. Кроме генов, кодирующих белки, существуют гены, кодирующие РНК, которые выполняют в клетке только структурную роль и не транслируются. Более того, в последние годы было обнаружено большое число ранее неизвестных РНК, которые выполняют в клетке специальные регуляторные функции. Обо всем этом мы поговорим далее. Современное понимание того, что же из себя представляет ген, складывалось на протяжении всех этих долгих лет, наполняясь постепенно конкретным и обобщенным содержанием, преодолевая сомнения, заблуждения и неожиданные повороты. В результате на сегодняшний день трудно дать однозначное определение термину «ген», хотя вроде бы этот вопрос в общем виде был решен Менделем почти 140 лет назад.
Конечно же, в секвенированном ДНКовом тексте генома человека в первую очередь внимание ученых было обращено на участки, кодирующие белки, то есть на белок–кодирующие гены. Отметим, что первый ген человека – ген, кодирующий белок со сложным названием хорионный соматомаммотропин, – был секвенирован еще в 1977 году. А что же выяснилось в результате секвенирования всего генома человека? Оказалось, что в геноме человека нуклеотидные последовательности, кодирующие белки (экзоны), занимают лишь 1,1—1,4% от длины всех молекул ДНК. Но именно эти мизерные проценты ДНКового текста человека и стали объектом пристального внимания. Для сравнения отметим, что у червя C. elegans доля таких последовательностей в геноме составляет 27%, у мухи дрозофилы – 20%, у дрожжей – 70%, а у бактерий – 86%. То есть, просматривается закономерность: по мере усложнения организмов доля кодирующих белки участков ДНК в их геномах резко падает.
Генам при их открытии, как людям при рождении, сразу же присваивают различные имена. Без этого невозможно было бы разобраться, о чем идет речь и что изучают разные ученые в каждом конкретном случае. Правда, путаница в этом вопросе пока еще существует, особенно если ген одновременно открывают в разных лабораториях и при этом называют по–разному. Известно, что в лингвистике существует целая наука об именах людей – ономастика. Эта наука давно уже отметила влияние разных факторов и причин на формирование имен, связь с историей, географией и многими другими науками, а также процессами, протекающими в обществе. Так, в постреволюционные годы появились имена Вилен (В. И. Ленин), Даздраперма (Да здравствует первое мая), при Б. Ельцине появились Борели, а при В. Путине уже официально зарегистрированы такие имена, как Путилла, Владпут и даже Затерсор (замочим террористов в сортире).
Конечно, и на имена, присваиваемые генам, также влияют разные обстоятельства. Но вообще–то этот процесс довольно стихийный. Нередко наименование гена связывают с выполняемой им функцией (например, дистрофин – ген, мутации в котором ответственны за дистрофию мышц). В других случаях используют особенности структуры гена (например, присутствие специфической последовательности в гене, называемой homeo box, автоматически приводит к имени Hox). Иногда авторы напрягают свою фантазию, и тогда появляются такие экзотические имена, как харакири (ген–самоубийца), икарус, vang (по имени художника ван Гога) или indy ( первые буквы от слов I'm not dead yet – я еще не умер – название для гена, который влияет на продолжительность жизни). Ген SHH назван в честь популярной видеоигры Соника Хеджхога, (англ. hedgehog —ежик). Некоторые гены называются именами богов (например, гефестин – от греческого бога огня и кузнечного дела Гефеста). На практике генетики используют сокращенные обозначения генов, состоящие обычно из 3—5 латинских букв. Поскольку функции многих генов еще не известны, а удается только определить, что какая–то конкретная область генома человека способна теоретически кодировать белок, то часто пока еще перед названиями генов появляется слово «гипотетический».
Анализ генома человека позволил на конец 2002 года обнаружить порядка 30 000—40 000 генов, кодирующих белки. Эти гены сильно отличаются друг от друга по размерам. Подсчитано, что средняя длина гена у человека составляет около 27 тыс. п. н. Такой усредненный ген содержит 9 экзонов (средний размер каждого около 150 п. н.) и 8 интронов (средний размер каждого чуть меньше 3400 п.н.). Но это лишь средние значения. Самые короткие гены содержат всего два десятка букв–нуклеотидов, например, гены эндорфинов – полипептидов, вызывающих ощущение удовольствия. В то же время самый длинный ген, кодирующий один из белков мышц, – миодистрофин содержит 2,4 млн. п. н.
