Страница:
Приведу простой пример того, как это, по моему представлению, происходит. Допустим, кора вашего головного мозга знает, что должна воспринять музыкальный интервал квинту. Колонки вашей зоны реагируют на отдельные интервалы, такие как «до-ми», «до-соль», «ре-ля» и так далее. Вам нужно решить, какие из колонок будут активными. Если зона выше говорит вам, что следует ожидать квинту, это активизирует клетки слоя 2 во всех колонках, представляющих квинты, то есть «до-соль», «ре-ля», «ми-си». Клетки слоя 2 из колонок, которые представляют другие интервалы, активизироваться не будут. Теперь вам нужно из колонок, представляющих квинты, выбрать только одну. Входными сигналами для вашей зоны будут отдельные ноты. Если «ре» была последней нотой, которую вы слышали, тогда все колонки, представляющие интервалы, в которых присутствует нота «ре», т. е. «ре-ми» и «ре-си», получают часть входного сигнала. Теперь в слое 2 активизировались все колонки, представляющие квинты, а в слое 4 мы получили часть сигнала во все колонки, представляющие интервалы, в которые входит нота «ре». Пересечение этих двух множеств и будет ответом на наш вопрос. Это будет колонка, представляющая интервал «ре-ля» (см. рис. 6.11).
Как кора головного мозга определяет пересечение? Мы раньше упоминали о том, что аксоны клеток слоев 2 и 3, выходя из коры головного мозга, формируют синапсы в слое 5. Аксоны, направляющиеся к слою 4 из более низких зон коры головного мозга, создают синапсы в слое 6. Пересечение этих двух синапсов (снизу вверх и сверху вниз) дает нам как раз то, что нужно. Клетки слоя 6, получающие эти два сигнала возбуждаются. Клетка слоя 6 «представляет мнение» зоны коры головного мозга о том, что происходит, т. е. специфический прогноз. Если бы клетка слоя 6 могла разговаривать, она бы сказала: «Я являюсь частью колонки, представляющей нечто. Конкретно в моем случае колонка представляет интервал „ре-ля“. У остальных колонок другие значения. Я представляю свою зону коры головного мозга. Когда я становлюсь активной, это означает, что, по нашему мнению, сейчас звучит интервал „ре-ля“, или же он прозвучит в ближайшем будущем. Возможно, я активизировалась, потому что сигнал снизу вверх, поступивший от ушей, заставил клетку слоя 4 моей колонки возбудить всю колонку. Или же моя активизация может означать, что мы узнали мелодию и прогнозируем следующий интервал. В любом случае моя задача состоит в том, чтобы довести до ведома низших зон коры головного мозга, что, по нашему мнению, происходит. Я представляю наше видение внешнего мира – независимо от того, является он настоящим или выдуманным».
Попробую описать указанный процесс с помощью визуальной картинки. Представьте себе два листа бумаги, усеянные множеством мелких дырочек. Дырочки на одном листе представляют колонки, у которых есть активные клетки слоев 2 и 3 (наш инвариантный прогноз). Дырочки на другом листе бумаги представляют колонки с частичным сигналом снизу. Если мы наложим листы бумаги друг на друга, то некоторые из дырочек совпадут, а некоторые – нет. Дырочки, которые совпали, представляют колонки, которые, как мы полагаем, должны активизироваться.
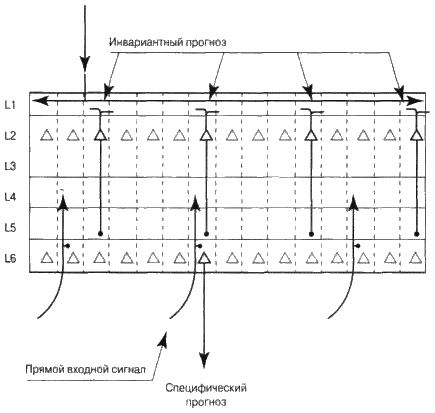
Рис. 6.11. Как зона коры головного мозга формирует специфические прогнозы, используя инвариантные репрезентации
Описанный механизм не только создает определенные прогнозы, он также разрешает неоднозначность входных сенсорных сигналов. Как мы помним из примеров с цветными листами бумаги и искаженным изображением, очень часто входные сигналы, полученные зоной коры головного мозга, являются неоднозначными. Механизм сочетания информации, поступающей снизу вверх и сверху вниз, помогает выбрать один из двух или более вариантов интерпретации. Как только вы приняли решение, каков будет выбор, вы сразу же транслируете этот выбор в область, расположенную ниже по иерархии.
Каждое мгновение нашей жизни каждая зона коры головного мозга сравнивает набор ожидаемых колонок, возбужденных сверху, с набором колонок, которые уже были возбуждены снизу. Там, где эти два набора пересекаются, создается наше восприятие. Если бы поступали совершенные сигналы снизу и у нас были бы совершенные прогнозы, тогда набор возбужденных колонок всегда совпадал бы с набором ожидаемых колонок. Но очень часто этого не происходит. Метод сочетания частичного прогноза и частичного входного сигнала позволяет определиться с неоднозначным входным сигналом, он позволяет заполнить пробелы в информации и остановить выбор на одном из альтернативных вариантов. Именно так мы комбинируем музыкальный интервал с последней услышанной нотой, чтобы спрогнозировать следующую ноту мелодии. Именно так мы определяемся, изображено на картинке два лица или ваза. Именно так мы разветвляем моторный поток на написание или произнесение вслух Геттисбергской речи.
И наконец, вдобавок к отправлению информации в более низкие зоны коры головного мозга, клетки слоя 6 могут посылать выходные сигналы назад, к клеткам слоя 4 той же колонки. Когда они это делают, наши прогнозы становятся входным сигналом. Именно этим мы занимаемся, размышляя или мечтая. Это позволяет нам видеть последовательности наших собственных прогнозов. Мы занимаемся этим, планируя свое будущее, вспоминая выступления или переживая о предстоящих событиях. Стефен Гросберг, длительное время занимавшийся моделированием деятельности коры головного мозга, называет это свернутой обратной связью, я же предпочитаю термин воображение.
