Страница:
Часто одни и те же виды занимают разные экологические ниши в разных местообитаниях. Луговой клещ в северной лесостепи Западной Сибири предпочитает опушки колков и поляны, а в южной лесостепи – глубину колков. В тех местах, где обитают три вида пестрых дятлов – большой, средний и малый – первый кормится на стволах, второй на ветвях, а третий в кронах, но если в каком‑либо местообитании встречается только один из этих видов, он кормится на всем дереве. Там, где норка не встречается с выдрой, она ест и рыбу, и моллюсков, и раков, и лягушек. Но там, где есть выдра, норка почти не ест рыбу, а выдра не ест моллюсков и лягушек. Серая крыса при совместном обитании с черной вытесняет ее на чердаки, а сама занимает подвалы. Но там, где серой крысы нет, черная селится и на чердаках, и в подвалах. Следовательно, при совместном обитании близких видов экологические ниши суживаются, а при раздельном – расширяются.
Наиболее широки экологические ниши в местах, где мало видов и они по преимуществу эврибионты – в тундрах, высокогорьях. Наиболее узкие экологические ниши там, где видов много и большинство из них стенобионтны – в тропических лесах, на коралловых рифах.
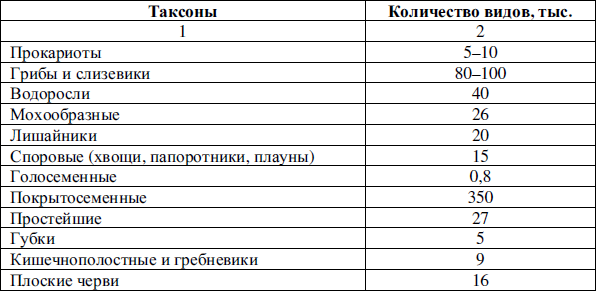
В настоящее время на земле обитает около трех миллионов видов различных живых существ (таблица 3), количество же экологических ниш неизмеримо меньше.
Таблица 3
Количество видов живых организмов в разных таксонах

Поэтому большое число видов должно укладываться в это небольшое количество жизненных форм. Причем сходные жизненные формы могут объединять виды, далеко не родственные друг другу и обитающие в весьма отдаленных друг от друга местообитаниях. По образному выражению Ф. Дарлингтона, на юге Африки, Австралии и Южной Америки млекопитающие объедают кусты, но и кусты, и млекопитающие на этих континентах разные.
Классификация жизненных форм растений предложена П. Раункиером. За основу им взято положение ростовых почек и способы их защиты.
1 тип. Фанерофиты. Ростовые почки находятся на высоте более 50 см над землей и ничем не защищены. Здесь выделяются классы: деревья, кустарники, стеблевые суккуленты, лианы и эпифиты. В каждом из классов выделяются более детальные подразделения. Например, деревья подразделяются на деревья с обычной кроной, пирамидальной кроной (ели, пихты и пр.) и с пальмовидной кроной.
2 тип. Хамефиты. Ростовые почки находятся на высоте ниже 50 см над землей и тоже ничем не защищены. Выделяются классы кустарничков (брусника, клюква и пр.) и стелющихся (карликовая березка, кедровый стланик). При неблагоприятных условиях ростовые почки защищены снегом или растительным опадом.
3 тип. Гемикриптофиты. Ростовая почка находится на уровне почвы и защищена частями самого растения: прикорневой розеткой листьев, чешуйками, неотпавшими прошлогодними листьями и пр.
4 тип. Криптофиты. Ростовая почка находится в подземных частях растения – в корневищах, клубнях, луковицах. Подразделяется на классы геофитов (почки под землей), гидрофитов (почки под водой) и гелофитов (почки в иле или болотистом грунте).
5 тип. Терофиты. Однолетние растения, у которых в неблагоприятный период растение погибает, а сохраняются до следующего сезона вегетации лишь семена (большинство злаков).
В отношении животных такой четкой классификации не выработано. Д. М. Кашкаров предложил классификацию жизненных форм животных на основании способов передвижения и типа среды обитания.
