Большая Советская Энциклопедия (ТК)
кровью
и
лимфой
внутренней средой организма. Из Т. ж. клетки поглощают необходимые питательные вещества и выводят в неё продукты обмена. Химический состав, физические и биологические свойства Т. ж. специфичны для отдельных органов и соответствуют их морфологическим и функциональным особенностям. Т. ж. близка к плазме крови, но содержит меньше белка (около 1,5
гна 100
мл), другое количество электролитов, ферментов, продуктов обмена (
метаболитов). Состав и свойства Т. ж. отличаются определённым постоянством (см.
гомеостаз
), что предохраняет клетки органов и тканей от воздействий, связанных с изменениями состава крови. Проникновение в Т. ж. из крови веществ, необходимых для питания тканей, и удаление из неё метаболитов осуществляются через
гисто-гематические барьеры
. Оттекая от органов в лимфатические сосуды, Т. ж. превращается в лимфу. Объём Т. ж. у кролика равен 23—25% массы тела, у человека — 23—29% (в среднем 26,5%). К Т. ж. многие авторы относят спинномозговую жидкость, жидкость передней камеры глаза, сердечной сумки, плевральной полости и др.
Схема диффузии веществ между капиллярами и клетками тела через тканевую жидкость, омывающую клетки: 1 — капилляр; 2 — эндотелий капилляра; 3 — тканевая жидкость; 4 — тканевые клетки; 5 — эритроциты.
антигенами
. Благодаря существующему в природе генетическому разнообразию клетки и ткани любых двух особей различаются по множеству антигенов тканевой совместимости (называемых также антигенами гистосовместимости, трансплантационными антигенами, изо- или аллоантигенами). В эволюционном ряду Т. н., основанная на иммунологических реакциях, встречается впервые у низших позвоночных — миног и миксин. [В примитивной форме в виде разнообразных биохимических реакций, направленных на поддержание постоянства внутренней среды (
гомеостаза
), «несовместимость» генетически разнородных организмов наблюдается даже у одноклеточных.] Все позвоночные животные имеют развитую систему иммунологического распознавания и устранения чужеродных антигенов. При пересадке органа или ткани (
трансплантации
) через короткий срок после приживления происходит отторжение трансплантата, повреждаемого лимфоцитами и цитотоксичными
антителами
организма-хозяина (реципиента). Если иммунная система реципиента повреждена специальными препаратами — иммуно-депрессантами, то лимфоциты донора, содержащиеся в трансплантате (например, в пересаживаемом костном мозге), атакуют и повреждают ткани хозяина. Явление Т. н. можно наблюдать в условиях эксперимента вне организма, например лимфоциты, полученные от разных людей, при совместном культивировании взаимно активируют друг друга к превращению в лимфобласты и к делению.
У человека судьба трансплантата определяется различиями по 3 основным системам аллоантигенов: антигенам
групп крови
ABO, групповым антигенам Р и лейкоцитарным антигенам HL-A (первые буквы англ. human leucocyte antigen — лейкоцитарные антигены человека). Чем меньше антигенные различия между донором и реципиентом по этим системам, тем легче добиться длительного приживления трансплантата и иммунологической
толерантности
. Наибольшие трудности подбора совместимых органов и тканей связаны с системой HL-A, включающей не менее 60 разных аллоантигенов. Аллоантигены HL-A представляют собой
гликопротеиды
(молекулярная масса свыше 200000), встроенные в мембраны всех клеток организма и находящиеся в растворённом виде в плазме крови. Молекула аллоантигена образована 2 полипептидными цепями, которые связаны с углеводной частью; аллоантигены различаются только аминокислотной последовательностью длинной полипептидной цепи (молекулярная масса около 30 000). Короткая полипептидная цепь (молекулярная масса около 10000), сходная у разных аллоантигенов, представляет собой молекулу
b
2-микроглобулина, который встречается в плазме и в свободном виде (аминокислотная последовательность Pz- микроглобулина повторяет последовательности постоянных участков лёгкой и тяжёлой цепей
иммуноглобулинов
). Многокомпонентность системы HL-A приводит к тому, что даже прямые родственники (кроме однояйцевых близнецов) могут различаться по набору аллоантигенов. Уже известно свыше 9 тыс. различных таких наборов. Биологическое значение различий по системам гистосовместимости ещё полностью не выяснено. Полагают, что столь сложная система поверхностных клеточных антигенов в сочетании с чрезвычайно чувствительной реакцией иммунной системы на чужеродные аллоантигены служит механизмом устранения злокачественных клеток собственного организма, появляющихся в результате
мутации
. По мнению австралийского иммунолога Ф.