И наконец, последний вопрос, который нам нужно затронуть, прежде чем перейти к следующему разделу. Я неоднократно подчеркивал, что чаще всего то, что мы видим, слышим, чувствуем, во многом зависит от наших собственных действий. То, что мы видим, зависит от того, на чем наши глаза остановились после очередной саккады, и от угла поворота головы. То, что мы ощущаем, зависит от того, как мы передвигаем пальцы и конечности. То, что мы слышим, иногда зависит от того, что мы делаем и говорим.
Значит, чтобы спрогнозировать, что мы почувствуем дальше, мы должны знать, что мы сейчас делаем. Моторное поведение и сенсорное восприятие тесно взаимосвязаны. Как нам удается составлять прогнозы, если то, что мы чувствуем, во многом является результатом наших собственных действий? К счастью, у данной проблемы существует неожиданное и простое решение, правда, многие его детали не очень понятны.
Неожиданное открытие: восприятие и поведение – это почти одно и то же. Как я уже упоминал раньше, большинство зон коры головного мозга (если не все) принимают участие в осуществлении движения.
Клеткам слоя 5, имеющим проекции в таламусе, а затем в слое 1, тоже свойственна моторная функция, поскольку у них одновременно есть проекции в моторных зонах «старого» мозга. Таким образом, знание о том, что только что произошло (как в сенсорном, так и в моторном плане), присутствует в слое 1.
Вторая неожиданность (являющаяся следствием первой): моторное поведение должно быть представлено в иерархии инвариантных репрезентаций. Вы генерируете движения, необходимые для осуществления определенного действия, не размышляя о нем во всех деталях. Когда моторная команда опускается вниз по иерархии, она трансформируется в сложные и подробные последовательности, необходимые для осуществления той деятельности, которая была запланирована. Это происходит как в сенсорной, так и в моторной зонах коры головного мозга, что размывает грань между ними. Если зона IT зрительной коры головного мозга воспринимает репрезентацию «нос», то простое действие переключения на репрезентацию «глаза» будет вызывать саккаду, необходимую для оправдания прогноза. Саккада, необходимая для того, чтобы перейти от зрительного восприятия носа к зрительному восприятию глаза, меняется в зависимости от расположения лица. Если лицо расположено поблизости, это требует большей саккады, и наоборот, более отдаленное лицо требует меньшей саккады. Наклоненное лицо требует саккады под углом, который отличается от используемого для лица без наклона. То, какой должна быть саккада, определяется, пока прогноз о том, что следующим мы увидим глаз, спускается к зоне V1. Чем дальше опустился прогноз, тем более точной становится саккада. Заканчивается все тем, что центральные ямки ваших глаз останавливается точно на объекте или же очень близко к нему.
Давайте рассмотрим еще один пример. Для того чтобы я физически переместился из гостиной на кухню, все, что нужно моему мозгу, – это мысленно переключиться с инвариантной репрезентации гостиной на инвариантную репрезентацию кухни. Данное переключение вызывает сложное разворачивание последовательностей. Процесс генерирования последовательности прогнозов того, что я увижу, услышу, почувствую, пока буду переходить из гостиной на кухню, тоже генерирует последовательность моторных команд, заставляющих меня перейти из гостиной на кухню и перемещать мой взгляд, пока я двигаюсь. Прогнозирование и моторное поведение идут рука об руку, в то время как потоки сигналов одновременно двигаются вверх и вниз по иерархии коры головного мозга. Каким бы странным это ни казалось, но, когда в дело вовлечено ваше поведение, ваши прогнозы не только предшествуют ощущению – они также его предопределяют. Прогнозирование перехода к следующему сигналу последовательности вызывает каскадное прогнозирование того, что вы должны пережить дальше. Когда каскадный прогноз разворачивается, он генерирует моторные команды, необходимые для осуществления прогноза. Воображение, прогнозирование и действие являются частями одного и того же разворачивания последовательностей в иерархии коры головного мозга.
Действие в воображении, параллельное разворачивание сенсорного и моторного поведения являются основой поведения, ориентированного на результат. Целенаправленное поведение, этот священный Грааль робототехники, изначально встроено в ткань коры головного мозга.
Конечно, мы можем отключить наше моторное поведение. Я могу представить себе, что вижу что-то, хотя на самом деле этого не вижу. Я могу представить себе, как иду на кухню, хотя на самом деле этого не делаю. Но действие всегда начинается в нашем воображении.
Поток вниз, поток вверх
На что способна обратная связь?
Как происходит процесс обучения
Как кора головного мозга определяет пересечение? Мы раньше упоминали о том, что аксоны клеток слоев 2 и 3, выходя из коры головного мозга, формируют синапсы в слое 5. Аксоны, направляющиеся к слою 4 из более низких зон коры головного мозга, создают синапсы в слое 6. Пересечение этих двух синапсов (снизу вверх и сверху вниз) дает нам как раз то, что нужно. Клетки слоя 6, получающие эти два сигнала возбуждаются. Клетка слоя 6 «представляет мнение» зоны коры головного мозга о том, что происходит, т. е. специфический прогноз. Если бы клетка слоя 6 могла разговаривать, она бы сказала: «Я являюсь частью колонки, представляющей нечто. Конкретно в моем случае колонка представляет интервал „ре-ля“. У остальных колонок другие значения. Я представляю свою зону коры головного мозга. Когда я становлюсь активной, это означает, что, по нашему мнению, сейчас звучит интервал „ре-ля“, или же он прозвучит в ближайшем будущем. Возможно, я активизировалась, потому что сигнал снизу вверх, поступивший от ушей, заставил клетку слоя 4 моей колонки возбудить всю колонку. Или же моя активизация может означать, что мы узнали мелодию и прогнозируем следующий интервал. В любом случае моя задача состоит в том, чтобы довести до ведома низших зон коры головного мозга, что, по нашему мнению, происходит. Я представляю наше видение внешнего мира – независимо от того, является он настоящим или выдуманным».
