Страница:
Глава 6
Неотения
В биологическом смысле организм становится половозрелым, или взрослым, когда делается способным к размножению. И он становится физически зрелым, когда приобретает взрослые формы. Скорости, с которыми организм созревает физически и сексуально, независимо контролируются разными генами[71]. Популяция способна эволюционировать так, что индивиды созревают физически раньше или позже, сохраняя неизменной скорость полового созревания, или индивиды созревают сексуально быстрее или медленнее, сохраняя постоянной скорость физического созревания, или же могут меняться обе скорости.
Существует два способа, с помощью которых популяция может эволюционировать таким образом, что индивиды сохраняют детские черты во взрослом состоянии (педоморфизм). Популяция может эволюционировать в сторону ускорения физического и полового созревания так, что индивиды становятся физически и сексуально зрелыми, когда они все еще детеныши (прогенез, например, у тритонов). Во втором случае популяция эволюционирует в сторону замедления или прекращения физического созревания, но возраст полового созревания остается приблизительно таким же, и индивиды, достигая половой зрелости, выглядят по-детски (неотения). Неотению (от греч. nйos – незрелый, юный и tйino – растягиваю, удлиняю) относят на счет генного контроля, изменяющего процесс созревания индивидов. Неотения имеет место, когда индивиды созревают в половом отношении примерно с нормальной скоростью, но хотя их тело и увеличивается в размерах по мере полового созревания, их ювенильные черты (и ювенильные черты их предков) сохраняются во взрослом состоянии и не меняются на отчетливо отличные от них взрослые черты. Другими словами, ребенок становится более крупным, но сексуально зрелым ребенком.

Эволюция человека происходила путем целого ряда генетических изменений, но одним из наиболее важных явилась неотения. Человек является наиболее неотеничным из всех приматов. Обратите внимание на рисунок 6–1[72], где дается поразительное и важное сравнение взрослого шимпанзе и его детеныша. У детеныша шимпанзе более сходная с человеческим лицом намного более плоская морда, тогда как у взрослого шимпанзе нижняя челюсть очень сильно выступает вперед[73]. Поскольку у взрослого шимпанзе не сохраняются детские формы морды, шимпанзе не является неотеничным. Теперь давайте представим, что детеныш шимпанзе вырастет и станет половозрелым, но его морда не изменится. Тогда шимпанзе станет неотеничным и будет гораздо сильнее походить на человека.
Теперь, когда вы знаете, что такое неотения, нетрудно понять, что человек неотеничен. В действительности человек неотеничен настолько, что его даже описывают как «половозрелый плод»[74]. Многие наши неотеничные признаки жизненно важны для нашей эволюции. У большинства плодов млекопитающих, включая человека, большое затылочное отверстие (foramen occipitale magnum); через которое спинной мозг проникает в череп, смещено к центру основания черепа. У четвероногих животных оно по мере взросления перемещается назад (Таблица 9–2), но у человека, передвигающегося на двух ногах, оно остается в младенческом положении (так, что глаза направлены перпендикулярно оси позвоночника). У эмбрионов млекопитающих вход во влагалище направлен вперед, а у взрослых женщин таким и остается (обеспечивая возможность соития лицом к лицу) и не перемещается назад (давая возможность соития в заднем положении) как у других млекопитающих. Наш большой палец на ногах остается параллельным другим пальцам (для ходьбы) и не отклоняется на 90° по отношению к ним (для захватывания) как у крупных человекообразных обезьян. К неотеничным признакам человека относится также более грацильный (т. е. менее мощный) скелет, более крупный (по отношению к размеру тела) и округлый, состоящий из более тонких костей череп, более плоское лицо с менее выступающей вперед нижней челюстью («прогнатизм»[75]), меньшие по размеру зубы, меньшая оволосенность тела, более короткие руки, ноги, пальцы и ступни, а кроме того, больший слой подкожного жира – все это признаки, свойственные детенышам приматов.
Телесный цвет кожи человека тоже может быть неотеничным признаком. Новорожденные у темнокожих родителей имеют более светлую кожу, темнеющую по мере взросления (Abner, 1998). Интересно отметить, что молодые шимпанзе имеют кожу телесного цвета, темнеющую или чернеющую на десятом – двенадцатом годах жизни (Baker, 1974, с. 112). Это предполагает, что наш последний общий предок с шимпанзе также мог иметь светлую кожу в юном возрасте (Jablonski, 2006). Имеются некоторые генетические свидетельства того, что «общие предки всех людей на земле имели белую кожу под темной шерстью – подобную по окраске коже и шерсти современных [молодых] шимпанзе»[76] (Rogers, 2004). Волосы у новорожденных также прямее, даже у негроидных младенцев, и плод имеет складку эпикантуса (жировая складка над верхним веком, частично прикрывающая и защищающая глаза, см. рис. 10–3 на стр. 141), так что эти признаки также являются неотеничными. Белая склера (глазного яблока) также может быть неотеничным признаком, поскольку: «У большинства млекопитающих склера темнеет с возрастом, [но] у человека склера остается белой в течение всей жизни» (Etcoff, 1999, с. 33).
