Страница:
Постепенно стали говорить не о динамичности возбуждения и динамичности торможения как самостоятельных свойствах нервной системы, а об их балансе с преобладанием либо первой, либо второй, причем между динамичностью возбуждения и торможения была найдена обратно пропорциональная зависимость: чем сильнее выражен один процесс, тем слабее проявлен другой (Э. А. Голубева, 1972). В ее работах показатель динамичности торможения по В. Д. Небылицыну (частота альфа-ритма ЭЭГ) рассматривается как показатель баланса по динамичности. В других исследованиях частота альфа-ритма рассматривается уже в качестве показателя активации (Е. П. Гусева и Н. Ф. Шляхта, 1974). Термин «динамичность» постепенно исчезает из статей по дифференциальной психофизиологии. Очевидно, это происходит не случайно, поскольку фоновые показатели ЭЭГ зависят не только от динамичности, но и от других типологических свойств.
Отметим еще мнение Д. Грея (1968), считающего, что и сила нервной системы, и динамичность могут представлять собой два подфактора, относящихся к большому единому фактору, т. е. выступают двумя проявлениями одного и того же физиологического явления. Справедливость такого предположения видна из того, что обнаружения слабости нервной системы и высокой динамичности возбуждения сходны, на это обратил внимание В. С. Мерлин. Найдено сходство проявления динамичности и лабильности в психологических феноменах (Э. А. Голубева и В. Д. Рождественская, 1969). Собственно, в этом нет ничего удивительного, так как быстрота возникновения возбуждения является составной частью того, что называется лабильностью.
Согласно сказанному, попытка выделить динамичность как новое свойство нервной системы не имеет серьезных оснований. Быстрота возникновения нервных процессов – это лишь одно из проявлений свойства силы нервной системы. Собственно, В. Д. Небылицын предвидел возможность исчезновения какого-либо подобного свойства из описанной им структуры. Он писал, например, что можно «представить себе и такую возможность, когда атрибуты какого-то предполагаемого свойства в результате экспериментального исследования будут отобраны у него и переданы другим параметрам нервной деятельности, и это предполагаемое свойство, в конца концов, окажется вычеркнутым из списка основных свойств нервной системы» (1966, с. 16).
Таким образом, пока мы не ушли далеко от той классификации свойств нервной системы, которая была предложена И. П. Павловым.
4.4. Парциальные и общие свойства нервной системы
Глава 5
5.1. Сила нервной системы
Отметим еще мнение Д. Грея (1968), считающего, что и сила нервной системы, и динамичность могут представлять собой два подфактора, относящихся к большому единому фактору, т. е. выступают двумя проявлениями одного и того же физиологического явления. Справедливость такого предположения видна из того, что обнаружения слабости нервной системы и высокой динамичности возбуждения сходны, на это обратил внимание В. С. Мерлин. Найдено сходство проявления динамичности и лабильности в психологических феноменах (Э. А. Голубева и В. Д. Рождественская, 1969). Собственно, в этом нет ничего удивительного, так как быстрота возникновения возбуждения является составной частью того, что называется лабильностью.
Согласно сказанному, попытка выделить динамичность как новое свойство нервной системы не имеет серьезных оснований. Быстрота возникновения нервных процессов – это лишь одно из проявлений свойства силы нервной системы. Собственно, В. Д. Небылицын предвидел возможность исчезновения какого-либо подобного свойства из описанной им структуры. Он писал, например, что можно «представить себе и такую возможность, когда атрибуты какого-то предполагаемого свойства в результате экспериментального исследования будут отобраны у него и переданы другим параметрам нервной деятельности, и это предполагаемое свойство, в конца концов, окажется вычеркнутым из списка основных свойств нервной системы» (1966, с. 16).
Таким образом, пока мы не ушли далеко от той классификации свойств нервной системы, которая была предложена И. П. Павловым.
4.4. Парциальные и общие свойства нервной системы
Говоря о свойствах нервной системы (или точнее – о свойствах нервных центров), нельзя понимать дело так, что какое-то изучаемое свойство относится ко всей нервной системе как аморфному образованию. Есть основания полагать, что хотя любой функциональной системе мозга (возбудительной, тормозной) присущи все свойства, однако степень их выраженности в разных системах (мозговых структурах) различается. В этой связи и возник вопрос об общих и парциальных свойствах нервной системы.
Если типологические особенности отражают определенные функциональные системы мозга, то спрашивается: к какой именно функциональной мозговой системе (или, по словам У. У. Ухтомского, «функциональному органу») относится данная методика, с помощью которой изучаются типологические особенности. Не может ли быть так, что разные методики, предъявляя обследуемым разные смысловые задачи, адресуются разным функциональным отделам мозга? Отсюда возникает важная для практики задача: установить, одинакова ли валидность разных методик, изучающих, например, свойство силы нервной системы.
Также рождается и теоретический вопрос: являются ли изучаемые свойства парциальными (т. е. отражающими только одну функциональную систему) или общими (т. е. относящимися ко всему мозгу в целом)?
Представление о парциальных свойствах нервной системы выдвинул Б. М. Теплов. Он писал, что у человека менее всего можно ожидать полного совпадения типологических параметров в разных анализаторах, а также в первой и второй сигнальной системах. Действительно, сопоставление типологических различий по свойству силы в различных анализаторах (зрительном и слуховом) не показало совпадения диагнозов. Тем самым был нанесен существенный удар по господствовавшим в то время представлениям, согласно которым свойства нервной системы характеризуют ее всю в целом. Теперь и наличие общемозговых свойств вызывало сомнение. По этому поводу В. Д. Небылицын заметил:
Должны ли мы теперь рассматривать СНС как параметры лишь узкого представительства, способные характеризовать только те мозговые структуры, к которым адресуются применяемые стимулы, или все-таки существуют такие, хотя бы относительно общие «сверханализаторные» характеристики мозга, которые могут служить основой для нейрофизиологической интерпретации целостных особенностей индивидуального поведения? (1976, с. 209).