1 тип. Плавающие, или водные. Делятся на классы: чисто водные (планктон, нектон и бентос) и полуводные (плавающие неныряющие – утки, гуси; плавающие ныряющие – гагары, пингвины; и собирающие в воде корм – цапли, кулики).
2 тип. Роющие. Делятся на классы: абсолютные землерои (дождевые черви, кроты) и относительные землерои (личинки многих насекомых, имаго которых живут не в земле – майских жуков, цикад).
3 тип. Наземные. Делятся на классы: не делающие нор (бегающие – копытные, хищниые, страусы; прыгающие – кенгуру; ползающие – змеи), делающих норы (бегающие – суслики, полевки; прыгающие – тушканчики, долгоноги; ползающие – некоторые змеи и безногие ящерицы) и скалолазы (некоторые ящерицы и птицы).
4 тип. Древесные. Делятся на классы: не сходящие с деревьев (обитатели крон – обезьяны, ленивцы; обитатели дупел – различные членистоногие, некоторые рептилии; ствололазы – различные членистоногие) и сходящие с деревьев (некоторые обезьяны, белки, куньи).
5 тип. Воздушные. Делятся на классы: добывающие пищу в воздухе (стрижи, ласточки, летучие мыши) и высматривающие пищу с воздуха (большинство птиц и насекомых).
Эта классификация не получила всеобщего признания, т. к. зачастую трудно найти в ней место какому‑либо виду. Так, суслик отнесен к бегающим, делающим норы, но его можно отнести и к относительным землероям. Чайку можно относить к воздушным, высматривающим пищу с воздуха, но можно отнести и к полуводным, плавающим и не ныряющим и мн. др.
В настоящее время существуют многочисленные классификации жизненных форм животных отдельных систематических категорий: рыб, птиц, насекомых – и даже по более мелким таксонам: грызунов, прямокрылых и пр.
К сходным жизненным формам зачастую относятся виды, далеко отстоящие друг от друга по систематическому положению. Скат и камбала относятся к одной жизненной форме, но скат – хрящевая рыба, а камбала – костистая. Тушканчик и кенгуру также представляют сходную жизненную форму, но кенгуру – сумчатое, а тушканчик – плацентарное млекопитающее. Сходные жизненные формы могут объединять даже представителей разных классов (дельфин – млекопитающее и акула – хрящевая рыба) и даже типов (крот – хордовое, медведка – членистоногое).
Жизненная форма не всегда совпадает с экологической нишей – овца и кенгуру относятся к разным жизненным формам, но занимают одну экологическую нишу (почему овца и вытесняет кенгуру из его местообитаний).
В любой экосистеме имеются разнообразные жизненные формы. В совокупности они максимально используют ее ресурсы. В этом отношении сходны даже такие различные экосистемы, как тундра и тропический лес: в том и в другом есть столько жизненных форм, чтобы заполнить все экологические ниши (другое дело, что количество ниш несоизмеримо). Например, среди степных грызунов мы встречаем роющих неглубокие норы и питающихся зелеными частями растений (полевки), роющих глубокие норы и питающиеся преимущественно семенами (суслики), роющих подземные галереи и питающиеся подземными частями растений (слепыши, цокоры), прыгающих (тушканчики), лазающих в высоких травах (мышь-малютка). В тундре же мы видим только леммингов, занимающих очень широкую экологическую нишу, да некоторых полевок, занимающих весьма специфические местообитания – чаще всего поймы рек. Чем богаче ресурсами экосистема, чем больше в ней экологических ниш, тем больше жизненных форм она содержит.
При изменении условий, при появлении новых экологических ниш или освобождении уже имеющихся, естественный отбор создает новые формы, стремящиеся занять освободившиеся или вновь возникшие ниши. Это творческая, формообразующая роль отбора. Если же природные условия стабильны, долгое время не меняются, все экологические ниши заполнены, то отбор отсекает все отклонения от средней нормы, возникающие в результате мутаций. Это стабилизирующая роль отбора. В итоге и виды, и биоценозы находятся в динамическом равновесии с окружающей средой, в состоянии гомеостаза. Это характерно в первую очередь для климаксовых биоценозов.