Попробую описать указанный процесс с помощью визуальной картинки. Представьте себе два листа бумаги, усеянные множеством мелких дырочек. Дырочки на одном листе представляют колонки, у которых есть активные клетки слоев 2 и 3 (наш инвариантный прогноз). Дырочки на другом листе бумаги представляют колонки с частичным сигналом снизу. Если мы наложим листы бумаги друг на друга, то некоторые из дырочек совпадут, а некоторые – нет. Дырочки, которые совпали, представляют колонки, которые, как мы полагаем, должны активизироваться.
Рис. 6.11. Как зона коры головного мозга формирует специфические прогнозы, используя инвариантные репрезентации
Описанный механизм не только создает определенные прогнозы, он также разрешает неоднозначность входных сенсорных сигналов. Как мы помним из примеров с цветными листами бумаги и искаженным изображением, очень часто входные сигналы, полученные зоной коры головного мозга, являются неоднозначными. Механизм сочетания информации, поступающей снизу вверх и сверху вниз, помогает выбрать один из двух или более вариантов интерпретации. Как только вы приняли решение, каков будет выбор, вы сразу же транслируете этот выбор в область, расположенную ниже по иерархии.
Каждое мгновение нашей жизни каждая зона коры головного мозга сравнивает набор ожидаемых колонок, возбужденных сверху, с набором колонок, которые уже были возбуждены снизу. Там, где эти два набора пересекаются, создается наше восприятие. Если бы поступали совершенные сигналы снизу и у нас были бы совершенные прогнозы, тогда набор возбужденных колонок всегда совпадал бы с набором ожидаемых колонок. Но очень часто этого не происходит. Метод сочетания частичного прогноза и частичного входного сигнала позволяет определиться с неоднозначным входным сигналом, он позволяет заполнить пробелы в информации и остановить выбор на одном из альтернативных вариантов. Именно так мы комбинируем музыкальный интервал с последней услышанной нотой, чтобы спрогнозировать следующую ноту мелодии. Именно так мы определяемся, изображено на картинке два лица или ваза. Именно так мы разветвляем моторный поток на написание или произнесение вслух Геттисбергской речи.
И наконец, вдобавок к отправлению информации в более низкие зоны коры головного мозга, клетки слоя 6 могут посылать выходные сигналы назад, к клеткам слоя 4 той же колонки. Когда они это делают, наши прогнозы становятся входным сигналом. Именно этим мы занимаемся, размышляя или мечтая. Это позволяет нам видеть последовательности наших собственных прогнозов. Мы занимаемся этим, планируя свое будущее, вспоминая выступления или переживая о предстоящих событиях. Стефен Гросберг, длительное время занимавшийся моделированием деятельности коры головного мозга, называет это свернутой обратной связью, я же предпочитаю термин воображение.
И наконец, последний вопрос, который нам нужно затронуть, прежде чем перейти к следующему разделу. Я неоднократно подчеркивал, что чаще всего то, что мы видим, слышим, чувствуем, во многом зависит от наших собственных действий. То, что мы видим, зависит от того, на чем наши глаза остановились после очередной саккады, и от угла поворота головы. То, что мы ощущаем, зависит от того, как мы передвигаем пальцы и конечности. То, что мы слышим, иногда зависит от того, что мы делаем и говорим.
Значит, чтобы спрогнозировать, что мы почувствуем дальше, мы должны знать, что мы сейчас делаем. Моторное поведение и сенсорное восприятие тесно взаимосвязаны. Как нам удается составлять прогнозы, если то, что мы чувствуем, во многом является результатом наших собственных действий? К счастью, у данной проблемы существует неожиданное и простое решение, правда, многие его детали не очень понятны.
Неожиданное открытие: восприятие и поведение – это почти одно и то же. Как я уже упоминал раньше, большинство зон коры головного мозга (если не все) принимают участие в осуществлении движения.
Клеткам слоя 5, имеющим проекции в таламусе, а затем в слое 1, тоже свойственна моторная функция, поскольку у них одновременно есть проекции в моторных зонах «старого» мозга. Таким образом, знание о том, что только что произошло (как в сенсорном, так и в моторном плане), присутствует в слое 1.
Вторая неожиданность (являющаяся следствием первой): моторное поведение должно быть представлено в иерархии инвариантных репрезентаций. Вы генерируете движения, необходимые для осуществления определенного действия, не размышляя о нем во всех деталях. Когда моторная команда опускается вниз по иерархии, она трансформируется в сложные и подробные последовательности, необходимые для осуществления той деятельности, которая была запланирована. Это происходит как в сенсорной, так и в моторной зонах коры головного мозга, что размывает грань между ними. Если зона IT зрительной коры головного мозга воспринимает репрезентацию «нос», то простое действие переключения на репрезентацию «глаза» будет вызывать саккаду, необходимую для оправдания прогноза. Саккада, необходимая для того, чтобы перейти от зрительного восприятия носа к зрительному восприятию глаза, меняется в зависимости от расположения лица. Если лицо расположено поблизости, это требует большей саккады, и наоборот, более отдаленное лицо требует меньшей саккады. Наклоненное лицо требует саккады под углом, который отличается от используемого для лица без наклона. То, какой должна быть саккада, определяется, пока прогноз о том, что следующим мы увидим глаз, спускается к зоне V1. Чем дальше опустился прогноз, тем более точной становится саккада. Заканчивается все тем, что центральные ямки ваших глаз останавливается точно на объекте или же очень близко к нему.