В. Д. Небылицын считал, что парциальность может быть обусловлена тремя моментами:
1) регистрацией разных эффекторных выражений рефлекторной деятельности (эффекторный аспект);
2) применением раздражителей различной модальности (анализаторный аспект);
3) использованием различных подкрепляющих воздействий (в случае изучения типологических особенностей с помощью вырабатываемых условных рефлексов).
Таким образом, парциальность свойств нервной системы (а точнее – типологических особенностей их проявления) может быть обусловлена тем, что в применяемых для их диагностики реакциях участвуют различные нервные структуры.
Роль различий в подкреплении, сказывающихся в проявлении парциальности типологических особенностей, была установлена еще А. Г. Ивановым-Смоленским (1935). Применяя три подкрепления – ориентировочное, пищевое и оборонительное, сотрудники этого исследователя обнаружили, что полное совпадение результатов по быстроте образования положительных и тормозных условных рефлексов было только в 22% случаев, а при двух подкреплениях совпадения регистрировались лишь в 50% случаев. Поэтому уже А. Г. Иванов-Смоленский говорил о парциальных типах нервной системы. Аналогичные результаты в исследовании с привлечением людей получил польский психолог Ян Стреляу (1982): только у 2 из 36 испытуемых диагноз типологических особенностей совпал при применении трех видов подкреплений: электрокожного, температурного и мышечного усилия.
В. Д. Небылицын и его сотрудники показали роль модальности сигнала для расхождения в результатах диагностики. Причем главную «вину» за проявление парциальности типологических особенностей авторы возложили на межанализаторные различия. Этому есть некоторые подтверждения. Например, В. С. Горожанин (1977) не выявил связи скорости бега и максимальной частоты движений с подвижностью нервных процессов в зрительном анализаторе, в то время как, по нашим данным, при использовании двигательных методик такая связь обнаруживается.
Однако Ян Стреляу обращает внимание, что расхождения в результатах диагностики могут быть связаны не с модальностью раздражителя, а с его силой. Действительно, фактор силы (интенсивности) раздражителя способен создать проблему парциальности искусственно, что отмечал и В. С. Мерлин: ведь физическая сила зрительного и слухового раздражителя в лаборатории Б. М. Теплова – В. Д. Небылицына не выравнивалась. Правда, частично это возражение снимается тем, что, как отмечал В. Д. Небылицын, расхождения в диагнозах силы нервной системы при использовании зрительного и звукового раздражителей было то в пользу одного, то в пользу другого анализатора (при одинаковой для всех испытуемых интенсивности того и другого раздражителя). Вместе с тем значение интенсивности раздражителей различной модальности полностью В. Д. Небылицыным не отвергалось. Он писал в связи с этим:
Межанализаторная парциальность – это парциальность по горизонтали. Есть основание говорить и о парциальности типологических особенностей по вертикали.
Так, Н. И. Красногорский строил классификацию типов высшей нервной деятельности, исходя из учета соотношений между корой и подкоркой. Он считал, что холерический тип – подкорковый, а флегматический – корковый тип высшей нервной деятельности. Выделялись им и центральные типы, отражающие равновесие между корой и подкоркой: сильный (сангвинический) и слабый меланхолический.
Идею о парциальности по вертикали типологических особенностей проявления свойств нервной системы разделял и В. С. Мерлин, тоже ссылаясь на соотношения коры и подкорки.
Можно полагать, что вертикальная парциальность проявляется и в двух видах баланса – «внешнего» и «внутреннего» (Е. П. Ильин, 1972в). Об этом свидетельствуют антагонистические взаимоотношения между ними, возникающие при ряде состояний (монотонии, психическом пресыщении): сдвиг одного баланса в сторону возбуждения сопровождается сдвигом другого баланса в сторону торможения. Это можно рассматривать как перераспределение активности (энергии) с одного уровня регуляции на другой.
Таким образом, вопрос о парциальности свойств нервной системы продолжает оставаться актуальным и требует дальнейшего изучения.
Однако, как отмечал В. Д. Небылицын, «особенности организации мозговой рецептивной системы принципиально, по самой ее функции, таковы, что ее функциональные параметры – частные свойства – едва ли могут служить нейрофизиологическими “измерениями” личности как целого, со всеми общими аспектами ее организации и поведения» (1976, с. 21). Соответственно он выдвинул гипотезу о существовании и общих свойств нервной системы, которые связаны с выделенной им регулирующей системой мозга.
К этой системе В. Д. Небылицын отнес передние отделы неокортекса и взаимодействующие с ними образования старой и древней коры головного мозга, а также его стволовой части. Структуры, составляющие этот комплекс, он рассматривал в известном приближении как единую мозговую систему, выполняющую функции регуляции и управления всеми процессами, протекающими в организме – от низших биологических до самых высших психических (таких как потребности и мотивация, эмоции и темперамент, программирование движения, и действий, интеллектуальное планирование и оценка результатов и т. д.). В отличие от анализаторной, регулирующая система не имеет прямого отношения к переработке сенсорных раздражителей.