Что послужило причиной человеческой неотении? Очевидный ответ состоит в том, что прежде, чем человек сделался неотеничным, индивиды слегка различались по степени своей неотеничности, как они различаются практически по всем признакам. Человек никогда бы не стал неотеничным, если бы не изменилась среда обитания. После ее изменения более неотеничные индивиды были репродуктивно успешнее индивидов, не имеющих аллелей, ответственных за неотению, и вся популяция стала более неотеничной.
Суть другого вопроса заключается в следующем: «Какие изменения среды могли сделать неотеничные признаки более благоприятными?» Меньшая по размеру и не выступающая вперед челюсть и меньшая массивность фигуры должны быть серьезным недостатком в схватке. Но если человек уже достаточно преуспел в разработке орудий труда и оружия, то эти признаки становятся необязательными, являясь расточительным использованием ресурсов организма и энергии, и снижают скорость и маневренность. Какие еще присущие младенцам признаки, обладай ими взрослые, могли бы увеличить шансы взрослых на выживание?
Другой возможностью является более крупный мозг. В отношении к размеру тела мозг младенцев крупнее, чем у взрослых[77], и у более неотеничных взрослых мозг обычно крупнее, чем у менее неотеничных. Верно также и то, что существует умеренная корреляция[78] между уровнем интеллекта и размером мозга (r = 0,44, Lynn, 2006a, с. 214; см. также Witelson, 2006; McDaniel, 2005). Это не очень высокая корреляция – люди с большим мозгом могут все же быть глупыми, – но это все-таки значимая корреляция. Так что изменения среды обитания, вероятно, потребовали более высокого интеллекта для выживания, поэтому более неотеничные и, следовательно, имевшие более крупный мозг и более высокий интеллект индивиды должны были подвергаться положительному отбору. Если популяция мигрирует из тропиков, где сезонные изменения невелики, на Север, где существуют четыре отчетливо различающиеся сезона, включая долгую холодную зиму, более высокий интеллект становится важным преимуществом в планировании своих действий и добыче продовольствия зимой. Таким образом, сезонные изменения климата будут фактором отбора в сторону увеличения интеллекта и, следовательно, в сторону возрастания неотеничности индивидов.
Трудно оценить, насколько суровым должен был быть отбор по интеллекту. Небольшой мозг все же способен обеспечить пропитание зимой – белки способны к этому на протяжении всего года, обладая намного меньшим мозгом (в отношении к размеру тела), чем человек. Более того, мозг является наиболее затратным органом, так как требует больше энергии (на единицу массы), чем любой другой орган. Мозг взрослых людей, составляя 2 % (Leakey, 1994, с. 54) или 3 % (Foley, 1995, с. 170) от массы тела, использует тем не менее 20 % потребляемой организмом энергии[79], а у новорожденных, поразительно, на долю мозга приходится в среднем 75 % суточного потребления энергии. Более крупный мозг способен помогать решать больше проблем, но это дополнительный вес, который необходимо переносить, и дополнительное питание для поддержания его функционирования. Чтобы увидеть, как сместились выгоды и ущербы, необходимо рассмотреть, насколько в действительности возрос интеллект на Севере, что мы проанализируем в этой книге позднее.
Младенцы почти везде, за исключением, возможно, тропиков, должны сохранять тепло для предотвращения смерти от гипотермии. Вследствие своего малого размера (высокого отношения поверхности к массе тела) им необходимо сохранять тепло и минимизировать сжигание калорий. Они должны обладать многими признаками, помогающими им делать это. Эти признаки были бы полезны и для мигрировавших на Север взрослых, одним из таких признаков является младенческий жир. Младенцы обладают дополнительным подкожным жиром, равномерно распределенным по телу, который сохраняет энергию для их быстро растущего мозга, обеспечивает некоторую защиту от ударов и сохраняет тепло. Другие неотеничные признаки, полезные в холодном климате, включают складку эпикантуса и признаки, уменьшающие поверхность тела, например, уплощенное лицо, короткие руки и ноги (Baker, 1974, с. 307) и широкое тело. Все эти признаки характерны для популяций на севере Азии. Все это подсказывает, что мигрировавшие в зоны более холодного климата популяции могли подвергаться жесткому отбору в сторону неотении.
Наиболее неотеничными на нашей планете являются аборигены Восточной Азии, а среди них корейцы, обладающие наибольшим слоем подкожного жира, за ними следуют ханьцы Китая и другие монголоиды. У аборигенов Восточной Азии, как и у младенцев, круглая голова с плоским пухлым лицом, маленький нос, короткие руки и ноги, очень мало волос на теле и избыток подкожного жира, равномерно распределенного по всей поверхности тела. Их «третье веко» (складка эпикантуса) и маленькие глазничные впадины помогают им предохранять глаза от холода. Очевидно, что эти народы эволюционировали в холодном климате, а поскольку они сделались столь неотеничными, это предполагает, что неотения была полезной в таком климате (Глава 4, Правило 11).
Европейцы также стали неотеничными, но в значительно меньшей степени, чем аборигены Азии. У европейцев длиннее голова, больше волос на теле, длиннее конечности и слой подкожного жира меньше и распределен не так равномерно; вместо этого он накапливается малосимпатичными массами вокруг живота, бедер и ляжек, обеспечивая хороший источник доходов индустрии снижения веса. Большинство африканцев по-прежнему менее неотеничны, но их родословная сложнее, что делает разные африканские популяции сильно различающимися по самым разнообразным признакам (Глава 26).