В настоящее время на Земле трудно найти какое‑либо сообщество, имеющее незаполненные экологические ниши. Некоторое исключение составляют океанические острова, особенно те из них, которые возникли сравнительно недавно (вулканические острова, коралловые атоллы). Заселяемые в известной степени случайно, они какое‑то время имеют незаполненные ниши, но именно наличие таких ниш стимулирует процесс видообразования: занятие свободной ниши, изначальное приспособление к непривычной нише влечет за собой его изменения – сначала экологические, потом морфофизиологические и наконец – закрепление новых признаков на генетическом уровне – образование нового вида.
Глава 10
Часть 2
Глава 11
Наиболее широки экологические ниши в местах, где мало видов и они по преимуществу эврибионты – в тундрах, высокогорьях. Наиболее узкие экологические ниши там, где видов много и большинство из них стенобионтны – в тропических лесах, на коралловых рифах.
В настоящее время на земле обитает около трех миллионов видов различных живых существ (таблица 3), количество же экологических ниш неизмеримо меньше.
Таблица 3
Количество видов живых организмов в разных таксонах
Поэтому большое число видов должно укладываться в это небольшое количество жизненных форм. Причем сходные жизненные формы могут объединять виды, далеко не родственные друг другу и обитающие в весьма отдаленных друг от друга местообитаниях. По образному выражению Ф. Дарлингтона, на юге Африки, Австралии и Южной Америки млекопитающие объедают кусты, но и кусты, и млекопитающие на этих континентах разные.
Классификация жизненных форм растений предложена П. Раункиером. За основу им взято положение ростовых почек и способы их защиты.
1 тип. Фанерофиты. Ростовые почки находятся на высоте более 50 см над землей и ничем не защищены. Здесь выделяются классы: деревья, кустарники, стеблевые суккуленты, лианы и эпифиты. В каждом из классов выделяются более детальные подразделения. Например, деревья подразделяются на деревья с обычной кроной, пирамидальной кроной (ели, пихты и пр.) и с пальмовидной кроной.
2 тип. Хамефиты. Ростовые почки находятся на высоте ниже 50 см над землей и тоже ничем не защищены. Выделяются классы кустарничков (брусника, клюква и пр.) и стелющихся (карликовая березка, кедровый стланик). При неблагоприятных условиях ростовые почки защищены снегом или растительным опадом.
3 тип. Гемикриптофиты. Ростовая почка находится на уровне почвы и защищена частями самого растения: прикорневой розеткой листьев, чешуйками, неотпавшими прошлогодними листьями и пр.
4 тип. Криптофиты. Ростовая почка находится в подземных частях растения – в корневищах, клубнях, луковицах. Подразделяется на классы геофитов (почки под землей), гидрофитов (почки под водой) и гелофитов (почки в иле или болотистом грунте).
5 тип. Терофиты. Однолетние растения, у которых в неблагоприятный период растение погибает, а сохраняются до следующего сезона вегетации лишь семена (большинство злаков).
В отношении животных такой четкой классификации не выработано. Д. М. Кашкаров предложил классификацию жизненных форм животных на основании способов передвижения и типа среды обитания.
1 тип. Плавающие, или водные. Делятся на классы: чисто водные (планктон, нектон и бентос) и полуводные (плавающие неныряющие – утки, гуси; плавающие ныряющие – гагары, пингвины; и собирающие в воде корм – цапли, кулики).
2 тип. Роющие. Делятся на классы: абсолютные землерои (дождевые черви, кроты) и относительные землерои (личинки многих насекомых, имаго которых живут не в земле – майских жуков, цикад).
3 тип. Наземные. Делятся на классы: не делающие нор (бегающие – копытные, хищниые, страусы; прыгающие – кенгуру; ползающие – змеи), делающих норы (бегающие – суслики, полевки; прыгающие – тушканчики, долгоноги; ползающие – некоторые змеи и безногие ящерицы) и скалолазы (некоторые ящерицы и птицы).
4 тип. Древесные. Делятся на классы: не сходящие с деревьев (обитатели крон – обезьяны, ленивцы; обитатели дупел – различные членистоногие, некоторые рептилии; ствололазы – различные членистоногие) и сходящие с деревьев (некоторые обезьяны, белки, куньи).