Важными являются связи свойств этой системы со многими общеличностными характеристиками – моторной активностью, интеллектуальной активностью и с особенностями установки личности, в то время как анализаторные (парциальные) свойства часто таких связей не обнаруживают.
Следует подчеркнуть, что В. Д. Небылицын, выдвигая понятие об общих свойствах нервной системы, рассматривал их не как чисто корковые, а как комплексные, отражающие функциональную систему, включающие в себя различные уровни центральной нервной системы: лобную кору, лимбический мозг, ретикулярную формацию. Можно считать, что под общими свойствами В. Д. Небылицын понимал, скорее всего, свойства интегративных систем мозга, осуществляющих произвольные акты человека. При этом он отмечал: свойства, обозначенные им как общие, строго говоря, тоже являются частными, поскольку представляют собой параметры пусть весьма обширного, но все же ограниченного анатомически и функционально комплекса структур головного мозга. Интерпретация этих свойств как общих оправдана только тем значением, которое имеют образования регуляторной системы для психической жизни и поведения всего организма. Во всяком случае, они, по мнению В. Д. Небылицына, дают лучшее приближение к функции «нервной системы в целом», чем парциальные свойства. Как полагал исследователь, представление об этих общих свойствах поможет понять, что же такое свойства нервной системы в целом, – категорию, остающуюся, по его мысли, на современном уровне наших психофизиологических знаний чисто гипотетической конструкцией.
Об этом высказывании В. Д. Небылицына (об относительности «общих» свойств) необходимо помнить, так как в более поздних работах других авторов содержатся высказывания, противоречащие положениям этого ученого.
Например, отличной от позиции В. Д. Небылицына точки зрения придерживается В. М. Русалов (1979). Он считает, что общие свойства – это индивидуально-устойчивые особенности целостной общемозговой интеграции нервных процессов, вовлеченных в целостную произвольную психическую деятельность, в то время как частные – особенности локальной интеграции. Автор полагает более правильным не увязывать строго общие свойства с деятельностью лобного отдела, а рассматривать их как особенности регуляции всего мозга. Свойства переднего мозга могут быть лишь аналогичными, но не идентичными свойствам всей нервной системы.
Со второй половины ХХ в. идея общемозговых свойств все больше замыкается на активации целого мозга и связанной с ней чувствительностью. Предполагается (В. С. Мерлин, 1973; И. М. Палей, 1966), что чувствительность прямо отражает «первичное» общее свойство. Согласно другой точке зрения, «первичным» и в значительной степени интегральным свойством является активированность покоя, а чувствительность – вторичная характеристика, зависящая от уровня активации в покое нервной системы (Е. П. Ильин, 1979).
Свойства, выявляемые с помощью методик, в которых используются произвольные двигательные реакции человека, обнаруживают постоянные связи с деятельностными и поведенческими характеристиками людей, а свойства, выявляемые в зрительном и слуховом анализаторах таких связей часто не демонстрируют или же показывают те, которые противоречат здравому смыслу. Следовательно, для практических целей прогнозирования склонностей, способностей, устойчивости к неблагоприятным состояниям и т. д. парциальные (анализаторные) свойства малопригодны.
Если типологические особенности отражают определенные функциональные системы мозга, то спрашивается: к какой именно функциональной мозговой системе (или, по словам У. У. Ухтомского, «функциональному органу») относится данная методика, с помощью которой изучаются типологические особенности. Не может ли быть так, что разные методики, предъявляя обследуемым разные смысловые задачи, адресуются разным функциональным отделам мозга? Отсюда возникает важная для практики задача: установить, одинакова ли валидность разных методик, изучающих, например, свойство силы нервной системы.
Также рождается и теоретический вопрос: являются ли изучаемые свойства парциальными (т. е. отражающими только одну функциональную систему) или общими (т. е. относящимися ко всему мозгу в целом)?
Представление о парциальных свойствах нервной системы выдвинул Б. М. Теплов. Он писал, что у человека менее всего можно ожидать полного совпадения типологических параметров в разных анализаторах, а также в первой и второй сигнальной системах. Действительно, сопоставление типологических различий по свойству силы в различных анализаторах (зрительном и слуховом) не показало совпадения диагнозов. Тем самым был нанесен существенный удар по господствовавшим в то время представлениям, согласно которым свойства нервной системы характеризуют ее всю в целом. Теперь и наличие общемозговых свойств вызывало сомнение. По этому поводу В. Д. Небылицын заметил:
Должны ли мы теперь рассматривать СНС как параметры лишь узкого представительства, способные характеризовать только те мозговые структуры, к которым адресуются применяемые стимулы, или все-таки существуют такие, хотя бы относительно общие «сверханализаторные» характеристики мозга, которые могут служить основой для нейрофизиологической интерпретации целостных особенностей индивидуального поведения? (1976, с. 209).
В. Д. Небылицын считал, что парциальность может быть обусловлена тремя моментами:
1) регистрацией разных эффекторных выражений рефлекторной деятельности (эффекторный аспект);
2) применением раздражителей различной модальности (анализаторный аспект);
3) использованием различных подкрепляющих воздействий (в случае изучения типологических особенностей с помощью вырабатываемых условных рефлексов).
Таким образом, парциальность свойств нервной системы (а точнее – типологических особенностей их проявления) может быть обусловлена тем, что в применяемых для их диагностики реакциях участвуют различные нервные структуры.