Существует два способа, с помощью которых популяция может эволюционировать таким образом, что индивиды сохраняют детские черты во взрослом состоянии (педоморфизм). Популяция может эволюционировать в сторону ускорения физического и полового созревания так, что индивиды становятся физически и сексуально зрелыми, когда они все еще детеныши (прогенез, например, у тритонов). Во втором случае популяция эволюционирует в сторону замедления или прекращения физического созревания, но возраст полового созревания остается приблизительно таким же, и индивиды, достигая половой зрелости, выглядят по-детски (неотения). Неотению (от греч. nйos – незрелый, юный и tйino – растягиваю, удлиняю) относят на счет генного контроля, изменяющего процесс созревания индивидов. Неотения имеет место, когда индивиды созревают в половом отношении примерно с нормальной скоростью, но хотя их тело и увеличивается в размерах по мере полового созревания, их ювенильные черты (и ювенильные черты их предков) сохраняются во взрослом состоянии и не меняются на отчетливо отличные от них взрослые черты. Другими словами, ребенок становится более крупным, но сексуально зрелым ребенком.
Эволюция человека происходила путем целого ряда генетических изменений, но одним из наиболее важных явилась неотения. Человек является наиболее неотеничным из всех приматов. Обратите внимание на рисунок 6–1[72], где дается поразительное и важное сравнение взрослого шимпанзе и его детеныша. У детеныша шимпанзе более сходная с человеческим лицом намного более плоская морда, тогда как у взрослого шимпанзе нижняя челюсть очень сильно выступает вперед[73]. Поскольку у взрослого шимпанзе не сохраняются детские формы морды, шимпанзе не является неотеничным. Теперь давайте представим, что детеныш шимпанзе вырастет и станет половозрелым, но его морда не изменится. Тогда шимпанзе станет неотеничным и будет гораздо сильнее походить на человека.
Теперь, когда вы знаете, что такое неотения, нетрудно понять, что человек неотеничен. В действительности человек неотеничен настолько, что его даже описывают как «половозрелый плод»[74]. Многие наши неотеничные признаки жизненно важны для нашей эволюции. У большинства плодов млекопитающих, включая человека, большое затылочное отверстие (foramen occipitale magnum); через которое спинной мозг проникает в череп, смещено к центру основания черепа. У четвероногих животных оно по мере взросления перемещается назад (Таблица 9–2), но у человека, передвигающегося на двух ногах, оно остается в младенческом положении (так, что глаза направлены перпендикулярно оси позвоночника). У эмбрионов млекопитающих вход во влагалище направлен вперед, а у взрослых женщин таким и остается (обеспечивая возможность соития лицом к лицу) и не перемещается назад (давая возможность соития в заднем положении) как у других млекопитающих. Наш большой палец на ногах остается параллельным другим пальцам (для ходьбы) и не отклоняется на 90° по отношению к ним (для захватывания) как у крупных человекообразных обезьян. К неотеничным признакам человека относится также более грацильный (т. е. менее мощный) скелет, более крупный (по отношению к размеру тела) и округлый, состоящий из более тонких костей череп, более плоское лицо с менее выступающей вперед нижней челюстью («прогнатизм»[75]), меньшие по размеру зубы, меньшая оволосенность тела, более короткие руки, ноги, пальцы и ступни, а кроме того, больший слой подкожного жира – все это признаки, свойственные детенышам приматов.
Телесный цвет кожи человека тоже может быть неотеничным признаком. Новорожденные у темнокожих родителей имеют более светлую кожу, темнеющую по мере взросления (Abner, 1998). Интересно отметить, что молодые шимпанзе имеют кожу телесного цвета, темнеющую или чернеющую на десятом – двенадцатом годах жизни (Baker, 1974, с. 112). Это предполагает, что наш последний общий предок с шимпанзе также мог иметь светлую кожу в юном возрасте (Jablonski, 2006). Имеются некоторые генетические свидетельства того, что «общие предки всех людей на земле имели белую кожу под темной шерстью – подобную по окраске коже и шерсти современных [молодых] шимпанзе»[76] (Rogers, 2004). Волосы у новорожденных также прямее, даже у негроидных младенцев, и плод имеет складку эпикантуса (жировая складка над верхним веком, частично прикрывающая и защищающая глаза, см. рис. 10–3 на стр. 141), так что эти признаки также являются неотеничными. Белая склера (глазного яблока) также может быть неотеничным признаком, поскольку: «У большинства млекопитающих склера темнеет с возрастом, [но] у человека склера остается белой в течение всей жизни» (Etcoff, 1999, с. 33).
Что послужило причиной человеческой неотении? Очевидный ответ состоит в том, что прежде, чем человек сделался неотеничным, индивиды слегка различались по степени своей неотеничности, как они различаются практически по всем признакам. Человек никогда бы не стал неотеничным, если бы не изменилась среда обитания. После ее изменения более неотеничные индивиды были репродуктивно успешнее индивидов, не имеющих аллелей, ответственных за неотению, и вся популяция стала более неотеничной.
Суть другого вопроса заключается в следующем: «Какие изменения среды могли сделать неотеничные признаки более благоприятными?» Меньшая по размеру и не выступающая вперед челюсть и меньшая массивность фигуры должны быть серьезным недостатком в схватке. Но если человек уже достаточно преуспел в разработке орудий труда и оружия, то эти признаки становятся необязательными, являясь расточительным использованием ресурсов организма и энергии, и снижают скорость и маневренность. Какие еще присущие младенцам признаки, обладай ими взрослые, могли бы увеличить шансы взрослых на выживание?