Давайте рассмотрим еще один пример. Для того чтобы я физически переместился из гостиной на кухню, все, что нужно моему мозгу, – это мысленно переключиться с инвариантной репрезентации гостиной на инвариантную репрезентацию кухни. Данное переключение вызывает сложное разворачивание последовательностей. Процесс генерирования последовательности прогнозов того, что я увижу, услышу, почувствую, пока буду переходить из гостиной на кухню, тоже генерирует последовательность моторных команд, заставляющих меня перейти из гостиной на кухню и перемещать мой взгляд, пока я двигаюсь. Прогнозирование и моторное поведение идут рука об руку, в то время как потоки сигналов одновременно двигаются вверх и вниз по иерархии коры головного мозга. Каким бы странным это ни казалось, но, когда в дело вовлечено ваше поведение, ваши прогнозы не только предшествуют ощущению – они также его предопределяют. Прогнозирование перехода к следующему сигналу последовательности вызывает каскадное прогнозирование того, что вы должны пережить дальше. Когда каскадный прогноз разворачивается, он генерирует моторные команды, необходимые для осуществления прогноза. Воображение, прогнозирование и действие являются частями одного и того же разворачивания последовательностей в иерархии коры головного мозга.
Действие в воображении, параллельное разворачивание сенсорного и моторного поведения являются основой поведения, ориентированного на результат. Целенаправленное поведение, этот священный Грааль робототехники, изначально встроено в ткань коры головного мозга.
Конечно, мы можем отключить наше моторное поведение. Я могу представить себе, что вижу что-то, хотя на самом деле этого не вижу. Я могу представить себе, как иду на кухню, хотя на самом деле этого не делаю. Но действие всегда начинается в нашем воображении.
Поток вниз, поток вверх
Давайте на минуту остановимся и поразмышляем над тем, как информация перемещается вниз и вверх в иерархии коры головного мозга. Когда вы передвигаетесь, потоки непрерывно меняющейся входной информации устремляются в нижние зоны коры головного мозга. Каждая зона пытается интерпретировать поток входящей информации как часть известной последовательности сигналов. Колонки пытаются предугадать свою активность. Если им это удается, они передают постоянный сигнал – название последовательности – в область, расположенную выше в иерархии. Зона коры головного мозга как бы говорит: «Я слышу песню, и вот вам ее название. Я сама разберусь с отдельными элементами».
Но что будет, если поступит неожиданный сигнал – нота, которой вы не ожидали? Что будет, если мы увидим что-то такое, чего на лице быть не должно? Неожиданный сигнал автоматически будет передан в область, стоящую выше в иерархии. Это случается, когда «вспыхивают» клетки слоя 3б, которые не были частью ожидаемой последовательности. Возможно, более высокая зона сможет понять сигнал как часть ее собственной последовательности. Возможно, она скажет: «Я вижу, что тут появилась новая нота. Вполне возможно, это первая нота следующей песни из альбома. Мне так кажется, поэтому я прогнозирую, что мы перешли к следующей песне из альбома. Низшая область, вот тебе название песни, которую, как мне кажется, ты сейчас слышишь». Но если не произойдет такого распознания, сигнал будет подниматься вверх по иерархии коры головного мозга, пока какая-нибудь из вышестоящих зон не идентифицирует сигнал как часть нормальной последовательности событий. Чем выше по иерархии поднимается сигнал, тем больше разных зон вовлекается в его обработку. Когда же наконец какая-то зона в верхней части иерархии считает, что она в состоянии обработать неожиданное событие, тогда формируется новый прогноз. Новый сигнал продвигается вниз по иерархии так далеко, как ему удается. Если новый прогноз оказался неверным, ошибка будет выявлена, и сигнал снова начнет подниматься вверх по иерархии до тех пор, пока не найдется область, которая интерпретирует его как часть своей непосредственной активной последовательности. Отсюда вывод: сигналы передвигаются вверх по иерархии, а прогнозы – вниз. В идеале, в хорошо известном и прогнозируемом мире, эти встречные потоки сигналов быстротечны и циркулируют преимущественно в нижних областях коры головного мозга. Мозг как можно быстрее пытается найти ту часть своей модели мира, которая бы соответствовала неожиданному входному сигналу. Только после этого он может понять входной сигнал и знать, чего ожидать дальше.
Когда я хожу по знакомой комнате в своем доме, лишь несколько ошибок поднимутся к верхним областям коры головного мозга. Хорошо усвоенные последовательности сигналов в моем доме обрабатываются на уровне нижних зон зрительной, соматосенсорной и моторной иерархии. Я знаю комнату настолько хорошо, что могу передвигаться по ней даже в кромешной тьме. Благодаря тому, что я хорошо знаю обстановку вокруг меня, большая часть моего неокортекса свободна и может размышлять о мозге и написании книг. В то же время если бы я находился в незнакомой мне комнате, особенно в такой, подобной которой я никогда не встречал, то мне не только пришлось бы постоянно присматриваться, куда идти. Неожиданные сигналы мощным потоком ринулись бы к верхним областям коры головного мозга. Чем больше мой сенсорный опыт не соответствует заученным последовательностям, тем больше ошибок будет возникать. В такой непривычной ситуации я не смогу думать о мозге, поскольку большая часть коры головного мозга будет занята проблемами ориентирования в комнате. С подобными ощущениями сталкиваются люди, сошедшие с трапа самолета в незнакомой стране. Дороги похожи на те, к которым вы привыкли, автомобили придерживаются левой стороны, язык, на котором все общаются, вам непонятен, а поиск туалета может стать сверхзадачей для всей коры головного мозга. Даже не пытайтесь готовиться к публичному выступлению, разгуливая по незнакомой стране.