Роль различий в подкреплении, сказывающихся в проявлении парциальности типологических особенностей, была установлена еще А. Г. Ивановым-Смоленским (1935). Применяя три подкрепления – ориентировочное, пищевое и оборонительное, сотрудники этого исследователя обнаружили, что полное совпадение результатов по быстроте образования положительных и тормозных условных рефлексов было только в 22% случаев, а при двух подкреплениях совпадения регистрировались лишь в 50% случаев. Поэтому уже А. Г. Иванов-Смоленский говорил о парциальных типах нервной системы. Аналогичные результаты в исследовании с привлечением людей получил польский психолог Ян Стреляу (1982): только у 2 из 36 испытуемых диагноз типологических особенностей совпал при применении трех видов подкреплений: электрокожного, температурного и мышечного усилия.
В. Д. Небылицын и его сотрудники показали роль модальности сигнала для расхождения в результатах диагностики. Причем главную «вину» за проявление парциальности типологических особенностей авторы возложили на межанализаторные различия. Этому есть некоторые подтверждения. Например, В. С. Горожанин (1977) не выявил связи скорости бега и максимальной частоты движений с подвижностью нервных процессов в зрительном анализаторе, в то время как, по нашим данным, при использовании двигательных методик такая связь обнаруживается.
Однако Ян Стреляу обращает внимание, что расхождения в результатах диагностики могут быть связаны не с модальностью раздражителя, а с его силой. Действительно, фактор силы (интенсивности) раздражителя способен создать проблему парциальности искусственно, что отмечал и В. С. Мерлин: ведь физическая сила зрительного и слухового раздражителя в лаборатории Б. М. Теплова – В. Д. Небылицына не выравнивалась. Правда, частично это возражение снимается тем, что, как отмечал В. Д. Небылицын, расхождения в диагнозах силы нервной системы при использовании зрительного и звукового раздражителей было то в пользу одного, то в пользу другого анализатора (при одинаковой для всех испытуемых интенсивности того и другого раздражителя). Вместе с тем значение интенсивности раздражителей различной модальности полностью В. Д. Небылицыным не отвергалось. Он писал в связи с этим:
Другое дело – изучение таких свойств нервной системы, как подвижность или динамичность. Здесь, действительно, для того чтобы исключить влияние параметра чувствительности – силы, следует уравнивать раздражители по их физиологической интенсивности (1966, с. 340).Теоретически парциальность может проявиться не от межанализаторных различий, а от того, что к одному анализатору адресуется своя задача, а к другому – иная (это зависит от методических приемов диагностики типологических особенностей). Роль данного обстоятельства отмечена В. И. Рождественской и др. (1969). Однако про такие случаи можно сказать, что эксперимент не был чистым и сравнение проведено некорректно. При одинаковости же заданий (как это имело место в работах В. Д. Небылицына по определению силы нервной системы или в исследовании М. И. Семенова и А. Х. Мамажанова (1972), сопоставлявших подвижность и баланс нервных процессов при двигательных и зрительных заданиях, идентичных по смыслу и процедуре выполнения) парциальность проявлялась вполне отчетливо. Так, в последней из указанных работ совпадение диагнозов типологических особенностей в двух выборках (73 студента и 50 школьников) было только в 42—56% случаев. Аналогичные данные на значительно большем материале с использованием тех же методических приемов были получены А. Г. Пинчуковым (1974).
Межанализаторная парциальность – это парциальность по горизонтали. Есть основание говорить и о парциальности типологических особенностей по вертикали.
Так, Н. И. Красногорский строил классификацию типов высшей нервной деятельности, исходя из учета соотношений между корой и подкоркой. Он считал, что холерический тип – подкорковый, а флегматический – корковый тип высшей нервной деятельности. Выделялись им и центральные типы, отражающие равновесие между корой и подкоркой: сильный (сангвинический) и слабый меланхолический.
Идею о парциальности по вертикали типологических особенностей проявления свойств нервной системы разделял и В. С. Мерлин, тоже ссылаясь на соотношения коры и подкорки.
Можно полагать, что вертикальная парциальность проявляется и в двух видах баланса – «внешнего» и «внутреннего» (Е. П. Ильин, 1972в). Об этом свидетельствуют антагонистические взаимоотношения между ними, возникающие при ряде состояний (монотонии, психическом пресыщении): сдвиг одного баланса в сторону возбуждения сопровождается сдвигом другого баланса в сторону торможения. Это можно рассматривать как перераспределение активности (энергии) с одного уровня регуляции на другой.
Таким образом, вопрос о парциальности свойств нервной системы продолжает оставаться актуальным и требует дальнейшего изучения.
Однако, как отмечал В. Д. Небылицын, «особенности организации мозговой рецептивной системы принципиально, по самой ее функции, таковы, что ее функциональные параметры – частные свойства – едва ли могут служить нейрофизиологическими “измерениями” личности как целого, со всеми общими аспектами ее организации и поведения» (1976, с. 21). Соответственно он выдвинул гипотезу о существовании и общих свойств нервной системы, которые связаны с выделенной им регулирующей системой мозга.
К этой системе В. Д. Небылицын отнес передние отделы неокортекса и взаимодействующие с ними образования старой и древней коры головного мозга, а также его стволовой части. Структуры, составляющие этот комплекс, он рассматривал в известном приближении как единую мозговую систему, выполняющую функции регуляции и управления всеми процессами, протекающими в организме – от низших биологических до самых высших психических (таких как потребности и мотивация, эмоции и темперамент, программирование движения, и действий, интеллектуальное планирование и оценка результатов и т. д.). В отличие от анализаторной, регулирующая система не имеет прямого отношения к переработке сенсорных раздражителей.