5 тип. Воздушные. Делятся на классы: добывающие пищу в воздухе (стрижи, ласточки, летучие мыши) и высматривающие пищу с воздуха (большинство птиц и насекомых).
Эта классификация не получила всеобщего признания, т. к. зачастую трудно найти в ней место какому‑либо виду. Так, суслик отнесен к бегающим, делающим норы, но его можно отнести и к относительным землероям. Чайку можно относить к воздушным, высматривающим пищу с воздуха, но можно отнести и к полуводным, плавающим и не ныряющим и мн. др.
В настоящее время существуют многочисленные классификации жизненных форм животных отдельных систематических категорий: рыб, птиц, насекомых – и даже по более мелким таксонам: грызунов, прямокрылых и пр.
К сходным жизненным формам зачастую относятся виды, далеко отстоящие друг от друга по систематическому положению. Скат и камбала относятся к одной жизненной форме, но скат – хрящевая рыба, а камбала – костистая. Тушканчик и кенгуру также представляют сходную жизненную форму, но кенгуру – сумчатое, а тушканчик – плацентарное млекопитающее. Сходные жизненные формы могут объединять даже представителей разных классов (дельфин – млекопитающее и акула – хрящевая рыба) и даже типов (крот – хордовое, медведка – членистоногое).
Жизненная форма не всегда совпадает с экологической нишей – овца и кенгуру относятся к разным жизненным формам, но занимают одну экологическую нишу (почему овца и вытесняет кенгуру из его местообитаний).
В любой экосистеме имеются разнообразные жизненные формы. В совокупности они максимально используют ее ресурсы. В этом отношении сходны даже такие различные экосистемы, как тундра и тропический лес: в том и в другом есть столько жизненных форм, чтобы заполнить все экологические ниши (другое дело, что количество ниш несоизмеримо). Например, среди степных грызунов мы встречаем роющих неглубокие норы и питающихся зелеными частями растений (полевки), роющих глубокие норы и питающиеся преимущественно семенами (суслики), роющих подземные галереи и питающиеся подземными частями растений (слепыши, цокоры), прыгающих (тушканчики), лазающих в высоких травах (мышь-малютка). В тундре же мы видим только леммингов, занимающих очень широкую экологическую нишу, да некоторых полевок, занимающих весьма специфические местообитания – чаще всего поймы рек. Чем богаче ресурсами экосистема, чем больше в ней экологических ниш, тем больше жизненных форм она содержит.
При изменении условий, при появлении новых экологических ниш или освобождении уже имеющихся, естественный отбор создает новые формы, стремящиеся занять освободившиеся или вновь возникшие ниши. Это творческая, формообразующая роль отбора. Если же природные условия стабильны, долгое время не меняются, все экологические ниши заполнены, то отбор отсекает все отклонения от средней нормы, возникающие в результате мутаций. Это стабилизирующая роль отбора. В итоге и виды, и биоценозы находятся в динамическом равновесии с окружающей средой, в состоянии гомеостаза. Это характерно в первую очередь для климаксовых биоценозов.
В настоящее время на Земле трудно найти какое‑либо сообщество, имеющее незаполненные экологические ниши. Некоторое исключение составляют океанические острова, особенно те из них, которые возникли сравнительно недавно (вулканические острова, коралловые атоллы). Заселяемые в известной степени случайно, они какое‑то время имеют незаполненные ниши, но именно наличие таких ниш стимулирует процесс видообразования: занятие свободной ниши, изначальное приспособление к непривычной нише влечет за собой его изменения – сначала экологические, потом морфофизиологические и наконец – закрепление новых признаков на генетическом уровне – образование нового вида.
Глава 10
Биогеографические царства
Наличие крупных преград или, наоборот, крупных соединений способствует обособлению биогеографических царств – наиболее крупных подразделений биосферы, имеющих общее происхождение, наличие наиболее характерных видов-индикаторов и резко отличающихся от соседних с ними других биогеографических царств. На следующих примерах можно показать, как формировались биоты этих царств.