Бёрнета
, не будь этого механизма защиты, рак превратился бы в инфекционное заболевание, передающееся от человека к человеку. Аллоантигенные различия между супругами, между сперматозоидом и яйцеклеткой, между плодом и материнским организмом могут быть важным фактором
естественного отбора
. Слияние сперматозоида с яйцеклеткой происходит, по-видимому, не случайно, а яйцеклетка «выбирает» более «совместимый» сперматозоид, что создаёт селективные преимущества для определённых наборов HL-A. Во время беременности иммунная система матери отвечает образованием антител на аллоантигены плода, унаследованные от отца; в плаценте же имеет место нечто подобное слабой реакции трансплантата против хозяина, что, однако, как правило, не приводит к аборту. Установлено также, что ряд заболеваний, в патогенезе которых имеет значение наследственность (лейкозы, лимфогранулематоз, красная волчанка, псориаз и аллергические заболевания), значительно чаще встречаются у лиц с определёнными наборами HL-A. Образование аллоантигенов HL-A кодируется
аллелями
трёх локусов, расположенных в 6-й хромосоме.
Лабораторное определение аллоантигенов системы HL-A (типирование тканей) осуществляется при помощи наборов моноспецифических, соответствующим образом очищенных аллоиммунных сывороток. Их готовят из сывороток крови много рожавших женщин, больных, которым часто переливали кровь, или добровольцев, которым пересаживали кожу или вводили донорские лимфоциты. Содержащиеся в типирующих сыворотках антитела к HL-A дают серологическую реакции с типируемыми лимфоцитами, что позволяет судить о наличии или отсутствии на их поверхности соответствующих аллоантигенов.
Совместимы только генетически однородные ткани, например ткани однояйцевых близнецов. Чтобы сделать совместимыми ткани генетически различающихся особей, нужно каким-то образом вмешаться в выражение генов гистосовместимости, вызвать подавление (репрессию) одних генов и компенсировать деятельность недостающих генов, а это остаётся пока невыполнимой задачей. При разведении лабораторных животных путём близкородственного скрещивания (брат — сестра, дети — родители) сравнительно легко можно вывести линии генетически сходных, а потому и совместимых особей. В трансплантационной иммунологии преодоление Т. н. достигается подавлением иммунного ответа реципиента и созданием иммунологической толерантности. Это не устраняет несовместимости как таковой, но обеспечивает сосуществование генетически разнородных тканей. Особые надежды возлагаются на создание иммунологической толерантности путём введения реципиенту небольших доз очищенных антигенов гистосовместимости в сочетании с иммунодепрессантами. У человека и ряда лабораторных животных (мыши) существует генетическая, структурная и функциональная взаимосвязь между Т. н. и способностью к иммунологическому ответу. См. также
Иммуногенетика
,
Иммунология
.
Лит.:Брондз Б. Д., Иммунологическое распознавание и реакции клеточного иммунитета in vitro, «Успехи современной биологии», 1972, т. 73, 11; Введение в иммуногенетику, пер. с англ., М., 1975; Batchelor J. R., Brent L., Histocompatibility in transplantation immunity, в кн.: Immuno-genicity, Amst. — L., 1972: Nathanson S. G., Histocompatibility antigens, в кн.: Transplantation, Phil., 1972; Immunological aspects of transplantation surgery, Lancaster, 1973; Immunological approaches to fertility control, [Stockh.], 1974.
А. Н. Мац.