Важными являются связи свойств этой системы со многими общеличностными характеристиками – моторной активностью, интеллектуальной активностью и с особенностями установки личности, в то время как анализаторные (парциальные) свойства часто таких связей не обнаруживают.
Следует подчеркнуть, что В. Д. Небылицын, выдвигая понятие об общих свойствах нервной системы, рассматривал их не как чисто корковые, а как комплексные, отражающие функциональную систему, включающие в себя различные уровни центральной нервной системы: лобную кору, лимбический мозг, ретикулярную формацию. Можно считать, что под общими свойствами В. Д. Небылицын понимал, скорее всего, свойства интегративных систем мозга, осуществляющих произвольные акты человека. При этом он отмечал: свойства, обозначенные им как общие, строго говоря, тоже являются частными, поскольку представляют собой параметры пусть весьма обширного, но все же ограниченного анатомически и функционально комплекса структур головного мозга. Интерпретация этих свойств как общих оправдана только тем значением, которое имеют образования регуляторной системы для психической жизни и поведения всего организма. Во всяком случае, они, по мнению В. Д. Небылицына, дают лучшее приближение к функции «нервной системы в целом», чем парциальные свойства. Как полагал исследователь, представление об этих общих свойствах поможет понять, что же такое свойства нервной системы в целом, – категорию, остающуюся, по его мысли, на современном уровне наших психофизиологических знаний чисто гипотетической конструкцией.
Об этом высказывании В. Д. Небылицына (об относительности «общих» свойств) необходимо помнить, так как в более поздних работах других авторов содержатся высказывания, противоречащие положениям этого ученого.
Например, отличной от позиции В. Д. Небылицына точки зрения придерживается В. М. Русалов (1979). Он считает, что общие свойства – это индивидуально-устойчивые особенности целостной общемозговой интеграции нервных процессов, вовлеченных в целостную произвольную психическую деятельность, в то время как частные – особенности локальной интеграции. Автор полагает более правильным не увязывать строго общие свойства с деятельностью лобного отдела, а рассматривать их как особенности регуляции всего мозга. Свойства переднего мозга могут быть лишь аналогичными, но не идентичными свойствам всей нервной системы.
Со второй половины ХХ в. идея общемозговых свойств все больше замыкается на активации целого мозга и связанной с ней чувствительностью. Предполагается (В. С. Мерлин, 1973; И. М. Палей, 1966), что чувствительность прямо отражает «первичное» общее свойство. Согласно другой точке зрения, «первичным» и в значительной степени интегральным свойством является активированность покоя, а чувствительность – вторичная характеристика, зависящая от уровня активации в покое нервной системы (Е. П. Ильин, 1979).
Свойства, выявляемые с помощью методик, в которых используются произвольные двигательные реакции человека, обнаруживают постоянные связи с деятельностными и поведенческими характеристиками людей, а свойства, выявляемые в зрительном и слуховом анализаторах таких связей часто не демонстрируют или же показывают те, которые противоречат здравому смыслу. Следовательно, для практических целей прогнозирования склонностей, способностей, устойчивости к неблагоприятным состояниям и т. д. парциальные (анализаторные) свойства малопригодны.
Глава 5
Характеристика отдельных свойств нервной системы
5.1. Сила нервной системы
Понятие о свойстве силы нервной системы выдвинуто И. П. Павловым в 1922 г. При изучении условно-рефлекторной деятельности у животных было выявлено, что чем больше интенсивность раздражителя или чем чаще он применяется, тем больше ответная условно-рефлекторная реакция. Однако при достижении определенной интенсивности или частоты раздражения условно-рефлекторный ответ начинает снижаться. В целом эта зависимость была сформулирована как «закон силы» (рис. 5.1).
Было отмечено, что у животных этот закон проявляется по-разному: запредельное торможение, при котором начинается снижение условно-рефлекторного ответа, у одних наступает при меньшей интенсивности или частоте раздражения, чем у других. Первые были отнесены к «слабому типу» нервной системы, вторые – к «сильному типу». Возникли и два способа диагностики силы нервной системы: по максимальной интенсивности однократного раздражения, еще не приводящего к снижению условно-рефлекторной реакции (измерение силы через «верхний порог»), и по наибольшему числу раздражения, тоже еще не приводящему к снижению рефлекторного ответа (измерение силы через ее «выносливость»).
В лаборатории Б. М. Теплова была обнаружена большая чувствительность лиц со слабой нервной системой по сравнению с теми, у кого та оказалась сильной. Отсюда возник еще один способ измерения силы: через быстроту реагирования человека на сигналы разной интенсивности. Субъекты со слабой нервной системой из-за своей более высокой чувствительности реагируют на слабые и средние по силе сигналы быстрее, чем субъекты с сильной нервной системой. По сути, в этом случае сила нервной системы определяется по «нижнему порогу».
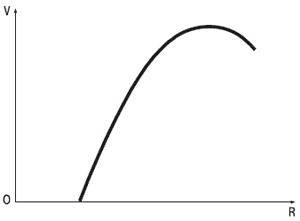
Рис. 5.1. Схема, показывающая проявление «закона силы». По вертикали – величина реакции; по горизонтали – сила разрушения.
В том же исследовательском коллективе сила нервной системы стала определяться и по уровню активации ЭЭГ. Однако этот метод технически сложен для массовых обследований.