Кроме того, гены не равномерно распределены между разными хромосомами. При средней плотности один ген на 100 т. п. н. генома их содержание в хромосоме 19 составляет около 2 на 100 т. п.н., а в хромосомах 2, 13 и Y – хромосоме – от 0,15 до 0,7. Если сравнить плотность генов с плотностью расселения людей, то Y – хромосома напоминает нашу Сибирь, а хромосома 19 – Европейскую часть России. Для сравнения: в геноме бактерий содержится свыше 1000 генов на 1,0 млн. п. н., у дрожжей около 450 генов на 1,0 млн. п. н., а у червя C. elegans – около 200. Следовательно, плотность расположения генов на единицу длины генома заметно падает по мере эволюционного усложнения организмов.
Конечно же, в секвенированном ДНКовом тексте генома человека в первую очередь внимание ученых было обращено на участки, кодирующие белки, то есть на белок–кодирующие гены. Отметим, что первый ген человека – ген, кодирующий белок со сложным названием хорионный соматомаммотропин, – был секвенирован еще в 1977 году. А что же выяснилось в результате секвенирования всего генома человека? Оказалось, что в геноме человека нуклеотидные последовательности, кодирующие белки (экзоны), занимают лишь 1,1—1,4% от длины всех молекул ДНК. Но именно эти мизерные проценты ДНКового текста человека и стали объектом пристального внимания. Для сравнения отметим, что у червя C. elegans доля таких последовательностей в геноме составляет 27%, у мухи дрозофилы – 20%, у дрожжей – 70%, а у бактерий – 86%. То есть, просматривается закономерность: по мере усложнения организмов доля кодирующих белки участков ДНК в их геномах резко падает.
Генам при их открытии, как людям при рождении, сразу же присваивают различные имена. Без этого невозможно было бы разобраться, о чем идет речь и что изучают разные ученые в каждом конкретном случае. Правда, путаница в этом вопросе пока еще существует, особенно если ген одновременно открывают в разных лабораториях и при этом называют по–разному. Известно, что в лингвистике существует целая наука об именах людей – ономастика. Эта наука давно уже отметила влияние разных факторов и причин на формирование имен, связь с историей, географией и многими другими науками, а также процессами, протекающими в обществе. Так, в постреволюционные годы появились имена Вилен (В. И. Ленин), Даздраперма (Да здравствует первое мая), при Б. Ельцине появились Борели, а при В. Путине уже официально зарегистрированы такие имена, как Путилла, Владпут и даже Затерсор (замочим террористов в сортире).
Конечно, и на имена, присваиваемые генам, также влияют разные обстоятельства. Но вообще–то этот процесс довольно стихийный. Нередко наименование гена связывают с выполняемой им функцией (например, дистрофин – ген, мутации в котором ответственны за дистрофию мышц). В других случаях используют особенности структуры гена (например, присутствие специфической последовательности в гене, называемой homeo box, автоматически приводит к имени Hox). Иногда авторы напрягают свою фантазию, и тогда появляются такие экзотические имена, как харакири (ген–самоубийца), икарус, vang (по имени художника ван Гога) или indy ( первые буквы от слов I'm not dead yet – я еще не умер – название для гена, который влияет на продолжительность жизни). Ген SHH назван в честь популярной видеоигры Соника Хеджхога, (англ. hedgehog —ежик). Некоторые гены называются именами богов (например, гефестин – от греческого бога огня и кузнечного дела Гефеста). На практике генетики используют сокращенные обозначения генов, состоящие обычно из 3—5 латинских букв. Поскольку функции многих генов еще не известны, а удается только определить, что какая–то конкретная область генома человека способна теоретически кодировать белок, то часто пока еще перед названиями генов появляется слово «гипотетический».
Анализ генома человека позволил на конец 2002 года обнаружить порядка 30 000—40 000 генов, кодирующих белки. Эти гены сильно отличаются друг от друга по размерам. Подсчитано, что средняя длина гена у человека составляет около 27 тыс. п. н. Такой усредненный ген содержит 9 экзонов (средний размер каждого около 150 п. н.) и 8 интронов (средний размер каждого чуть меньше 3400 п.н.). Но это лишь средние значения. Самые короткие гены содержат всего два десятка букв–нуклеотидов, например, гены эндорфинов – полипептидов, вызывающих ощущение удовольствия. В то же время самый длинный ген, кодирующий один из белков мышц, – миодистрофин содержит 2,4 млн. п. н.
Кроме того, гены не равномерно распределены между разными хромосомами. При средней плотности один ген на 100 т. п. н. генома их содержание в хромосоме 19 составляет около 2 на 100 т. п.н., а в хромосомах 2, 13 и Y – хромосоме – от 0,15 до 0,7. Если сравнить плотность генов с плотностью расселения людей, то Y – хромосома напоминает нашу Сибирь, а хромосома 19 – Европейскую часть России. Для сравнения: в геноме бактерий содержится свыше 1000 генов на 1,0 млн. п. н., у дрожжей около 450 генов на 1,0 млн. п. н., а у червя C. elegans – около 200. Следовательно, плотность расположения генов на единицу длины генома заметно падает по мере эволюционного усложнения организмов.