Филатовым
. Согласно его концепции в сохраняемой на холоде (или консервируемой иным способом) ткани в процессе её адаптации к неблагоприятным условиям среды накапливаются вещества с высокой биологической активностью —
биогенные стимуляторы
, которые и определяют лечебный эффект Т. т. Как и
протеинотерапия
, Т. т. относится к неспецифическим методам лечения. Активируя иммунные и регенераторные функции организма, она нередко оказывается эффективной при вяло протекающих патологических процессах различной природы — воспалительных, дегенеративных, атрофических и др. В современной медицине Т. т. находит применение главным образом при некоторых глазных и кожных болезнях.
В ветеринарии Т. т. применяется с лечебной целью, а также для повышения продуктивности с.-х. животных: при длительно незаживающих ранах, язвах, некоторых болезнях кожи, лёгких и др.; в качестве стимуляторов при откорме молодняка крупного рогатого скота и свиней, для повышения молочной продуктивности коров и шёрстной продуктивности овец.
Лит.:Филатов В. П., Оптическая пересадка роговицы и тканевая терапия, М., 1945; Калашник И. А., Тканевая терапия в ветеринарии, М., 1960.
макроэргических соединений
. Т. д. отличают от внешнего
дыхания
—совокупности физиологических процессов, обеспечивающих поступление в организм кислорода и выведение из него углекислого газа. Многие ферменты, катализирующие эти реакции, находятся в особых клеточных органоидах —
митохондриях
.
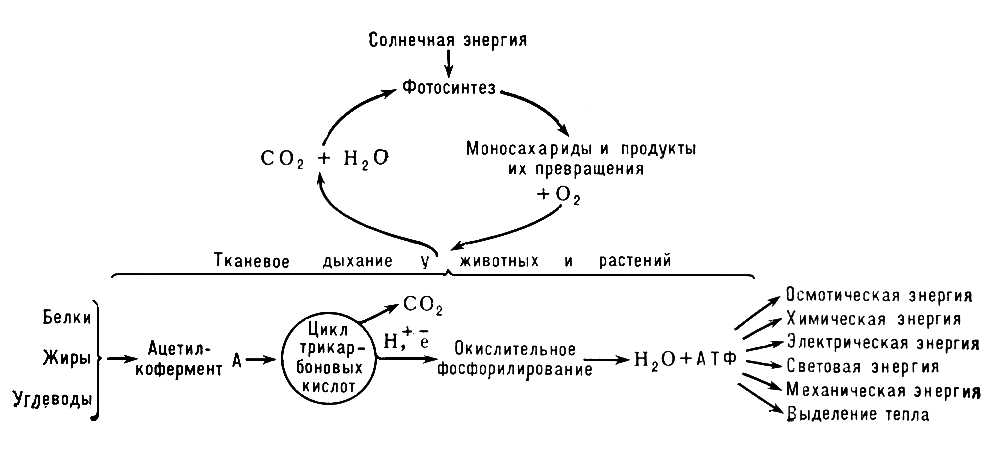
На все проявления
жизни
—рост, движение, раздражимость, самовоспроизведение и др. — организм расходует энергию. Формой энергии, пригодной для использования клетками, является энергия химических связей (главным образом фосфатных) в макроэргических соединениях — аденозинтрифосфорной кислоте (АТФ) и др. Для синтеза АТФ необходим приток энергии извне. По способам извлечения энергии существует принципиальное различие между
автотрофными организмами
и
гетеротрофными организмами
. Клетки зелёных растений — наиболее типичных автотрофов — в процессе фотосинтеза используют энергию солнечного света для синтеза АТФ и глюкозы. (Образование из глюкозы более сложных молекул происходит в клетках растений также в процессе Т. д.) В клетках гетеротрофов — животных и человека — единственным источником энергии является энергия химических связей молекул пищевых веществ. Молекулы различных соединений, выполняющие роль биологического «топлива» (глюкоза, жирные кислоты, некоторые аминокислоты), образовавшись в клетках животного организма или поступив в кровь из пищеварительного тракта, претерпевают ряд последовательных химических превращений. В процессе Т. д. можно наметить три основные стадии: 1) окислительное образование ацетилкофермента А (активная форма уксусной кислоты) из пировиноградной кислоты (промежуточный продукт расщепления глюкозы), жирных кислот и аминокислот; 2) разрушение ацетильных остатков в
трикарбоновых кислот цикле
с освобождением 2 молекул углекислого газа и 4 пар атомов водорода, частично акцептируемых коферментами
никотинамидадениндинуклеотидом
и
флавинадениндинуклеотидом
и частично переходящих в раствор в виде протонов; 3) перенос электронов и протонов к молекулярному кислороду (образование H
2O) — процесс, катализируемый набором дыхательных ферментов и сопряжённый с образованием АТФ (так называемое
окислительное фосфорилирование
). Первые две стадии подготавливают третью, в ходе которой в результате последовательных окислительно-восстановительных реакций происходит освобождение основной части энергии, вырабатываемой в клетке. При этом около 50% энергии в результате окислительного фосфорилирования запасается в форме богатых энергией связей АТФ, а остальная часть её выделяется в виде тепла.