Ощущение внезапного понимания (момент озарения) можно объяснить в рамках модели. Представьте, что вы смотрите на неоднозначную картинку. Пятна чернил и какие-то линии ничего вам не напоминают. Полнейшая бессмыслица. Когда мозг не может найти воспоминания, соответствующего полученному сигналу, человек пребывает в замешательстве. Ваши глаза блуждают по картинке. Входные сигналы поднялись на самый верх иерархии. Высшие зоны выдвигают множество самых разных гипотез, но, как только прогнозы начинают опускаться вниз по иерархии, каждая из гипотез конфликтует с входными сигналами, и мозг вынужден начинать все сначала. На протяжении всего состояния замешательства ваш мозг полностью поглощен тем, что пытается понять картинку. Наконец он формирует прогноз, который оказывается правильным. Когда это происходит, прогноз, сформированный в верхней зоне коры головного мозга, стремительно спускается вниз. Менее чем за секунду каждая зона получает последовательность, соответствующую полученным данным. Ни одна ошибка больше не транслируется к верхним зонам иерархии. Вы понимаете картинку и вместо точек и пятен видите далматинца (рис. 6.12).
Но что будет, если поступит неожиданный сигнал – нота, которой вы не ожидали? Что будет, если мы увидим что-то такое, чего на лице быть не должно? Неожиданный сигнал автоматически будет передан в область, стоящую выше в иерархии. Это случается, когда «вспыхивают» клетки слоя 3б, которые не были частью ожидаемой последовательности. Возможно, более высокая зона сможет понять сигнал как часть ее собственной последовательности. Возможно, она скажет: «Я вижу, что тут появилась новая нота. Вполне возможно, это первая нота следующей песни из альбома. Мне так кажется, поэтому я прогнозирую, что мы перешли к следующей песне из альбома. Низшая область, вот тебе название песни, которую, как мне кажется, ты сейчас слышишь». Но если не произойдет такого распознания, сигнал будет подниматься вверх по иерархии коры головного мозга, пока какая-нибудь из вышестоящих зон не идентифицирует сигнал как часть нормальной последовательности событий. Чем выше по иерархии поднимается сигнал, тем больше разных зон вовлекается в его обработку. Когда же наконец какая-то зона в верхней части иерархии считает, что она в состоянии обработать неожиданное событие, тогда формируется новый прогноз. Новый сигнал продвигается вниз по иерархии так далеко, как ему удается. Если новый прогноз оказался неверным, ошибка будет выявлена, и сигнал снова начнет подниматься вверх по иерархии до тех пор, пока не найдется область, которая интерпретирует его как часть своей непосредственной активной последовательности. Отсюда вывод: сигналы передвигаются вверх по иерархии, а прогнозы – вниз. В идеале, в хорошо известном и прогнозируемом мире, эти встречные потоки сигналов быстротечны и циркулируют преимущественно в нижних областях коры головного мозга. Мозг как можно быстрее пытается найти ту часть своей модели мира, которая бы соответствовала неожиданному входному сигналу. Только после этого он может понять входной сигнал и знать, чего ожидать дальше.
Когда я хожу по знакомой комнате в своем доме, лишь несколько ошибок поднимутся к верхним областям коры головного мозга. Хорошо усвоенные последовательности сигналов в моем доме обрабатываются на уровне нижних зон зрительной, соматосенсорной и моторной иерархии. Я знаю комнату настолько хорошо, что могу передвигаться по ней даже в кромешной тьме. Благодаря тому, что я хорошо знаю обстановку вокруг меня, большая часть моего неокортекса свободна и может размышлять о мозге и написании книг. В то же время если бы я находился в незнакомой мне комнате, особенно в такой, подобной которой я никогда не встречал, то мне не только пришлось бы постоянно присматриваться, куда идти. Неожиданные сигналы мощным потоком ринулись бы к верхним областям коры головного мозга. Чем больше мой сенсорный опыт не соответствует заученным последовательностям, тем больше ошибок будет возникать. В такой непривычной ситуации я не смогу думать о мозге, поскольку большая часть коры головного мозга будет занята проблемами ориентирования в комнате. С подобными ощущениями сталкиваются люди, сошедшие с трапа самолета в незнакомой стране. Дороги похожи на те, к которым вы привыкли, автомобили придерживаются левой стороны, язык, на котором все общаются, вам непонятен, а поиск туалета может стать сверхзадачей для всей коры головного мозга. Даже не пытайтесь готовиться к публичному выступлению, разгуливая по незнакомой стране.
Ощущение внезапного понимания (момент озарения) можно объяснить в рамках модели. Представьте, что вы смотрите на неоднозначную картинку. Пятна чернил и какие-то линии ничего вам не напоминают. Полнейшая бессмыслица. Когда мозг не может найти воспоминания, соответствующего полученному сигналу, человек пребывает в замешательстве. Ваши глаза блуждают по картинке. Входные сигналы поднялись на самый верх иерархии. Высшие зоны выдвигают множество самых разных гипотез, но, как только прогнозы начинают опускаться вниз по иерархии, каждая из гипотез конфликтует с входными сигналами, и мозг вынужден начинать все сначала. На протяжении всего состояния замешательства ваш мозг полностью поглощен тем, что пытается понять картинку. Наконец он формирует прогноз, который оказывается правильным. Когда это происходит, прогноз, сформированный в верхней зоне коры головного мозга, стремительно спускается вниз. Менее чем за секунду каждая зона получает последовательность, соответствующую полученным данным. Ни одна ошибка больше не транслируется к верхним зонам иерархии. Вы понимаете картинку и вместо точек и пятен видите далматинца (рис. 6.12).
На что способна обратная связь?
Десятки лет нам было известно, что связи в коре головного мозга являются взаимными. Если у зоны А есть проекция в зоне Б, то у зоны Б есть проекция в зоне А. Часто аксонов обратной связи оказывается больше, чем волокон прямой связи. Несмотря на то что все признают существование такого феномена, считается, что обратная связь играет второстепенную, или «модуляторную», роль в работе мозга. Мысль о том, что сигнал обратной связи на может постоянно и точно активизировать различные группы клеток слоя 2, не имеет большой поддержки в рядах нейробиологов.