Австралия отделилась от других материковых плит Гондваны около 60 млн лет назад и по каким‑то причинам не имела на своей территории плацентарных млекопитающих. Поэтому эволюция сумчатых шла здесь без пресса со стороны сильных конкурентов и привела к исключительному разнообразию жизненных форм, заполнивших все экологические ниши и давшие жизненные формы, конвергентно сходные с аналогичными жизненными формами плацентарных. На других же континентах сумчатые подвергались сильному давлению со стороны более конкурентоспособных плацентарных. Есть теория, что сумчатые возникли в Гондване, а плацентарные в Лавразии. Подтверждение тому мы видим в богатой фауне ископаемых сумчатых в Южной Америке, где произошло своеобразное «разделение труда» – плацентарные были в основном фитофагами, а сумчатые – хищниками. После соединения обеих Америк континентальным мостом сумчатые хищники не выдержали конкуренции со стороны более жизнеспособных плацентарных хищников и вымерли. Сейчас в Южной Америке есть несколько видов мелких насекомоядных сумчатых, а в Северной Америке – один вид, явно проникший сюда с юга. В Евразии нет современных сумчатых, а ископаемые представлены немногочисленными примитивными многорезцовыми, которые, вероятно, проникли сюда еще в Мезозое, но не выдержали конкуренции с бурно развивающимися плацентарными.
Неоднократные соединения Евразии с Северной Америкой через Берингию приводили к частому обмену мигрантами и к высокой общности населения этих двух континентов. Было минимум три соединения. Первое происходило в ту эпоху, когда Берингия была занята субтропическими лесами и степями (саваннами). Второе соединение происходило в период развития в Берингии таежных и, возможно, в ее южной части хвойно-широколиственных лесов. Третье соединение происходило в период, когда Берингия была покрыта тундрой или тундростепью. Возможно, это было уже не материковое соединение, а ледник, покрытый сверху лессовыми надувами, на которых и формировался растительный покров. Поэтому давно отделившиеся от общих предков виды, населяющие субтропики и степи обоих континентов, длительное время эволюционировали самостоятельно и различаются существенно (на уровне родов и семейств). Обитатели лесов разделились сравнительно недавно, поэтому их эволюционное расхождение не так велико – на уровне близких видов одного рода, подвидов одного вида, даже общих видов. Тундровые обитатели разделились совсем недавно, поэтому здесь обитают в основном те же виды, иногда подвиды одного вида, реже близкие виды одного рода. В то же время длительная изоляция между Северной и Южной Америкой привела к тому, что здесь крайне мало общих видов, в основном за счет миграций уже в Голоцене. Панамский перешеек не очень способствует миграциям, т. к. за ним находится непреодолимая для многих видов полоса пустынь. Больше мигрантов в обе стороны проникали через Карибский мост, существовавший в эпоху наибольшего оледенения и в связи с этим обмеления морей. Поэтому, например, Флорида имеет больше сходства с Южной Америкой, чем Мексика.
Раннее разделение Южной Америки от Африки вызвало несходство их современных биот, несмотря на сходство климатов. В то же время соединение Африки с Южной Азией, более влажный, чем теперь, климат в зоне соединения и отсутствие рифтовых разломов (нынешние Красное море и Персидский залив) способствовали интенсивному обмену флорой и фауной между ними. Возникновение упомянутых рифтов и опустынивание Северной Африки, Аравии и части Ближнего Востока привело к разделу этих биот, но, так как это произошло в сравнительно недавнее время, биоты их еще имеют много общего. Отрезанные от тропиков пустынями Северная Африка и Ближний Восток подверглись сильному влиянию биоты умеренного пояса Евразии, представители которой вытеснили остатки тропических растений и животных. Этому способствовало то, что во время наиболее сильных оледенений существовал континентальный мост от Апеннинского полуострова через Сицилию и Мальту, и отсутствие Гибралтарского пролива.
В настоящее время существует некоторая разница в выделении флористических и фаунистических царств, что затрудняет создание общеупотребительной системы биогеографических царств.
Ботаники выделяют шесть флористических царств, подразделяемых на ряд областей (по A. Л. Тахтаджяну).