До недавнего времени все эти способы измерения силы нервной системы не имели единого теоретического обоснования и поэтому рассматривались как независимые друг от друга, обнаруживающие различные проявления силы нервной системы, связанные, как казалось, с разными физиологическими механизмами. Поэтому оправданным было и требование изучать типологические проявления свойств сразу несколькими методиками, о чем говорилось в предыдущем параграфе. Однако возможно единое объяснение различных проявлений силы нервной системы (Е. П. Ильин, 1979), которое делает равноправными различные методики, с помощью которых устанавливается сила нервных процессов. Объединяющим фактором оказался уровень активации в покое (суждение о котором выносилось на основании уровня энерготрат в покое – рис. 5.2): у одних людей он выше, а у других ниже. Отсюда и различия в проявлении «закона силы».
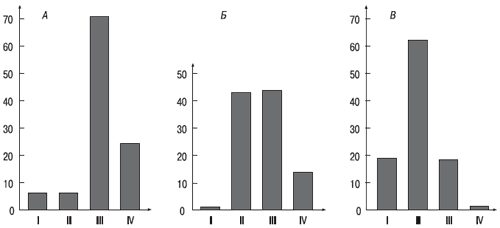
Рис. 5.2. Распределение испытуемых с различными энерготратами в покое (уровнем активации) в группах с различной силой нервной системы. По вертикали – число лиц, 5; по горизонтали – уровень энерготрат (ккал/кг/ч): I – от 0.50 до 0,99; II – от 1,00 до 1,50; III – от 1,51 до 2,00; IV – от 2, 01 и выше. А – лица с малой силой нервной системы; Б – лица со средней силой нервной системы; В – лица с большой силой нервной системы.
Сила нервной системы как реактивность. Для возникновения видимой ответной реакции (ощущение раздражителя или движение рукой) нужно, чтобы раздражитель превысил определенную (пороговую) величину или по крайней мере ее достиг. Это значит, что данный раздражитель вызывает такие физиологические и физико-химические изменения раздражаемого субстрата, которые достаточны для появления ощущения или ответной двигательной реакции. Следовательно, чтобы получить ответную реакцию, надо достичь порогового уровня активации нервной системы. Но в состоянии физиологического покоя последняя уже находится на определенном уровне активированности, правда, ниже порогового. У субъектов со слабой нервной системой уровень активации в покое выше (это следует из того, что в покое у них выше потребление кислорода и энерготраты на 1 кг веса тела); соответственно они ближе к пороговому уровню активации, с которого начинается реагирование (рис. 5.3), чем лица с сильной нервной системой. Для доведения этого уровня до порогового им, как следует из схемы, нужен меньший по интенсивности раздражитель. Субъектам же с сильной нервной системой, у которых уровень активации покоя ниже, требуется большая величина раздражителя, чтобы довести уровень активации до порогового. Этим и обусловлены различия между «слабыми» и «сильными» по нижнему порогу раздражения (r1 < r2).
При нарастании интенсивности одиночных раздражителей уровень активации (возбуждения) и величина (или быстрота, как при измерении времени реакции) реагирования повышаются. Однако субъекты со слабой нервной системой, начав реагировать раньше, чем лица с сильной нервной системой, раньше достигают и предельного уровня активации, при котором наблюдаются наибольшие и самые быстрые ответные реакции. После этого у них эффект реагирования снижается, в то время как у субъектов с сильной нервной системой он еще возрастает. Они достигают предела активации позже, при большей силе одиночного раздражителя (R1 < R2). Следовательно, и «верхний» порог у «слабых» меньший, чем у «сильных», т. е. запредельное торможение у первых наступает раньше, чем у вторых, при меньшей интенсивности достаточно сильного раздражителя (рис. 5.3).
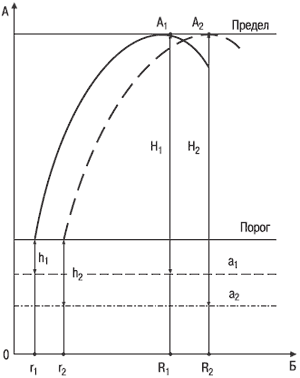
Рис. 5.3. Схема, показывающая различия в силе нервной системы в зависимости от интенсивности раздражителя. По вертикали – уровень активации: а1– в состоянии покоя у лиц со слабой нервной системой; а2– у лиц с сильной нервной системой; нижняя сплошная линия – пороговый уровень активации покоя, с которого начинается реакция на раздражитель; верхняя сплошная линия – предельный уровень реагирования (А1– для лиц со слабой нервной системой; А2– для лиц с сильной нервной системой). По горизонтали – интенсивность раздражителя: r1– нижний порог для лиц со слабой нервной системой, r2– то же для лиц с сильной нервной системой; R1 – верхний порог раздражителя для лиц со слабой нервной системой, R2– то же для лиц с сильной нервной системой; h1– величина дополнительной активации, необходимая для достижения порога реагирования лицами со слабой нервной системой, h2– то же для лиц с сильной нервной системой.
На выявление этих различий в реагировании людей на стимулы разной интенсивности направлена методика, разработанная В. Д. Небылицыным и получившая краткое название «наклон кривой» (рис. 5.4; описание методики см. в приложении).