Почему так? Частично потому, что, как я уже упоминал, если вы не считаете функцию прогнозирования центральной функцией головного мозга, то не возникает особой необходимости обращать внимание на обратную связь. Если вы предполагаете, что информация движется прямолинейно, к моторной системе, зачем вам эта обратная связь? Еще одна причина игнорирования обратной связи – то, что сигнал обратной, связи широко распространяется в слое 1. Интуитивно мы ожидаем, что сигнал, рассеянный в большой зоне, будет оказывать очень слабое влияние на большое количество нейронов. В самом деле, у мозга есть несколько таких модуляторных сигналов, которые не оказывают влияния на отдельные нейроны, зато меняют общие характеристики (например, возбудимость).

Рис. 6.12. Вы видите далматинца?
И последняя причина игнорирования обратной связи обусловлена тем, как многие нейробиологи представляют себе работу нейронов. У типичных нейронов имеются тысячи или даже десятки тысяч синапсов. Одни из них расположены ближе к телу клетки, другие – дальше. Синапсы, расположенные ближе к телу клетки, оказывают сильное влияние на ее импульсацию. Дюжина активных синапсов, расположенных у тела клетки, может заставить ее генерировать потенциал действия – электрический разряд. В этом не ничего нового. Тем не менее большинство синапсов находятся вдали от тела клетки. Они разбросаны по разветвленной структуре дендритов клетки. Поскольку эти синапсы расположены далеко от тела клетки, ученые склонны полагать, что импульс, пришедший на один из этих синапсов, будет очень слабо либо вообще не будет влиять на активность клетки. Импульс с отдаленного синапса рассеется по дороге, не достигнув тела клетки.
Как правило, информация, поднимающаяся вверх по иерархии коры головного мозга, передается по синапсам, расположенным близко к телу клетки. В этом случае сведения с большей вероятностью будут успешно передаваться от зоны к зоне. Как правило, обратная информация, поступающая вниз по иерархии коры головного мозга, передается по синапсам, расположенным дальше от тела клетки. Клетки слоев 2, 3 и 5 посылают дендриты в слой 1 и формируют там много синапсов. Слой 1 представляет собой совокупность синапсов, но все они значительно отдалены от тел клеток слоев 2, 3 и 5. Более того, каждая отдельная клетка слоя 2 формирует чрезвычайно мало синапсов с каждым отдельным волокном обратной связи (если формирует их вообще). Следовательно, некоторые ученые могут возразить по поводу того, что непродолжительный сигнал в слое 1 может вызвать упорядоченное возбуждение группы клеток слоев 2, 3 и 5, однако моя теория базируется именно на этом предположении.
Дилемма разрешается благодаря тому, что фактическое поведение нейронов отличается от поведения, приписываемого им классической моделью. В последние годы многие ученые придерживаются мнения, что синапсы на отдаленных тонких дендритах могут играть самую непосредственную роль в возбуждении клеток. В указанных моделях эти дальние синапсы функционируют не так, как синапсы на более толстых дендритах, расположенных вблизи тела клетки. Например, если два синапса на тонком дендрите расположены очень близко друг к другу, они будут действовать как «детектор совпадения». Другими словами, если оба синапса получили входящий импульс близко во времени, то они могут оказать большое влияние на клетку, хотя и расположены далеко от ее тела. Они могут заставить тело клетки генерировать разряд. Поведение дендритов нейронов до сих пор остается загадкой для науки, поэтому я не могу остановиться на этом вопросе более подробно. Важным моментом является то, что модель коры головного мозга «Память-предсказание» требует, чтобы синапсы, отдаленные от тела клетки, могли идентифицировать специфические сигналы.
Не слишком прозорливым, почти глупым было заявление, что большинство из тысяч синапсов нейрона играют только модуляторную роль. Обширная обратная связь и огромное количество синапсов не могли возникнуть без веской причины. Используя наши знания, мы можем сказать, что типичный нейрон обладает способностью выучить сотни точных совпадений благодаря волокнам обратной связи, когда они создают синапсы на тонких дендритах. Это, в свою очередь, означает, что колонка коры головного мозга очень гибка в отношении того, какие сигналы обратной связи могут вызвать ее активность. Таким образом, каждое отдельное свойство можно точно ассоциировать с тысячами различных объектов и последовательностей. Моя модель требует того, чтобы обратная связь была быстрой и точной. Клетки должны возбуждаться, когда они обнаруживают любое количество точных совпадений на отдаленных дендритах. Новая нейронная модель основывается именно на этом.
Почему так? Частично потому, что, как я уже упоминал, если вы не считаете функцию прогнозирования центральной функцией головного мозга, то не возникает особой необходимости обращать внимание на обратную связь. Если вы предполагаете, что информация движется прямолинейно, к моторной системе, зачем вам эта обратная связь? Еще одна причина игнорирования обратной связи – то, что сигнал обратной, связи широко распространяется в слое 1. Интуитивно мы ожидаем, что сигнал, рассеянный в большой зоне, будет оказывать очень слабое влияние на большое количество нейронов. В самом деле, у мозга есть несколько таких модуляторных сигналов, которые не оказывают влияния на отдельные нейроны, зато меняют общие характеристики (например, возбудимость).
Рис. 6.12. Вы видите далматинца?
И последняя причина игнорирования обратной связи обусловлена тем, как многие нейробиологи представляют себе работу нейронов. У типичных нейронов имеются тысячи или даже десятки тысяч синапсов. Одни из них расположены ближе к телу клетки, другие – дальше. Синапсы, расположенные ближе к телу клетки, оказывают сильное влияние на ее импульсацию. Дюжина активных синапсов, расположенных у тела клетки, может заставить ее генерировать потенциал действия – электрический разряд. В этом не ничего нового. Тем не менее большинство синапсов находятся вдали от тела клетки. Они разбросаны по разветвленной структуре дендритов клетки. Поскольку эти синапсы расположены далеко от тела клетки, ученые склонны полагать, что импульс, пришедший на один из этих синапсов, будет очень слабо либо вообще не будет влиять на активность клетки. Импульс с отдаленного синапса рассеется по дороге, не достигнув тела клетки.