Другой возможностью является более крупный мозг. В отношении к размеру тела мозг младенцев крупнее, чем у взрослых[77], и у более неотеничных взрослых мозг обычно крупнее, чем у менее неотеничных. Верно также и то, что существует умеренная корреляция[78] между уровнем интеллекта и размером мозга (r = 0,44, Lynn, 2006a, с. 214; см. также Witelson, 2006; McDaniel, 2005). Это не очень высокая корреляция – люди с большим мозгом могут все же быть глупыми, – но это все-таки значимая корреляция. Так что изменения среды обитания, вероятно, потребовали более высокого интеллекта для выживания, поэтому более неотеничные и, следовательно, имевшие более крупный мозг и более высокий интеллект индивиды должны были подвергаться положительному отбору. Если популяция мигрирует из тропиков, где сезонные изменения невелики, на Север, где существуют четыре отчетливо различающиеся сезона, включая долгую холодную зиму, более высокий интеллект становится важным преимуществом в планировании своих действий и добыче продовольствия зимой. Таким образом, сезонные изменения климата будут фактором отбора в сторону увеличения интеллекта и, следовательно, в сторону возрастания неотеничности индивидов.
Трудно оценить, насколько суровым должен был быть отбор по интеллекту. Небольшой мозг все же способен обеспечить пропитание зимой – белки способны к этому на протяжении всего года, обладая намного меньшим мозгом (в отношении к размеру тела), чем человек. Более того, мозг является наиболее затратным органом, так как требует больше энергии (на единицу массы), чем любой другой орган. Мозг взрослых людей, составляя 2 % (Leakey, 1994, с. 54) или 3 % (Foley, 1995, с. 170) от массы тела, использует тем не менее 20 % потребляемой организмом энергии[79], а у новорожденных, поразительно, на долю мозга приходится в среднем 75 % суточного потребления энергии. Более крупный мозг способен помогать решать больше проблем, но это дополнительный вес, который необходимо переносить, и дополнительное питание для поддержания его функционирования. Чтобы увидеть, как сместились выгоды и ущербы, необходимо рассмотреть, насколько в действительности возрос интеллект на Севере, что мы проанализируем в этой книге позднее.
Младенцы почти везде, за исключением, возможно, тропиков, должны сохранять тепло для предотвращения смерти от гипотермии. Вследствие своего малого размера (высокого отношения поверхности к массе тела) им необходимо сохранять тепло и минимизировать сжигание калорий. Они должны обладать многими признаками, помогающими им делать это. Эти признаки были бы полезны и для мигрировавших на Север взрослых, одним из таких признаков является младенческий жир. Младенцы обладают дополнительным подкожным жиром, равномерно распределенным по телу, который сохраняет энергию для их быстро растущего мозга, обеспечивает некоторую защиту от ударов и сохраняет тепло. Другие неотеничные признаки, полезные в холодном климате, включают складку эпикантуса и признаки, уменьшающие поверхность тела, например, уплощенное лицо, короткие руки и ноги (Baker, 1974, с. 307) и широкое тело. Все эти признаки характерны для популяций на севере Азии. Все это подсказывает, что мигрировавшие в зоны более холодного климата популяции могли подвергаться жесткому отбору в сторону неотении.
Наиболее неотеничными на нашей планете являются аборигены Восточной Азии, а среди них корейцы, обладающие наибольшим слоем подкожного жира, за ними следуют ханьцы Китая и другие монголоиды. У аборигенов Восточной Азии, как и у младенцев, круглая голова с плоским пухлым лицом, маленький нос, короткие руки и ноги, очень мало волос на теле и избыток подкожного жира, равномерно распределенного по всей поверхности тела. Их «третье веко» (складка эпикантуса) и маленькие глазничные впадины помогают им предохранять глаза от холода. Очевидно, что эти народы эволюционировали в холодном климате, а поскольку они сделались столь неотеничными, это предполагает, что неотения была полезной в таком климате (Глава 4, Правило 11).
Европейцы также стали неотеничными, но в значительно меньшей степени, чем аборигены Азии. У европейцев длиннее голова, больше волос на теле, длиннее конечности и слой подкожного жира меньше и распределен не так равномерно; вместо этого он накапливается малосимпатичными массами вокруг живота, бедер и ляжек, обеспечивая хороший источник доходов индустрии снижения веса. Большинство африканцев по-прежнему менее неотеничны, но их родословная сложнее, что делает разные африканские популяции сильно различающимися по самым разнообразным признакам (Глава 26).
Глава 7
Генетическое расстояние
Популяции, находящиеся в репродуктивной изоляции (обычно вследствие географической изолированности), постепенно приобретают генетические особенности. Главной причиной возникновения особенностей служат различающиеся селекторы разных сред обитания (или давление отбора, обусловленное этими селекторами). Иногда часть популяции перемещается на другую территорию или становится изолированной от оставшейся популяции из-за подъема уровня вод, смещения русла рек, оледенений, образования пустынь или по другим причинам. Тогда, при условии, что некоторые индивиды этой изолированной популяции оказались немного генетически отличными от остающихся в исходной популяции, вероятнее всего, вся изолированная популяция станет еще более генетически отличной от родительской. Это называется «эффектом основателя». Случайные мутации также могут возникать в одной популяции и не возникать в другой, либо только одна из популяций имеет возможность интербридинга с третьей популяцией.