«Жи, ши – пиши через и»
После секвенирования генома человека и ряда других организмов был проведен детальный компьютерный анализ нуклеотидных последовательностей. Подсчитано, например, сколько в ДНК может идти подряд букв А, или как часто Г встречается после Ц. И тут выяснилось, что в генах эти сочетания подчиняются определенным правилам, тогда как в промежутках между генами, там, где ничего существенного в ДНК не записано, частота сочетаний разных нуклеоти–дов близка к случайной. Где много генов, там много букв Г и Ц, а где генов нет, там много букв А и Т. Здесь опять можно провести параллель с обычным текстом. По правилам грамматики русского языка, которые мы учили в школе, «жи, ши – пиши через и», в словах после букв ж и ш буква ы не встречается. Нечто подобное характерно и для генетических текстов – в кодирующих участках некоторые сочетания нуклеотидов практически не встречаются, а распределение других сочетаний сильно отличается от случайного. Это был один из первых, но далеко не единственный результат анализа ДНК.
Знаки препинания (Генетическая пунктуация)
Как в письменном языке пунктуация (точки, запятые, тире и т.д.) используется для членения и графической организации текста, так и в ДНКовом тексте существуют всевозможные знаки–сигналы, обозначающие границы отдельных функциональных элементов. Без таких сигналов генетическая информация не может быть правильно считана с носителя. Классический пример из лингвистики – предложение «Убить нельзя помиловать». Если не поставить запятую, то остается непонятным, то ли убить, то ли помиловать. Так и в гене. Знаки препинания, в отличие от генетического кода, не так универсальны в живом мире. Часть из них у низших организмов (прокариот) существенно отличаются по сравнению с высшими (эукариота–ми), включая человека.
Вот как выглядит, например, одна из страниц ДНКового текста, содержащаяся в томе (хромосоме) 11 человека (рис. 17).
Глядя на этот рисунок, даже специалисту сразу трудно что–то понять. Однако простая компьютерная программа быстро позволяет найти здесь участок, кодирующий белок интерферон. Это осуществляется в результате того, что в генах имеются специальные сигналы–ярлычки, которые можно рассматривать как знаки препинания в ДНКовом тексте. И компьютер способен их различить. Рассмотрим главные их них.
Так, в генах человека, как и у других высших организмов, имеются специальные последовательности–сигналы, которые определяют начало и конец процесса транскрипции, то есть синтеза мРНК.
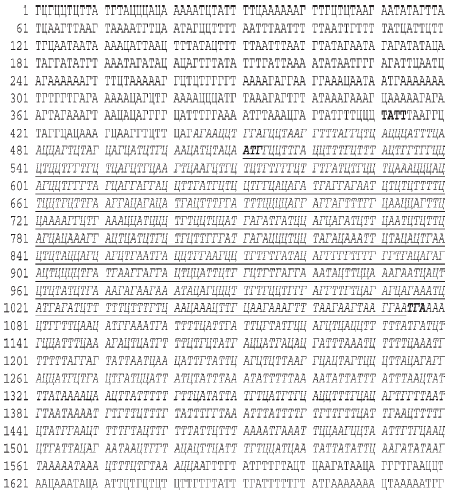
Рис. 17. Страница Энциклопедии человека, на которой записана последовательность нуклеотидов ДНК человека, кодирующая белок альфа–интерферон. Ген интерферона выделен курсивом. Подчеркнут участок, кодирующий белок, состоящий из 188 аминокислот. Жирным шрифтом выделены знаки пунктуации в этом гене, обеспечивающие его правильную транскрипцию с образованием мРНК и последующую правильную трансляцию этой мРНК
Фермент, называемый РНК–полимеразой, узнает эти сигналы и осуществляет синтез «правильных» молекул мРНК (со строго запрограммированными в геноме концами). Одним из таких знаков препинания в ДНКовом тексте, отмечающим начало процесса транскрипции, служит короткая последовательность нуклеотидов: ТАТА (как пример, см. рис. 17).
Для процесса трансляции, то есть синтеза белка на мРНК, также имеются свои специальные знаки препинания. Таковыми являются отдельные тройки нуклеотидов (триплеты). На рис. 17 они выделены жирным шрифтом. Место, с которого начинается кодирование белка, – старт–сигнал – триплет АУТ. Далее следуют триплеты, которые кодируют аминокислоты в белке. А в конце текста имеется специальный стоп–сигнал (триплет УГА).