Как правило, информация, поднимающаяся вверх по иерархии коры головного мозга, передается по синапсам, расположенным близко к телу клетки. В этом случае сведения с большей вероятностью будут успешно передаваться от зоны к зоне. Как правило, обратная информация, поступающая вниз по иерархии коры головного мозга, передается по синапсам, расположенным дальше от тела клетки. Клетки слоев 2, 3 и 5 посылают дендриты в слой 1 и формируют там много синапсов. Слой 1 представляет собой совокупность синапсов, но все они значительно отдалены от тел клеток слоев 2, 3 и 5. Более того, каждая отдельная клетка слоя 2 формирует чрезвычайно мало синапсов с каждым отдельным волокном обратной связи (если формирует их вообще). Следовательно, некоторые ученые могут возразить по поводу того, что непродолжительный сигнал в слое 1 может вызвать упорядоченное возбуждение группы клеток слоев 2, 3 и 5, однако моя теория базируется именно на этом предположении.
Дилемма разрешается благодаря тому, что фактическое поведение нейронов отличается от поведения, приписываемого им классической моделью. В последние годы многие ученые придерживаются мнения, что синапсы на отдаленных тонких дендритах могут играть самую непосредственную роль в возбуждении клеток. В указанных моделях эти дальние синапсы функционируют не так, как синапсы на более толстых дендритах, расположенных вблизи тела клетки. Например, если два синапса на тонком дендрите расположены очень близко друг к другу, они будут действовать как «детектор совпадения». Другими словами, если оба синапса получили входящий импульс близко во времени, то они могут оказать большое влияние на клетку, хотя и расположены далеко от ее тела. Они могут заставить тело клетки генерировать разряд. Поведение дендритов нейронов до сих пор остается загадкой для науки, поэтому я не могу остановиться на этом вопросе более подробно. Важным моментом является то, что модель коры головного мозга «Память-предсказание» требует, чтобы синапсы, отдаленные от тела клетки, могли идентифицировать специфические сигналы.
Не слишком прозорливым, почти глупым было заявление, что большинство из тысяч синапсов нейрона играют только модуляторную роль. Обширная обратная связь и огромное количество синапсов не могли возникнуть без веской причины. Используя наши знания, мы можем сказать, что типичный нейрон обладает способностью выучить сотни точных совпадений благодаря волокнам обратной связи, когда они создают синапсы на тонких дендритах. Это, в свою очередь, означает, что колонка коры головного мозга очень гибка в отношении того, какие сигналы обратной связи могут вызвать ее активность. Таким образом, каждое отдельное свойство можно точно ассоциировать с тысячами различных объектов и последовательностей. Моя модель требует того, чтобы обратная связь была быстрой и точной. Клетки должны возбуждаться, когда они обнаруживают любое количество точных совпадений на отдаленных дендритах. Новая нейронная модель основывается именно на этом.
Как происходит процесс обучения
У всех нейронов из всех слоев коры головного мозга есть синапсы, большинство из которых может модифицироваться под влиянием опыта. Можно с уверенностью сказать, что процесс обучения и запоминания происходит во всех слоях, во всех колонках и во всех зонах коры головного мозга. Я упоминал ранее правило «обучения Хебба», названное так в честь Дональда Хебба. Сущность этого правила очень проста: когда два нейрона активизируются одновременно, синаптические связи между ними усиливаются. (Его легко запомнить как фразу «Возбуждены вместе – связаны вместе».) Сейчас мы знаем, что Хебб был прав. Конечно, ничего в природе не бывает настолько простым, свойства настоящего мозга сложнее. Наша нервная система работает на основе многих вариаций «обучения Хебба». Некоторые синапсы изменяют силу связи в ответ на незначительные отклонения координации нейронных сигналов. Некоторые синаптические изменения оказываются краткосрочными, некоторые – долгосрочными. Хебб очертил основные рамки, а не создал конечную теорию процесса обучения, причем эти рамки оказались очень и очень полезными.
Принципы «обучения Хебба» объясняют большинство проявлений поведения коры головного мозга, о которых мы говорили в данной главе. Помните, еще в 1970-х годах было установлено, что автоассоциативная память при помощи алгоритма «обучения Хебба» может усвоить пространственные сигналы и их последовательности. Основная проблема заключалась в том, что автоассоциативная память не в состоянии справиться с отклонениями. Согласно теории, предложенной в нашей книге, кора головного мозга нашла выход из такой ситуации, частично путем складывания автоассоциативных запоминаний в иерархию, а частично – путем использования сложной столбчатой архитектуры. Эта глава почти полностью посвящена иерархической структуре мозга и ее работе, поскольку именно иерархия обусловливает высокую мощность коры головного мозга. Вместо того чтобы пускаться в пространные объяснения, как та или иная клетка учится тем или иным вещам, я хотел бы раскрыть несколько общих принципов обучения в иерархии.
У новорожденного кора головного мозга «не знает» фактически ничего. Она не знает его языка, его культуры, его дома, его города, песен, людей, вместе с которыми он будет расти, – ничего. Всю эту информацию – структуру внешнего мира – нужно выучить. Основными составляющими обучения являются формирование классификаций и создание последовательностей. Эти два взаимодополняющих компонента взаимодействуют друг с другом. По мере того как одна зона коры головного мозга учит последовательности, входящие сигналы, которые она посылает в клетки слоя 4 зоной выше в иерархии, меняются. Клетки слоя 4 учатся формировать новые классификации, что изменяет сигнал, проецируемый назад, в слой 1, зоной ниже, а это влияет на последовательности.