Генетическое расстояние – это способ численно выразить, насколько генетически различны два индивида или две популяции. Как объясняется в Главе 3, все люди имеют одни и те же гены, например, все мы имеем ген, определяющий цвет глаз, но каждый ген в среднем имеет 14 различных последовательностей нуклеотидов, называемых «аллелями». Для определения генетического расстояния между двумя индивидами можно подсчитать число аллелей, которые у них различаются. В случае популяций подсчитывается число индивидов в каждой из них, имеющих определенные аллели (предпочтительно использовать большое число аллелей для повышения точности), и результаты выражаются математически. Если индивид имеет близнеца, его аллели будут идентичны аллелям его близнеца и генетическое расстояние между ними будет равно нулю. Если другой индивид является вашим ребенком, одинаковой будет, по крайней мере, половина аллелей. (Если его другой родитель имеет некоторые аллели, общие с вашими, у ребенка будет больше половины ваших аллелей.) Если брак является кровосмесительным, число общих с родительскими аллелей у детей будет выше, чем когда родители не состоят в родстве. Число общих аллелей в целом ниже у двоюродных братьев и сестер, еще ниже у не связанных родством представителей одного этноса или расы, и еще ниже у представителей разных рас и разных биологических видов, и оно продолжает снижаться по мере удаления от последнего общего предка (LCA) человека и других видов.
Если мы построим график генетического расстояния от вас (предположив, что вы европеоид) до других людей на планете, он может выглядеть аналогично изображенному на рисунке 7–1. На этом рисунке весьма приближенно показано, как быстро увеличивается генетическое расстояние при удалении от ближайших родственников. Затем имеет место существенное увеличение генетического расстояния между вами и монголоидами и еще намного большее его увеличение между вами и африканцами[80].

Пока что невозможно полностью проанализировать ДНК каждого человека на планете[81] и сравнить его ДНК с ДНК любого другого человека для определения того, сколько у них общих аллелей, но существуют упрощенные методики, дающие примерно одинаковые результаты. Генетическое расстояние между отдельными людьми и популяциями может быть рассчитано на основании анализа образцов ДНК (Salter, 2003). Собирая образцы ДНК в разных регионах всего мира и рассчитывая число однонуклеотидных полиморфизмов, ученые определяют генетические расстояния между разными популяциями, этносами, расами и биологическими видами. Числа в верхней части рисунка 7–2 (Cavalli-Sforza, 1991) показывают в процентах генетические расстояния (умноженные на 10 000) между разными человеческими популяциями, вычисленные с помощью модифицированного метода Нея.
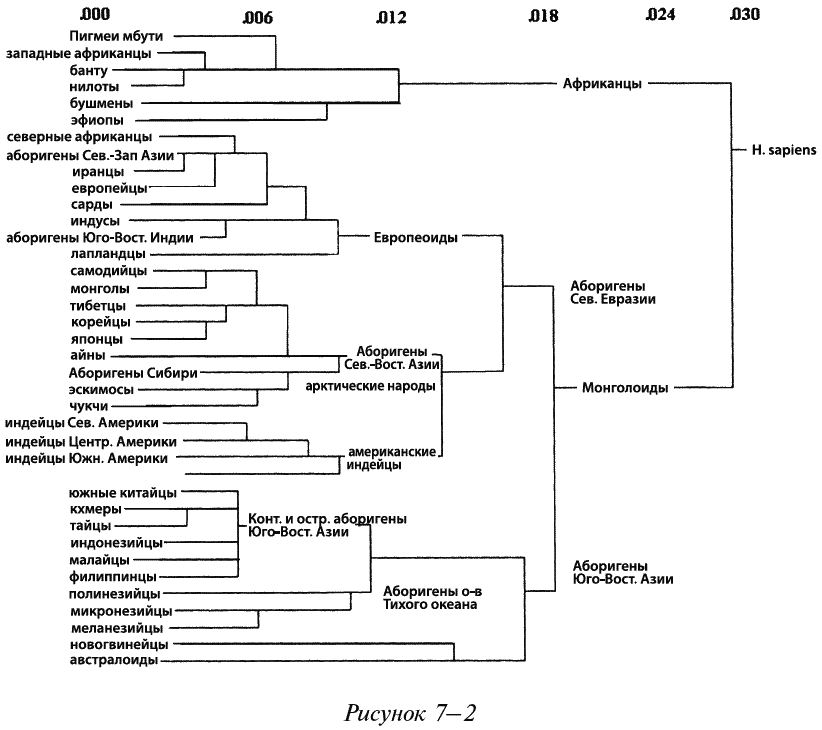
Что касается трех основных рас, то на рисунке 7–2 показано, что африканцы из тропической Африки и из любых других регионов состоят с остальными людьми в наименьшем родстве, а аборигены Севера Евразии и Юго-Восточной Азии занимают второе место по генетическому расстоянию между собой. Обратите внимание, что к европеоидам отнесены аборигены Северной Африки (обитающие вокруг Средиземного моря), Северо-Западной Азии (Ближнего Востока) и индусы. Также заметьте, что аборигены Северо-Восточной Азии и американские индейцы состоят в более тесном родстве с европеоидами, чем с аборигенами Юго-Восточной Азии.