Но это еще не все. ДНК иногда претерпевает некоторые химические изменения. Известна такая химическая модификация ДНК, как метилирование, которая происходит строго по динуклеотидам–сигналам, представляющим собой пару нуклеотидов, в которой первым идет Г, а за ним Ц. Наконец, важными сигналами в генах являются участки, определяющие такой важный процесс, как сплайсинг, который происходит уже после образования РНК на гене. Об этом и пойдет речь дальше.
Вот как выглядит, например, одна из страниц ДНКового текста, содержащаяся в томе (хромосоме) 11 человека (рис. 17).
Глядя на этот рисунок, даже специалисту сразу трудно что–то понять. Однако простая компьютерная программа быстро позволяет найти здесь участок, кодирующий белок интерферон. Это осуществляется в результате того, что в генах имеются специальные сигналы–ярлычки, которые можно рассматривать как знаки препинания в ДНКовом тексте. И компьютер способен их различить. Рассмотрим главные их них.
Так, в генах человека, как и у других высших организмов, имеются специальные последовательности–сигналы, которые определяют начало и конец процесса транскрипции, то есть синтеза мРНК.
Рис. 17. Страница Энциклопедии человека, на которой записана последовательность нуклеотидов ДНК человека, кодирующая белок альфа–интерферон. Ген интерферона выделен курсивом. Подчеркнут участок, кодирующий белок, состоящий из 188 аминокислот. Жирным шрифтом выделены знаки пунктуации в этом гене, обеспечивающие его правильную транскрипцию с образованием мРНК и последующую правильную трансляцию этой мРНК
Фермент, называемый РНК–полимеразой, узнает эти сигналы и осуществляет синтез «правильных» молекул мРНК (со строго запрограммированными в геноме концами). Одним из таких знаков препинания в ДНКовом тексте, отмечающим начало процесса транскрипции, служит короткая последовательность нуклеотидов: ТАТА (как пример, см. рис. 17).
Для процесса трансляции, то есть синтеза белка на мРНК, также имеются свои специальные знаки препинания. Таковыми являются отдельные тройки нуклеотидов (триплеты). На рис. 17 они выделены жирным шрифтом. Место, с которого начинается кодирование белка, – старт–сигнал – триплет АУТ. Далее следуют триплеты, которые кодируют аминокислоты в белке. А в конце текста имеется специальный стоп–сигнал (триплет УГА).
Но это еще не все. ДНК иногда претерпевает некоторые химические изменения. Известна такая химическая модификация ДНК, как метилирование, которая происходит строго по динуклеотидам–сигналам, представляющим собой пару нуклеотидов, в которой первым идет Г, а за ним Ц. Наконец, важными сигналами в генах являются участки, определяющие такой важный процесс, как сплайсинг, который происходит уже после образования РНК на гене. Об этом и пойдет речь дальше.
Гены построены из кусков
Успехи в изучении генома человека, а еще ранее – проведенный анализ отдельных генов других высших организмов, привели к ряду принципиально новых и совершенно неожиданных для генетиков результатов в отношении устройства генов, особенно тех, которые кодируют белки. Со времени появления центральной догмы молекулярной биологии думали, что непрерывная линейная последовательность нуклеотидов в ДНК с помощью триплетов нуклеоти–дов кодирует непрерывную последовательность нуклеотидов в мРНК, а последняя как посредник передает эту информацию на рибосомы, которые, считывая ее, осуществляют синтез линейных молекул белков. Это соответствовало ситуации, которую первоначально наблюдали при изучении генов у бактерий. Однако у высших организмов, включая человека, (и даже у некоторых низших) картина оказалась более сложной.
Когда сравнивали мРНК и соответствующий белок, то действительно обнаруживали непрерывность их нуклеотидных и аминокислотных последовательностей. То же наблюдали и при сопоставлении последовательностей аминокислот в белках и нуклеотидов в геномах вирусов. Однако с появлением возможности секвенировать протяженные участки ДНК (т. е. непосредственно генов) как у некоторых вирусов, так и у высших организмов получили совершенно неожиданные результаты. Внутри нуклеотидных последовательностей ДНК, кодирующих белок, были обнаружены какие–то иные, ничего не кодирующие участки. Чудеса, да и только! В чем же дело? Оказалось, что многие гены «расчленены» на отдельные куски. Одни из этих кусков, как и положено, кодируют белок (их назвали экзонами), а другие никакие белки кодировать не способны (их назвали интронами) и расположены между экзонами. Таким образом, в гене кодирующие и не кодирующие участки перемежаются друг с другом (рис. 18). Так устроено большинство белок–кодирующих генов. Хотя имеются и исключения. В этих случаях говорят, что ген устроен из одного экзона и интронов не содержит.