Т. д. обеспечивает образование и постоянное пополнение АТФ в клетках. В случае недостатка в снабжении клеток животных и человека кислородом запасы АТФ не исчерпываются сразу. Их пополнение может происходить в результате включения дополнительных механизмов — систем анаэробного (без участия кислорода) распада углеводов —
гликолиза
и гликогенолиза. Однако этот путь энергетически во много раз менее эффективен и не может обеспечить функции и целостность структуры органов и тканей. Биологическая роль Т. д. не исчерпывается существенным вкладом в энергетический обмен организма. На различных его этапах образуются молекулы органических соединений, используемых клетками в качестве промежуточных продуктов для различных биосинтезов. См. также
Аденозинфосфорные кислоты
,
Биоэнергетика
,
Обмен веществ
,
Окисление биологическое
.
Лит.:Северин С. Е., Биологическое окисление и окислительное фосфорилирование, в кн.: Химические основы процессов жизнедеятельности, М., 1962; Ленинджер А., Превращение энергии в клетке, в кн.: Живая клетка, пер. с англ., 2 изд., М., 1962; его же. Биохимия, пер. с англ., М., 1974; Скулачев В. П., Аккумуляция энергии в клетке, М., 1969; Вилли К., Детье В., Биология. (Биологические процессы и законы), пер. с англ., М., 1974.
В. Г. Иванова.
Схема превращения энергии в живых клетках: тканевое дыхание, образование АТФ и пути его использования.
Печатание тканей
). Различают цилиндрическую Т. м. с медными гравированными печатными валами и машины для печатания сетчатыми шаблонами.
Наиболее распространены цилиндрическая Т. м. Основные рабочие органы этих машин — свободно вращающийся чугунный пустотелый цилиндр (грузовик), на который накладывается ткань при печатании, и один или несколько (для многокрасочной печати) печатных валов, располагаемых вокруг грузовика. Поверхность грузовика имеет эластичное, упругое покрытие (так называемый печатный стол), состоящее из 10—16 слоев специальной ткани и слоя
кирзы
, которая предохраняется чехлом от закрашивания. Краска на печатный вал наносится с помощью вращающейся щётки или валика, избыток её счищается стальной пластиной — раклей. В процессе работы печатные валы прижимаются к непрерывно движущемуся полотну ткани; скорость движения ткани достигает 150
м/мин.
Принцип работы Т. м. с сетчатыми шаблонами основан на протирании или продавливании краски с помощью ракли через шаблон — тонкую сетку (плоскую или в виде цилиндра). Сетка покрыта лаковой плёнкой в местах, которые должны быть непроницаемы для краски (в соответствии с рисунком). Скорость движения ткани на машинах с плоскими шаблонами 3,5—20
м/мин, с цилиндрическим — 45—70
м/мин(иногда до 100
м/мин).
Лит.:Бельцов В. М., Технологическое оборудование отделочных фабрик текстильной промышленности, Л., 1974.
М. Н. Кириллова.
эпителий
, образует покровы тела и оболочки внутренних органов. Производные ее выполняют секреторную функцию, составляя, например, основную массу печени, поджелудочной железы.