1. Голарктическое царство. Занимает большую часть Евразии, кроме самого юга и юго-востока и юго-запада Аравии, почти всю Северную Америку, кроме самого юга Мексики и Флориды. Делится на области: циркумбореальную (зоны тундры, лесотундры, вся лесная и степная зоны Евразии, тундра, лесотундра и тайга Северной Америки), восточноазиатскую (хвойно-широколиственные леса и субтропики Японии, Кореи, Восточного Китая, Российское Приморье и Сахалин), Аппалачско-североамериканскую (от Атлантического океана до Скалистых гор и от Великих Озер до Мексиканского залива), область Скалистых гор, Макаронрезийскую (Канарские, Азорские о-ва и о. Мадейра), Средиземноморскую, Ирано-Туранскую (от Ливанских гор и Закавказья до среднеазиатских пустынно и гор, до Гоби и Тибета включительно), и Мадреанскую (пустыни и субтропики, юго-запад США и Мексики).
2. Палеотропическое царство. Занимает большую часть Африки к югу от Сахары (кроме самого юга), Мадагаскар, южную и юго-восточную Азию, Зондские и Филиппинские острова и большую часть островов Тихого океана, включая Гавайские. Делится на области: Гвинео– Конголезскую (побережье Гвинейского залива и бассейн р. Конго), Судано-Замбезийскую (почти вся Африка, кроме Гвиано-Конголезской области, Сахары и крайнего юга), Карру-Намибийская (крайний юг-запад Африки), островов св. Елены и Вознесения (упомянутые острова в Атлантическом океане), Мадагаскарскую (о. Мадагаскар с прилегающими островами), Индийскую (п-ов Индостан), Индокитайскую (п-ов Индокитай, южный Китай и северная часть Малаккского п-ва), Малезийскую (южная часть Малаккского п-ва, острова Филиппинские и Зондские, включая Новую Гвинею с прилегающими островами), Фиджийскую (о-ва Фиджи и прилегающие), Гавайскую (Гавайские о-ва), Полинезийскую (острова между Фиджи и Гавайскими), Новокаледонскую (о. Новая Каледония и прилегающие).
3. Неотропическое царство. Занимает Флориду, Панамский перешеек, острова Карибского моря и Южную Америку на юг до устья Ла-Платы. Делится на области: Карибскую (острова Карибского моря, Панамский перешеек, Флорида и крайний северо-запад Южной Америки), Амазонскую (большая часть бассейнов Амазонки и Ориноко), Гвианского нагорья (небольшой участок на северо-западе Амазонской области), Бразильскую (к югу от Амазонской до устья Ла-Платы и до побережья Атлантического океана) и Андийскую (горы Анды на юг немного южнее южного тропика).
4. Капское царство. Самый юг Африки, между Драконовыми горами и океаном.
5. Австралийское царство. Континент Австралии и остров Тасмания. Делится на области: Северо-Восточную (северное и восточное побережья Австралии и о. Тасмания), Юго-Западную (крайний юго– запад континента) и Центральную (большая часть Австралии, центр континента).
6. Голантарктическое царство. Делится на области: Островов Хуан-Фернандес (упомянутые острова), Чилийско-Патагонскую (Южная Америка к югу от устья Ла-Платы), Новозеландскую (Новая Зеландия) и субантарктических островов (мелкие острова по периферии Антарктиды).
Зоологи также выделяют шесть зоогеографических царств (по В. Г. Гептнеру).
6.1. Голарктическое царство. Включает всю северную Америку (с Флоридой), всю Евразию кроме крайнего юга и юго-востока и север Африки. Делится на области: Арктическую (тундры и лесотундры обоих континентов и острова Северного Ледовитого океана), Канадскую (большая часть Северной Америки кроме южной части континента), Европейско-Сибирскую (лесная и степная зоны Евразии), Средиземноморскую (побережье Средиземного моря, Сахара, Аравия, Кавказ, Ближний и Средний Восток до гор Средней Азии, на восток до Индии), Центральноазиатскую (пустыни и горы Средней Азии, Гоби и Тибет), Маньчжурско-Китайскую (Япония, Корея, Восточный Китай, российское Приморье, Сахалин), Сонорскую (южная часть Северной Америки).