Такое разорванное (экзон–интронное) строение оказалось характерным для подавляющего большинства (хотя и не для всех) генов человека. Вместе с тем у большинства простейших организмов инт
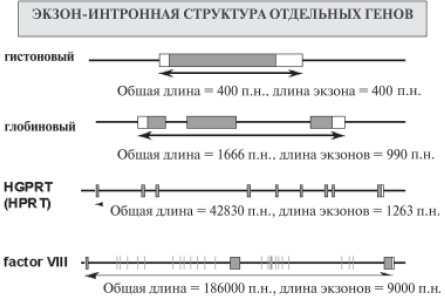
Рис. 18. Схематическое изображение экзон–интронного строения некоторых генов человека. Экзоны – прямоугольнники, интроны – линии между прямоугольниками. Темные прямоугольники – участки генов, кодирующие белок, белые прямоугольники в первом и последнем экзонах – специальные участки гена, кодирующие мРНК, но не кодирующие белок
роны не обнаружены. Размеры интронов часто значительно превышают размеры экзонов, что существенно увеличивает общую длину нуклеотидных последовательностей ДНК, образующих ген. При этом в геноме человека интроны в целом существенно длиннее, чем у других организмов.
Каким же образом разорванный ДНКовый текст реализуется в клетках в виде неразорванного РНКового, а затем белкового? Выяснилось, что первоначально при транскрипции гена синтезируется большой РНК–предшественник, копия ДНКового текста (с экзона–ми и интронами вместе). Далее в результате работы специфических ферментов происходит нарезание РНК–предшественника на куски. Те из них, которые ничего не кодируют (интроны), отбрасываются, а кодирующие куски (экзоны) соединяются между собой, обычно в том же порядке, как они располагались исходно в ДНК. В результате формируется зрелая молекула мРНК. Этот сложный процесс и получил название сплайсинга (рис. 19).
Когда сравнивали мРНК и соответствующий белок, то действительно обнаруживали непрерывность их нуклеотидных и аминокислотных последовательностей. То же наблюдали и при сопоставлении последовательностей аминокислот в белках и нуклеотидов в геномах вирусов. Однако с появлением возможности секвенировать протяженные участки ДНК (т. е. непосредственно генов) как у некоторых вирусов, так и у высших организмов получили совершенно неожиданные результаты. Внутри нуклеотидных последовательностей ДНК, кодирующих белок, были обнаружены какие–то иные, ничего не кодирующие участки. Чудеса, да и только! В чем же дело? Оказалось, что многие гены «расчленены» на отдельные куски. Одни из этих кусков, как и положено, кодируют белок (их назвали экзонами), а другие никакие белки кодировать не способны (их назвали интронами) и расположены между экзонами. Таким образом, в гене кодирующие и не кодирующие участки перемежаются друг с другом (рис. 18). Так устроено большинство белок–кодирующих генов. Хотя имеются и исключения. В этих случаях говорят, что ген устроен из одного экзона и интронов не содержит.
Такое разорванное (экзон–интронное) строение оказалось характерным для подавляющего большинства (хотя и не для всех) генов человека. Вместе с тем у большинства простейших организмов инт
Рис. 18. Схематическое изображение экзон–интронного строения некоторых генов человека. Экзоны – прямоугольнники, интроны – линии между прямоугольниками. Темные прямоугольники – участки генов, кодирующие белок, белые прямоугольники в первом и последнем экзонах – специальные участки гена, кодирующие мРНК, но не кодирующие белок
роны не обнаружены. Размеры интронов часто значительно превышают размеры экзонов, что существенно увеличивает общую длину нуклеотидных последовательностей ДНК, образующих ген. При этом в геноме человека интроны в целом существенно длиннее, чем у других организмов.
Каким же образом разорванный ДНКовый текст реализуется в клетках в виде неразорванного РНКового, а затем белкового? Выяснилось, что первоначально при транскрипции гена синтезируется большой РНК–предшественник, копия ДНКового текста (с экзона–ми и интронами вместе). Далее в результате работы специфических ферментов происходит нарезание РНК–предшественника на куски. Те из них, которые ничего не кодируют (интроны), отбрасываются, а кодирующие куски (экзоны) соединяются между собой, обычно в том же порядке, как они располагались исходно в ДНК. В результате формируется зрелая молекула мРНК. Этот сложный процесс и получил название сплайсинга (рис. 19).