На рисунке 7–3 отмечены положения 42 популяций человечества относительно двух осей, показывающих различия по двум высоко вариабельным участкам мтДНК (Cavalli-Sforza, 1994, с. 82). Оси первой и второй главных компонент делят данные на две половины, обуславливающие наибольшую и вторую по величине дисперсии, соответственно (см. Википедия, Метод главных компонент). Африканцы находятся на одном конце обеих осей, а все остальные на другом, потому что африканцы отличаются генетически от большинства других популяций. Так как некоторые популяции (аборигены Евразии) подверглись более значительной эволюции, чем остальные (африканцы), то точка, где пересекаются оси первый и второй главных компонент, не обязательно совпадает или близка к положению на графике последнего общего предка популяций.
Монголы: Кочевые народы Монголии.
Тибетцы: Народы Тибета.
Эскимосы: Народы, населяющие арктические регионы Северной Америки, Гренландии и северо-востока Сибири.
На-дене: Североамериканская языковая группа.
Уральцы: Языковая семья, включающая финно-угорскую и самодийскую подсемьи (название от Уральских гор).
Северные турки: Турция.
Айны: Отдельная аборигенная группа в Японии.
Дравиды: Народы Южной Индии и севера Шри-Ланки.
Чукчи: Народы северо-востока Сибири.
Лапландцы: Кочевые скотоводы севера Скандинавии.
Баски: Народ, обитающий на севере Центральной Испании (считающийся наиболее гомогенной расовой группой; согласно данным Кавалли-Сфорца – они ранние европейцы со своим собственным уникальным языком).
Сардинцы: Аборигены острова Сардиния к западу от Италии.
Тайцы: Население Таиланда.
Полинезийцы: Население части Океании, включающей разбросанные острова в центральной и южной частях Тихого океана, приблизительно между Новой Зеландией, Гавайскими островами и островом Пасхи.
Меланезийцы: Обитатели островов в области к северо-востоку от Австралии и к югу от Экватора.
Кхмеры: Население Камбоджи.
Микронезийцы: Народы, населяющие часть Океании в западной части Тихого океана, включающей острова к востоку от Филиппин и к северу от Экватора.
Малазийцы: Население южной части Малайского полуострова и острова Борнео.
Берберы: Аборигены Северной Африки.
Койсаны: Кочевые охотники Юго-Западной Африки.
Мбути: Африканские пигмеи.
Банту: Близкие по языкам народы Центральной и Южной Африки.
Нило-сахарцы: Близкие по языкам африканцы тропической Африки от Нигерии до Кении.

Как можно видеть на рисунке 7–3, европейцы попадают в правый верхний угол, африканцы располагаются в правом нижнем углу, а монголоиды располагаются с левой стороны[82]. Нгуни, сотхо и тсонга – это народы Южной Африки, блака – это пигмеи из Нигера, а мбути – пигмеи из Республики Конго (Киншаса). Обратите внимание, что центральная часть графика практически пуста, даже при том, что представлены средние значения показателей. Это имеет место вследствие того, что хотя все эти популяции были некогда единой популяцией, генетические различия между ними возрастают на пути их становления разными биологическими видами.
На рисунке 7–4 (см. вклейку) приведена карта из той же работы, показывающая группировку человеческих популяций на основе их генетического сходства. Африканцы показаны желтым цветом, европеоиды зеленым, монголоиды темно-синим, а австралийские аборигены коричнево-красным. У народов Северной Африки имеется европеоидная составляющая, не нашедшая должного отражения на этой карте. На карте четко видно, что генетически схожие народы, как и следовало ожидать[83], населяют одни и те же географические регионы. Другими словами, раса – это реальность.
На рисунке 7–5 (см. вклейку) представлены сравнительные генетические расстояния (числа в нижней части рисунка) между африканскими (синие на графиках А и В и зеленые на графике C) и европейскими популяциями (красные на графиках А и В и желтые на графике C). Вертикальными черными линиями на вершинах графиков отмечены средние, а горизонтальными черными линиями на вершинах показаны стандартные отклонения.
На рисунке 7–5 видно, что при сравнении аллелей, распространенных в Африке и в Европе (график C), популяции могут быть разделены с точностью, близкой к 100 %. Средние значения отстоят дальше всего друг от друга и генетические расстояния наибольшие на графике C. На графиках A и B средние значения ближе друг к другу, генетические расстояния меньше и перекрывание значительно выше, потому что у этих популяций гораздо меньше уникальных аллелей, использовавшихся при сравнениях.
Возвращаясь к численному выражению генетических расстояний, отметим, что Кавалли-Сфорца и соавторы (Cavalli-Sforza et al., 1994) составили таблицы, где показаны генетические расстояния, разделяющие между собой 2000 расовых групп. В таблице 7–1 представлены генетические расстояния (на основе использования метода расчета FST) между отдельными избранными популяциями в процентах (умноженными на 10 000), например, банту – австралийские аборигены FST = 0,3272 % (по Salter, 2003, с. 64, на основе данных Cavalli-Sforza et al., 1994).