В. Д. Небылицын выдвинул гипотезу, что диапазон между нижним (r) и верхним (R) порогами должен сохраняться от индивида к индивиду неизменным:
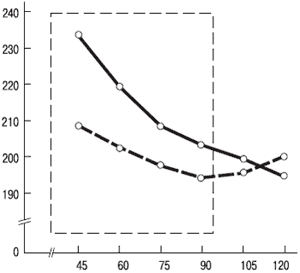
Рис. 5.4. Изменение времени реакции на различные по интенсивности звуковые сигналы у лиц с сильной и слабой нервной системой. По вертикали – время реакции, мс; по горизонтали – громкость звука, дБ. Сплошная линия – данные для лиц с сильной нервной системой; штрихпунктирная – для лиц со слабой нервной системой. Пунктиром обозначена зона слабых и средних интенсивностей звука, используемых в методике В. Д. Небылицына.
Из приведенной формулы следует, что и сильная и слабая нервная система должны выдерживать одну и ту же величину градиента (прироста) сверхпорогового раздражителя. Если принять абсолютный порог за нулевую точку отсчета величины физиологической силы раздражителя, то при увеличении его силы и сильная и слабая нервная система будут реагировать одинаково: увеличится в два раза сила раздражителя – во столько же раз возрастет и величина ответной реакции со стороны как сильной, так и слабой нервной системы.
Из этого должно также вытекать, что различий между последними при выравнивании физиологической силы раздражителя не будет; в обеих нервных системах запредельное торможение возникнет при одной и той же физиологической силе раздражителя. Это означает, что ход кривой реагирования на стимулы разной физиологической силы сильной и слабой нервной системы совпадет. Таким образом, согласно данной гипотезе В. Д. Небылицына, различия в силе нервной системы обнаруживаются потому, что используется физическая шкала интенсивности раздражителя, в которой одинаковая физическая величина последнего является разной физиологической силой для сильной и слабой нервной системы. Причиной этого, как теперь стало ясно, служит их различная фоновая активированность: чем она выше, тем большим по физиологической силе становится физический раздражитель.
Однако эта правдоподобная гипотеза В. Д. Небылицына остается пока не доказанной на практике. Более того, П. О. Макаров (1955) использовал разницу между верхним и нижним порогами в качестве показателя силы нервной системы: чем больше диапазон между порогами (который автором принят за энергетический потенциал), тем больше сила нервной системы. Но и эта гипотеза осталась непроверенной экспериментально.
Сила нервной системы как выносливость. Многократное повторное предъявление одинакового по силе раздражителя через короткие интервалы времени вызывает явление суммации, т. е. усиление рефлекторных реакций за счет роста фоновой активации, так как каждое предыдущее возбуждение оставляет после себя след, и поэтому каждая последующая реакция испытуемого начинается на более высоком функциональном уровне, чем предыдущая (заштрихованная область на рис. 5.5).
Было отмечено, что у животных этот закон проявляется по-разному: запредельное торможение, при котором начинается снижение условно-рефлекторного ответа, у одних наступает при меньшей интенсивности или частоте раздражения, чем у других. Первые были отнесены к «слабому типу» нервной системы, вторые – к «сильному типу». Возникли и два способа диагностики силы нервной системы: по максимальной интенсивности однократного раздражения, еще не приводящего к снижению условно-рефлекторной реакции (измерение силы через «верхний порог»), и по наибольшему числу раздражения, тоже еще не приводящему к снижению рефлекторного ответа (измерение силы через ее «выносливость»).
В лаборатории Б. М. Теплова была обнаружена большая чувствительность лиц со слабой нервной системой по сравнению с теми, у кого та оказалась сильной. Отсюда возник еще один способ измерения силы: через быстроту реагирования человека на сигналы разной интенсивности. Субъекты со слабой нервной системой из-за своей более высокой чувствительности реагируют на слабые и средние по силе сигналы быстрее, чем субъекты с сильной нервной системой. По сути, в этом случае сила нервной системы определяется по «нижнему порогу».
Рис. 5.1. Схема, показывающая проявление «закона силы». По вертикали – величина реакции; по горизонтали – сила разрушения.
В том же исследовательском коллективе сила нервной системы стала определяться и по уровню активации ЭЭГ. Однако этот метод технически сложен для массовых обследований.
До недавнего времени все эти способы измерения силы нервной системы не имели единого теоретического обоснования и поэтому рассматривались как независимые друг от друга, обнаруживающие различные проявления силы нервной системы, связанные, как казалось, с разными физиологическими механизмами. Поэтому оправданным было и требование изучать типологические проявления свойств сразу несколькими методиками, о чем говорилось в предыдущем параграфе. Однако возможно единое объяснение различных проявлений силы нервной системы (Е. П. Ильин, 1979), которое делает равноправными различные методики, с помощью которых устанавливается сила нервных процессов. Объединяющим фактором оказался уровень активации в покое (суждение о котором выносилось на основании уровня энерготрат в покое – рис. 5.2): у одних людей он выше, а у других ниже. Отсюда и различия в проявлении «закона силы».
Рис. 5.2. Распределение испытуемых с различными энерготратами в покое (уровнем активации) в группах с различной силой нервной системы. По вертикали – число лиц, 5; по горизонтали – уровень энерготрат (ккал/кг/ч): I – от 0.50 до 0,99; II – от 1,00 до 1,50; III – от 1,51 до 2,00; IV – от 2, 01 и выше. А – лица с малой силой нервной системы; Б – лица со средней силой нервной системы; В – лица с большой силой нервной системы.