Соединительная ткань
, в том числе и Т. внутренней среды, осуществляет трофическую и защитную функции организма. Производные соединит. Т. — хрящ и кость — несут у позвоночных животных опорную функцию, образуя скелет.
Мышечная ткань
выполняет двигательные функции, перемещая организм и вызывая сократительные движения его органов.
Нервная ткань
регулирует и координирует жизнедеятельность всех Т., воспринимает сигналы из внешней среды и определяет ответные реакции организма. Развитие каждого типа Т. — результат определённого
гистогенеза
, протекающего в эмбриональном периоде. Во многих Т. гистогенезы продолжаются и у взрослых животных, обеспечивая
регенерацию
, а иногда и рост Т. Специфические для каждого органа функции осуществляются обычно одной Т. или даже некоторыми специализированными ее клетками. Но в любом органе взаимодействуют различные Т., способствуя трофике и координации основных функциональных элементов. Активность тканевых клеток зависит как от непосредственных их контактов Т., так и от отдаленных гормональных и нервных влияний. У низших многоклеточных Т. не столь строго детерминированы, как у высших. Эволюция организмов привела к специализации клеток, взаимообусловленности их функционирования и самого существования в многотканевой системе. Однако моделируя окружение клеток, можно не только обеспечить их жизнь вне организма, но и многие гистогенезы (см.
Культуры тканей
), что стаяло одним из основных методов изучения тканей. Т. животных изучает гистология.
Лит. см. при статьях
Гистология
,
Гистогенез
.
В.
Я.
Бродский.
Т. растений. Рост растения и развитие его внутренней структуры обусловлены деятельностью образовательной Т., или меристемы, производные которой претерпевают сложную структурную и функциональную дифференцировку, превращаясь в элементы постоянных Т. Классификации постоянных Т. основываются на морфологических, функциональных, генетических и др. признаках. Различают, например, Т. паренхимные (см.
Паренхима
) и прозенхимные (см.
Прозенхима
). Постоянные Т. относят к трём системам: покровной, проводящей и основной, появление которых в онтогенезе растений отражает главные этапы внутренней дифференцировки растительного организма в процессе эволюции. По наиболее распространённой физиологической классификации Т., предложенной Г.
Габерландтом
, постоянные Т. составляют системы: покровную, представленную
эпидермисом
,
пробкой
и
коркой
, механическую, включающую
колленхиму
, состоящую из живых паренхимных клеток с неравномерно утолщёнными стенками, и
склеренхиму
, представленную одревесневшими волокнами и более или менее изодиаметрическими
склереидами
; абсорбционную, осуществляющую поглощение веществ с помощью ризоидов, корневых волосков, образованных
эпиблемой
, многослойного покрова (веламена) воздушных корней орхидных; ассимиляционную, состоящую из паренхимных клеток с обилием хлоропластов; проводящую, представленную
ксилемой
, осуществляющей проведение воды, и
флоэмой
, участвующей в перемещении органических веществ; запасающую, состоящую из паренхимных клеток; секреторную, включающую
гидатоды
,
млечники
, вместилища выделений различного происхождения; систему проветривания, представленную
межклетниками
,
устьицами
,
чечевичками
. Все Т., кроме покровной, проводящей и системы проветривания, можно считать разновидностями основной Т. Ткани растений изучает
анатомия растений
.
Лит.:Имс А. Дж., Мак Даниэльс Л. Г., Введение в анатомию растений, пер. с англ., М. — Л., 1935; Крашенинников Ф. Н., Лекции по анатомии растений, М. — Л., 1937; Бородин И. П., Курс анатомии растений, 5-е изд., М. — Л., 1938; Раздорский В. Ф., Анатомия растений, М., 1949; Яценко-Хмелевский А. А., Краткий курс анатомии растений, М., 1961; Эсау К., Анатомия растений, пер. с англ., М., 1969.
Л. И. Лотова.
декоративно-прикладного искусства
. Орнаментация Т. х., строящаяся обычно по принципу ритмичных повторов (см.