6.2. Неотропическое царство. Острова Карибского моря, Панамский перешеек и вся Южная Америка. Делится на области: Центральноамериканскую (Панамский перешеек), Антильскую (острова Карибского моря), Бразильскую (северная часть материка, бассейны Амазонки и Ориноко, до Атлантического океана), Чилийскую (Анды и южная часть континента, к югу от устья Ла-Платы).
6.3. Эфиопское царство. Африка к югу от Сахары и Мадагаскар. Делится на области: Восточноафриканскую (побережье Гвинейского залива и бассейн р. Конго), Западноафриканскую (остальная часть Африки кроме юга), Южноафриканскую (юг континента), Мадагаскарскую (Мадагаскар).
6.4. Восточное (Индо-Малайсоке) царство. Индостан и Индокитай, Филиппинские и Зондские острова (до Новой Гвинеи). Делится на области: Индийскую (полуостров Индостан и Цейлон), Индомалайскую (Индокитай, южный Китай, Филиппинские и Зондские острова до Новой Гвинеи).
6.5. Австралазийское царство. Включает Австралию и все острова Тихого океана вплоть до Гавайских. Делится на области: Австралийскую (вся Австралия, кроме самого севера), Папуасскую (север Австралии, Новая Гвинея с прилегающими островами), Новозеландскую (Новая Зеландия), Полинезийскую (острова в Тихом океане, кроме Гавайских), Гавайскую (Гавайские острова).
6.6. Антарктическое царство. Мелкие острова вдоль побережья Антарктиды.
Иногда Голарктическое царство делят на два подцарства: Heap ктику (Северная Америка) и Палеарктику (Евразия). Также иногда выделяют лишь четыре царства: Арктогею (Голарктическое царство), Неогею (Неотропическое царство), Палеогею (Эфиопское и Восточное царства, делят на три области: Эфиопскую, Мадагаскарскую и Восточную). Нотогея (Австралазийское царство). Антарктическая область не входит по этой классификации, ни в одно из царств.
Из изложенного видно, что довольно трудно совместить зоогеографическое и флористическое районирование в единое биогеографическое. По А. Г. Воронову, Н. Н. Дроздову и В. В. Второву, выделяется восемь биогеографических царств: Голарктическое, Неотропическое, Эфиопское, Восточное, Австралийское, Антарктическое, Мадагаскарское и Капское. В основном принято зоогеографическое деление, только в последнем случае приоритет отдан флористическому.
Биогеографическое разделение Мирового Океана не столь четко. Флористы выделяют здесь три царства: Бореальное (к северу от северного тропика), Тропическое (от северного тропика до южного) и Южное (к югу от южного тропика). Зоогеографы выделяют пять царств: Арктическое, Атлантическое, Тихоокеанское, Тропическое (Индийский океан и тропические зоны Тихого и Атлантического океанов) и Антарктическое (южные части всех трех океанов).
Австралия отделилась от других материковых плит Гондваны около 60 млн лет назад и по каким‑то причинам не имела на своей территории плацентарных млекопитающих. Поэтому эволюция сумчатых шла здесь без пресса со стороны сильных конкурентов и привела к исключительному разнообразию жизненных форм, заполнивших все экологические ниши и давшие жизненные формы, конвергентно сходные с аналогичными жизненными формами плацентарных. На других же континентах сумчатые подвергались сильному давлению со стороны более конкурентоспособных плацентарных. Есть теория, что сумчатые возникли в Гондване, а плацентарные в Лавразии. Подтверждение тому мы видим в богатой фауне ископаемых сумчатых в Южной Америке, где произошло своеобразное «разделение труда» – плацентарные были в основном фитофагами, а сумчатые – хищниками. После соединения обеих Америк континентальным мостом сумчатые хищники не выдержали конкуренции со стороны более жизнеспособных плацентарных хищников и вымерли. Сейчас в Южной Америке есть несколько видов мелких насекомоядных сумчатых, а в Северной Америке – один вид, явно проникший сюда с юга. В Евразии нет современных сумчатых, а ископаемые представлены немногочисленными примитивными многорезцовыми, которые, вероятно, проникли сюда еще в Мезозое, но не выдержали конкуренции с бурно развивающимися плацентарными.