Сила нервной системы как реактивность. Для возникновения видимой ответной реакции (ощущение раздражителя или движение рукой) нужно, чтобы раздражитель превысил определенную (пороговую) величину или по крайней мере ее достиг. Это значит, что данный раздражитель вызывает такие физиологические и физико-химические изменения раздражаемого субстрата, которые достаточны для появления ощущения или ответной двигательной реакции. Следовательно, чтобы получить ответную реакцию, надо достичь порогового уровня активации нервной системы. Но в состоянии физиологического покоя последняя уже находится на определенном уровне активированности, правда, ниже порогового. У субъектов со слабой нервной системой уровень активации в покое выше (это следует из того, что в покое у них выше потребление кислорода и энерготраты на 1 кг веса тела); соответственно они ближе к пороговому уровню активации, с которого начинается реагирование (рис. 5.3), чем лица с сильной нервной системой. Для доведения этого уровня до порогового им, как следует из схемы, нужен меньший по интенсивности раздражитель. Субъектам же с сильной нервной системой, у которых уровень активации покоя ниже, требуется большая величина раздражителя, чтобы довести уровень активации до порогового. Этим и обусловлены различия между «слабыми» и «сильными» по нижнему порогу раздражения (r1 < r2).
При нарастании интенсивности одиночных раздражителей уровень активации (возбуждения) и величина (или быстрота, как при измерении времени реакции) реагирования повышаются. Однако субъекты со слабой нервной системой, начав реагировать раньше, чем лица с сильной нервной системой, раньше достигают и предельного уровня активации, при котором наблюдаются наибольшие и самые быстрые ответные реакции. После этого у них эффект реагирования снижается, в то время как у субъектов с сильной нервной системой он еще возрастает. Они достигают предела активации позже, при большей силе одиночного раздражителя (R1 < R2). Следовательно, и «верхний» порог у «слабых» меньший, чем у «сильных», т. е. запредельное торможение у первых наступает раньше, чем у вторых, при меньшей интенсивности достаточно сильного раздражителя (рис. 5.3).
Рис. 5.3. Схема, показывающая различия в силе нервной системы в зависимости от интенсивности раздражителя. По вертикали – уровень активации: а1– в состоянии покоя у лиц со слабой нервной системой; а2– у лиц с сильной нервной системой; нижняя сплошная линия – пороговый уровень активации покоя, с которого начинается реакция на раздражитель; верхняя сплошная линия – предельный уровень реагирования (А1– для лиц со слабой нервной системой; А2– для лиц с сильной нервной системой). По горизонтали – интенсивность раздражителя: r1– нижний порог для лиц со слабой нервной системой, r2– то же для лиц с сильной нервной системой; R1 – верхний порог раздражителя для лиц со слабой нервной системой, R2– то же для лиц с сильной нервной системой; h1– величина дополнительной активации, необходимая для достижения порога реагирования лицами со слабой нервной системой, h2– то же для лиц с сильной нервной системой.
На выявление этих различий в реагировании людей на стимулы разной интенсивности направлена методика, разработанная В. Д. Небылицыным и получившая краткое название «наклон кривой» (рис. 5.4; описание методики см. в приложении).
В. Д. Небылицын выдвинул гипотезу, что диапазон между нижним (r) и верхним (R) порогами должен сохраняться от индивида к индивиду неизменным:
R: r= сопst.Автор исходил из того факта, что оба этих порога у лиц с сильной нервной системой выше, чем у лиц со слабой нервной системой.
Рис. 5.4. Изменение времени реакции на различные по интенсивности звуковые сигналы у лиц с сильной и слабой нервной системой. По вертикали – время реакции, мс; по горизонтали – громкость звука, дБ. Сплошная линия – данные для лиц с сильной нервной системой; штрихпунктирная – для лиц со слабой нервной системой. Пунктиром обозначена зона слабых и средних интенсивностей звука, используемых в методике В. Д. Небылицына.
Из приведенной формулы следует, что и сильная и слабая нервная система должны выдерживать одну и ту же величину градиента (прироста) сверхпорогового раздражителя. Если принять абсолютный порог за нулевую точку отсчета величины физиологической силы раздражителя, то при увеличении его силы и сильная и слабая нервная система будут реагировать одинаково: увеличится в два раза сила раздражителя – во столько же раз возрастет и величина ответной реакции со стороны как сильной, так и слабой нервной системы.
Из этого должно также вытекать, что различий между последними при выравнивании физиологической силы раздражителя не будет; в обеих нервных системах запредельное торможение возникнет при одной и той же физиологической силе раздражителя. Это означает, что ход кривой реагирования на стимулы разной физиологической силы сильной и слабой нервной системы совпадет. Таким образом, согласно данной гипотезе В. Д. Небылицына, различия в силе нервной системы обнаруживаются потому, что используется физическая шкала интенсивности раздражителя, в которой одинаковая физическая величина последнего является разной физиологической силой для сильной и слабой нервной системы. Причиной этого, как теперь стало ясно, служит их различная фоновая активированность: чем она выше, тем большим по физиологической силе становится физический раздражитель.
Однако эта правдоподобная гипотеза В. Д. Небылицына остается пока не доказанной на практике. Более того, П. О. Макаров (1955) использовал разницу между верхним и нижним порогами в качестве показателя силы нервной системы: чем больше диапазон между порогами (который автором принят за энергетический потенциал), тем больше сила нервной системы. Но и эта гипотеза осталась непроверенной экспериментально.
Сила нервной системы как выносливость. Многократное повторное предъявление одинакового по силе раздражителя через короткие интервалы времени вызывает явление суммации, т. е. усиление рефлекторных реакций за счет роста фоновой активации, так как каждое предыдущее возбуждение оставляет после себя след, и поэтому каждая последующая реакция испытуемого начинается на более высоком функциональном уровне, чем предыдущая (заштрихованная область на рис. 5.5).