Таблица 7–1

Обратите внимание, что африканские народы банту и сан, живущие в Южной Африке, генетически близки. Восточные африканцы, живущие на Африканском Роге, являются проникшими в Африку обитателями Евразии, и они ближе к не-африканцам, чем к каким-либо другим африканцам, и являются популяцией, генетически в наибольшей степени отстоящей от всех остальных африканцев. Отметьте также, что наименее родственными друг другу народами являются банту и австралийские аборигены.
Как только были собраны данные о генетических расстояниях, стало возможным рассчитать другие результаты, некоторые из которых весьма впечатляют. Например, мы все считаем, что мать находится в более тесном родстве со своим собственным ребенком, чем к с каким-либо другим, но это не всегда так. Для большинства обитателей Азии и для значительной части (по меньшей мере половины) белых европейцев его совместный ребенок-мулат с африканцем банту будет находиться с ним в меньшем родстве, чем взятый наугад ребенок его собственной расы! Объяснение этого странного результата простое – изоляция банту от жителей Евразии привела к тому, что эти две популяции значительно разошлись генетически. Вследствие скрещивания обитателей Евразии между собой по меньшей мере на протяжении десятков тысяч лет соседский ребенок будет иметь больше общих аллелей с аборигеном Евразии, чем последний со своим собственным ребенком-мулатом[84].
Генетическое расстояние – это способ численно выразить, насколько генетически различны два индивида или две популяции. Как объясняется в Главе 3, все люди имеют одни и те же гены, например, все мы имеем ген, определяющий цвет глаз, но каждый ген в среднем имеет 14 различных последовательностей нуклеотидов, называемых «аллелями». Для определения генетического расстояния между двумя индивидами можно подсчитать число аллелей, которые у них различаются. В случае популяций подсчитывается число индивидов в каждой из них, имеющих определенные аллели (предпочтительно использовать большое число аллелей для повышения точности), и результаты выражаются математически. Если индивид имеет близнеца, его аллели будут идентичны аллелям его близнеца и генетическое расстояние между ними будет равно нулю. Если другой индивид является вашим ребенком, одинаковой будет, по крайней мере, половина аллелей. (Если его другой родитель имеет некоторые аллели, общие с вашими, у ребенка будет больше половины ваших аллелей.) Если брак является кровосмесительным, число общих с родительскими аллелей у детей будет выше, чем когда родители не состоят в родстве. Число общих аллелей в целом ниже у двоюродных братьев и сестер, еще ниже у не связанных родством представителей одного этноса или расы, и еще ниже у представителей разных рас и разных биологических видов, и оно продолжает снижаться по мере удаления от последнего общего предка (LCA) человека и других видов.
Если мы построим график генетического расстояния от вас (предположив, что вы европеоид) до других людей на планете, он может выглядеть аналогично изображенному на рисунке 7–1. На этом рисунке весьма приближенно показано, как быстро увеличивается генетическое расстояние при удалении от ближайших родственников. Затем имеет место существенное увеличение генетического расстояния между вами и монголоидами и еще намного большее его увеличение между вами и африканцами[80].
Пока что невозможно полностью проанализировать ДНК каждого человека на планете[81] и сравнить его ДНК с ДНК любого другого человека для определения того, сколько у них общих аллелей, но существуют упрощенные методики, дающие примерно одинаковые результаты. Генетическое расстояние между отдельными людьми и популяциями может быть рассчитано на основании анализа образцов ДНК (Salter, 2003). Собирая образцы ДНК в разных регионах всего мира и рассчитывая число однонуклеотидных полиморфизмов, ученые определяют генетические расстояния между разными популяциями, этносами, расами и биологическими видами. Числа в верхней части рисунка 7–2 (Cavalli-Sforza, 1991) показывают в процентах генетические расстояния (умноженные на 10 000) между разными человеческими популяциями, вычисленные с помощью модифицированного метода Нея.
Что касается трех основных рас, то на рисунке 7–2 показано, что африканцы из тропической Африки и из любых других регионов состоят с остальными людьми в наименьшем родстве, а аборигены Севера Евразии и Юго-Восточной Азии занимают второе место по генетическому расстоянию между собой. Обратите внимание, что к европеоидам отнесены аборигены Северной Африки (обитающие вокруг Средиземного моря), Северо-Западной Азии (Ближнего Востока) и индусы. Также заметьте, что аборигены Северо-Восточной Азии и американские индейцы состоят в более тесном родстве с европеоидами, чем с аборигенами Юго-Восточной Азии.
На рисунке 7–3 отмечены положения 42 популяций человечества относительно двух осей, показывающих различия по двум высоко вариабельным участкам мтДНК (Cavalli-Sforza, 1994, с. 82). Оси первой и второй главных компонент делят данные на две половины, обуславливающие наибольшую и вторую по величине дисперсии, соответственно (см. Википедия, Метод главных компонент). Африканцы находятся на одном конце обеих осей, а все остальные на другом, потому что африканцы отличаются генетически от большинства других популяций. Так как некоторые популяции (аборигены Евразии) подверглись более значительной эволюции, чем остальные (африканцы), то точка, где пересекаются оси первый и второй главных компонент, не обязательно совпадает или близка к положению на графике последнего общего предка популяций.
Монголы: Кочевые народы Монголии.
Тибетцы: Народы Тибета.
Эскимосы: Народы, населяющие арктические регионы Северной Америки, Гренландии и северо-востока Сибири.
На-дене: Североамериканская языковая группа.