Основой формирования последовательностей является группирование паттернов, которые относятся к одному и тому же объекту. Один из способов – группировать сигналы, следующие друг за другом. Когда ребенок держит в руке игрушку и медленно ее переворачивает, его мозг может с уверенностью считать, что непрерывно изменяющееся изображение на сетчатке все равно принадлежит одному и тому же объекту, а значит, изменяющийся набор сигналов можно сгруппировать вместе. Иногда нужна внешняя подсказка, чтобы определить, что сигналы принадлежат одному и тому же целому. Чтобы выучить, что яблоки и бананы принадлежат к фруктам, а морковь и картофель – нет, нужен учитель, который помог бы сгруппировать указанные объекты как фрукты. В любом случае ваш мозг создает последовательности сигналов, принадлежащих одному целому. Однако когда зона мозга уже создала последовательности, то входной сигнал, поступающий в следующую область, изменяется. Если раньше он представлял отдельные сигналы, то теперь представляет группу сигналов. Входной сигнал превращается из ноты в мелодию, из букв в слово, из носов в лица и так далее. Поскольку сигналы снизу вверх становятся более объектно-ориентированными, высшая зона коры головного мозга теперь может запоминать последовательности этих объектов высшего порядка. Там, где раньше строились последовательности букв, теперь строятся последовательности слов. Неожиданным результатом процесса обучения является то, что на протяжении повторного обучения представления объектов передвигаются вниз по иерархии. В первые годы вашей жизни «записи» о мире формируются в высших зонах коры головного мозга. Однако по мере вашего дальнейшего обучения они перемещаются во все более низкие зоны иерархии коры головного мозга. Не то чтобы мозг их передвигал туда, просто он должен учить их заново, снова и снова. (Я не хочу сказать, что все запоминания обязательно начинаются с высших зон иерархии коры головного мозга. В реальной жизни усвоение информации выглядит сложнее. Я считаю, что классификация сигналов слоя 4 начинается снизу и передвигается вверх. По мере передвижения формируются последовательности, которые опускаются вниз. Запоминания последовательностей опускаются все ниже и ниже в коре головного мозга.) Когда простые репрезентации передвигаются вниз, высшие зоны коры головного мозга получают возможность учить новые, более сложные, сигналы.
Принципы «обучения Хебба» объясняют большинство проявлений поведения коры головного мозга, о которых мы говорили в данной главе. Помните, еще в 1970-х годах было установлено, что автоассоциативная память при помощи алгоритма «обучения Хебба» может усвоить пространственные сигналы и их последовательности. Основная проблема заключалась в том, что автоассоциативная память не в состоянии справиться с отклонениями. Согласно теории, предложенной в нашей книге, кора головного мозга нашла выход из такой ситуации, частично путем складывания автоассоциативных запоминаний в иерархию, а частично – путем использования сложной столбчатой архитектуры. Эта глава почти полностью посвящена иерархической структуре мозга и ее работе, поскольку именно иерархия обусловливает высокую мощность коры головного мозга. Вместо того чтобы пускаться в пространные объяснения, как та или иная клетка учится тем или иным вещам, я хотел бы раскрыть несколько общих принципов обучения в иерархии.
У новорожденного кора головного мозга «не знает» фактически ничего. Она не знает его языка, его культуры, его дома, его города, песен, людей, вместе с которыми он будет расти, – ничего. Всю эту информацию – структуру внешнего мира – нужно выучить. Основными составляющими обучения являются формирование классификаций и создание последовательностей. Эти два взаимодополняющих компонента взаимодействуют друг с другом. По мере того как одна зона коры головного мозга учит последовательности, входящие сигналы, которые она посылает в клетки слоя 4 зоной выше в иерархии, меняются. Клетки слоя 4 учатся формировать новые классификации, что изменяет сигнал, проецируемый назад, в слой 1, зоной ниже, а это влияет на последовательности.
Основой формирования последовательностей является группирование паттернов, которые относятся к одному и тому же объекту. Один из способов – группировать сигналы, следующие друг за другом. Когда ребенок держит в руке игрушку и медленно ее переворачивает, его мозг может с уверенностью считать, что непрерывно изменяющееся изображение на сетчатке все равно принадлежит одному и тому же объекту, а значит, изменяющийся набор сигналов можно сгруппировать вместе. Иногда нужна внешняя подсказка, чтобы определить, что сигналы принадлежат одному и тому же целому. Чтобы выучить, что яблоки и бананы принадлежат к фруктам, а морковь и картофель – нет, нужен учитель, который помог бы сгруппировать указанные объекты как фрукты. В любом случае ваш мозг создает последовательности сигналов, принадлежащих одному целому. Однако когда зона мозга уже создала последовательности, то входной сигнал, поступающий в следующую область, изменяется. Если раньше он представлял отдельные сигналы, то теперь представляет группу сигналов. Входной сигнал превращается из ноты в мелодию, из букв в слово, из носов в лица и так далее. Поскольку сигналы снизу вверх становятся более объектно-ориентированными, высшая зона коры головного мозга теперь может запоминать последовательности этих объектов высшего порядка. Там, где раньше строились последовательности букв, теперь строятся последовательности слов. Неожиданным результатом процесса обучения является то, что на протяжении повторного обучения представления объектов передвигаются вниз по иерархии. В первые годы вашей жизни «записи» о мире формируются в высших зонах коры головного мозга. Однако по мере вашего дальнейшего обучения они перемещаются во все более низкие зоны иерархии коры головного мозга. Не то чтобы мозг их передвигал туда, просто он должен учить их заново, снова и снова. (Я не хочу сказать, что все запоминания обязательно начинаются с высших зон иерархии коры головного мозга. В реальной жизни усвоение информации выглядит сложнее. Я считаю, что классификация сигналов слоя 4 начинается снизу и передвигается вверх. По мере передвижения формируются последовательности, которые опускаются вниз. Запоминания последовательностей опускаются все ниже и ниже в коре головного мозга.) Когда простые репрезентации передвигаются вниз, высшие зоны коры головного мозга получают возможность учить новые, более сложные, сигналы.