Раппорт
), достигается либо путём переплетения нитей, либо набивным способом (см.
Набойка
), а также вышивкой. Все 3 способа орнаментации применялись с глубокой древности и были известны почти всем народам. Художественное оформление тканей обусловливается их назначением и техническими возможностями производства; узоры Т. х. отражают стилистическое своеобразие различных эпох в развитии искусства и национальных художественных школ (см. также
Ковёр
).
Т. х. Древнего Востока и античного мира известны по памятникам изобразительного искусства и литературы, а также по отдельным образцам тканей, найденным в раскопках (например, фрагменты древнеегипетских льняных узорных тканей 2-го тыс. до н. э., фрагменты античных тканей из городов Северного Причерноморья). Высокой художественностью с древних времен славились китайские, главным образом шёлковые и «золотные» (с металлической нитью), ткани, известные с конца 2-го тыс. до н. э. [сохранились фрагменты узорчатых тканей 5—3 вв. до н. э. из раскопок в Чанша, богатых по расцветке тканей эпохи Хань (206 до н. э. — 220 н. э.) и др.]. Широкое распространение получили изготовлявшиеся в 16 и особенно в 17 вв. в Китае шёлковая «камка-китайка» (русское название), различные виды бархата (шёлковые
основа
и
уток
) и т.д. Орнамент китайских тканей — геометрические узоры, стилизованные мотивы растительного и звериного мира, символические изображения (круг, дракон, облака), иероглифы. К китайским Т. х. близки (по материалу, выработке, орнаменту) японские ткани. На протяжении веков (известны с 3-го тыс. до н. э.) славились индийские хлопчатобумажные (реже полушёлковые) ткани с набивным узором, шерстяные узорные и тонкие льняные ткани, орнамент которых состоял главным образом из растительных мотивов (часто с обилием деталей), а также из повторяющихся сюжетных сцен. Иран с древних времён вырабатывал изысканные по мастерству шерстяные и шёлковые ткани: сохранились фрагменты шёлковых тканей эпохи Сасанидов (3—7 вв.); их узор состоял обычно из медальонов (кругов, овалов и т.д.) с изображениями апофеоза царской власти, сцен охоты, фантастических животных. В 15—17 вв. высоко ценились иранские шёлковые и «золотные» ткани — так называемые объярь,
атлас
,
бархат
(основа бумажная); для их орнаментов характерны стилизованные мотивы растительного и животного мира, сцены из эпоса и т.д. Сохранилось много образцов тканей коптского Египта (4—7 вв.) с изображением различных религиозных сцен. Орнаментация византийских Т. х. испытала значительное воздействие позднеантичного и сасанидского искусства. Для узоров византийских тканей типичны круги с орлами, колесницами, библейскими сценами и т.д. В арабских странах выделывались шёлковые и «золотные» Т. х., узорные и гладкие (атлас). Своеобразны турецкие атласы и бархаты (имеющие бумажную основу) 15—18 вв., обычно с крупным узором из «опахал», полумесяцев, цветов гвоздики и др.
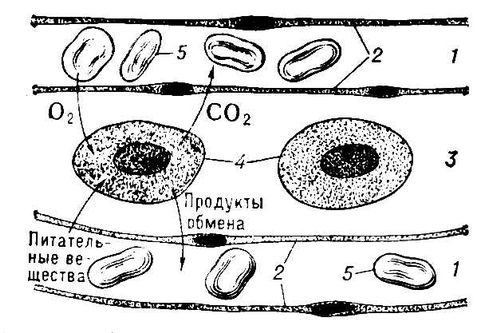 Схема диффузии веществ между капиллярами и клетками тела через тканевую жидкость, омывающую клетки: 1 — капилляр; 2 — эндотелий капилляра; 3 — тканевая жидкость; 4 — тканевые клетки; 5 — эритроциты.
Схема диффузии веществ между капиллярами и клетками тела через тканевую жидкость, омывающую клетки: 1 — капилляр; 2 — эндотелий капилляра; 3 — тканевая жидкость; 4 — тканевые клетки; 5 — эритроциты.