Уральцы: Языковая семья, включающая финно-угорскую и самодийскую подсемьи (название от Уральских гор).
Северные турки: Турция.
Айны: Отдельная аборигенная группа в Японии.
Дравиды: Народы Южной Индии и севера Шри-Ланки.
Чукчи: Народы северо-востока Сибири.
Лапландцы: Кочевые скотоводы севера Скандинавии.
Баски: Народ, обитающий на севере Центральной Испании (считающийся наиболее гомогенной расовой группой; согласно данным Кавалли-Сфорца – они ранние европейцы со своим собственным уникальным языком).
Сардинцы: Аборигены острова Сардиния к западу от Италии.
Тайцы: Население Таиланда.
Полинезийцы: Население части Океании, включающей разбросанные острова в центральной и южной частях Тихого океана, приблизительно между Новой Зеландией, Гавайскими островами и островом Пасхи.
Меланезийцы: Обитатели островов в области к северо-востоку от Австралии и к югу от Экватора.
Кхмеры: Население Камбоджи.
Микронезийцы: Народы, населяющие часть Океании в западной части Тихого океана, включающей острова к востоку от Филиппин и к северу от Экватора.
Малазийцы: Население южной части Малайского полуострова и острова Борнео.
Берберы: Аборигены Северной Африки.
Койсаны: Кочевые охотники Юго-Западной Африки.
Мбути: Африканские пигмеи.
Банту: Близкие по языкам народы Центральной и Южной Африки.
Нило-сахарцы: Близкие по языкам африканцы тропической Африки от Нигерии до Кении.
Как можно видеть на рисунке 7–3, европейцы попадают в правый верхний угол, африканцы располагаются в правом нижнем углу, а монголоиды располагаются с левой стороны[82]. Нгуни, сотхо и тсонга – это народы Южной Африки, блака – это пигмеи из Нигера, а мбути – пигмеи из Республики Конго (Киншаса). Обратите внимание, что центральная часть графика практически пуста, даже при том, что представлены средние значения показателей. Это имеет место вследствие того, что хотя все эти популяции были некогда единой популяцией, генетические различия между ними возрастают на пути их становления разными биологическими видами.
На рисунке 7–4 (см. вклейку) приведена карта из той же работы, показывающая группировку человеческих популяций на основе их генетического сходства. Африканцы показаны желтым цветом, европеоиды зеленым, монголоиды темно-синим, а австралийские аборигены коричнево-красным. У народов Северной Африки имеется европеоидная составляющая, не нашедшая должного отражения на этой карте. На карте четко видно, что генетически схожие народы, как и следовало ожидать[83], населяют одни и те же географические регионы. Другими словами, раса – это реальность.
На рисунке 7–5 (см. вклейку) представлены сравнительные генетические расстояния (числа в нижней части рисунка) между африканскими (синие на графиках А и В и зеленые на графике C) и европейскими популяциями (красные на графиках А и В и желтые на графике C). Вертикальными черными линиями на вершинах графиков отмечены средние, а горизонтальными черными линиями на вершинах показаны стандартные отклонения.
На рисунке 7–5 видно, что при сравнении аллелей, распространенных в Африке и в Европе (график C), популяции могут быть разделены с точностью, близкой к 100 %. Средние значения отстоят дальше всего друг от друга и генетические расстояния наибольшие на графике C. На графиках A и B средние значения ближе друг к другу, генетические расстояния меньше и перекрывание значительно выше, потому что у этих популяций гораздо меньше уникальных аллелей, использовавшихся при сравнениях.
Возвращаясь к численному выражению генетических расстояний, отметим, что Кавалли-Сфорца и соавторы (Cavalli-Sforza et al., 1994) составили таблицы, где показаны генетические расстояния, разделяющие между собой 2000 расовых групп. В таблице 7–1 представлены генетические расстояния (на основе использования метода расчета FST) между отдельными избранными популяциями в процентах (умноженными на 10 000), например, банту – австралийские аборигены FST = 0,3272 % (по Salter, 2003, с. 64, на основе данных Cavalli-Sforza et al., 1994).
Таблица 7–1
Обратите внимание, что африканские народы банту и сан, живущие в Южной Африке, генетически близки. Восточные африканцы, живущие на Африканском Роге, являются проникшими в Африку обитателями Евразии, и они ближе к не-африканцам, чем к каким-либо другим африканцам, и являются популяцией, генетически в наибольшей степени отстоящей от всех остальных африканцев. Отметьте также, что наименее родственными друг другу народами являются банту и австралийские аборигены.
Как только были собраны данные о генетических расстояниях, стало возможным рассчитать другие результаты, некоторые из которых весьма впечатляют. Например, мы все считаем, что мать находится в более тесном родстве со своим собственным ребенком, чем к с каким-либо другим, но это не всегда так. Для большинства обитателей Азии и для значительной части (по меньшей мере половины) белых европейцев его совместный ребенок-мулат с африканцем банту будет находиться с ним в меньшем родстве, чем взятый наугад ребенок его собственной расы! Объяснение этого странного результата простое – изоляция банту от жителей Евразии привела к тому, что эти две популяции значительно разошлись генетически. Вследствие скрещивания обитателей Евразии между собой по меньшей мере на протяжении десятков тысяч лет соседский ребенок будет иметь больше общих аллелей с аборигеном Евразии, чем последний со своим собственным ребенком-мулатом[84].