Неоднократные соединения Евразии с Северной Америкой через Берингию приводили к частому обмену мигрантами и к высокой общности населения этих двух континентов. Было минимум три соединения. Первое происходило в ту эпоху, когда Берингия была занята субтропическими лесами и степями (саваннами). Второе соединение происходило в период развития в Берингии таежных и, возможно, в ее южной части хвойно-широколиственных лесов. Третье соединение происходило в период, когда Берингия была покрыта тундрой или тундростепью. Возможно, это было уже не материковое соединение, а ледник, покрытый сверху лессовыми надувами, на которых и формировался растительный покров. Поэтому давно отделившиеся от общих предков виды, населяющие субтропики и степи обоих континентов, длительное время эволюционировали самостоятельно и различаются существенно (на уровне родов и семейств). Обитатели лесов разделились сравнительно недавно, поэтому их эволюционное расхождение не так велико – на уровне близких видов одного рода, подвидов одного вида, даже общих видов. Тундровые обитатели разделились совсем недавно, поэтому здесь обитают в основном те же виды, иногда подвиды одного вида, реже близкие виды одного рода. В то же время длительная изоляция между Северной и Южной Америкой привела к тому, что здесь крайне мало общих видов, в основном за счет миграций уже в Голоцене. Панамский перешеек не очень способствует миграциям, т. к. за ним находится непреодолимая для многих видов полоса пустынь. Больше мигрантов в обе стороны проникали через Карибский мост, существовавший в эпоху наибольшего оледенения и в связи с этим обмеления морей. Поэтому, например, Флорида имеет больше сходства с Южной Америкой, чем Мексика.
Раннее разделение Южной Америки от Африки вызвало несходство их современных биот, несмотря на сходство климатов. В то же время соединение Африки с Южной Азией, более влажный, чем теперь, климат в зоне соединения и отсутствие рифтовых разломов (нынешние Красное море и Персидский залив) способствовали интенсивному обмену флорой и фауной между ними. Возникновение упомянутых рифтов и опустынивание Северной Африки, Аравии и части Ближнего Востока привело к разделу этих биот, но, так как это произошло в сравнительно недавнее время, биоты их еще имеют много общего. Отрезанные от тропиков пустынями Северная Африка и Ближний Восток подверглись сильному влиянию биоты умеренного пояса Евразии, представители которой вытеснили остатки тропических растений и животных. Этому способствовало то, что во время наиболее сильных оледенений существовал континентальный мост от Апеннинского полуострова через Сицилию и Мальту, и отсутствие Гибралтарского пролива.
В настоящее время существует некоторая разница в выделении флористических и фаунистических царств, что затрудняет создание общеупотребительной системы биогеографических царств.
Ботаники выделяют шесть флористических царств, подразделяемых на ряд областей (по A. Л. Тахтаджяну).
1. Голарктическое царство. Занимает большую часть Евразии, кроме самого юга и юго-востока и юго-запада Аравии, почти всю Северную Америку, кроме самого юга Мексики и Флориды. Делится на области: циркумбореальную (зоны тундры, лесотундры, вся лесная и степная зоны Евразии, тундра, лесотундра и тайга Северной Америки), восточноазиатскую (хвойно-широколиственные леса и субтропики Японии, Кореи, Восточного Китая, Российское Приморье и Сахалин), Аппалачско-североамериканскую (от Атлантического океана до Скалистых гор и от Великих Озер до Мексиканского залива), область Скалистых гор, Макаронрезийскую (Канарские, Азорские о-ва и о. Мадейра), Средиземноморскую, Ирано-Туранскую (от Ливанских гор и Закавказья до среднеазиатских пустынно и гор, до Гоби и Тибета включительно), и Мадреанскую (пустыни и субтропики, юго-запад США и Мексики).
2. Палеотропическое царство. Занимает большую часть Африки к югу от Сахары (кроме самого юга), Мадагаскар, южную и юго-восточную Азию, Зондские и Филиппинские острова и большую часть островов Тихого океана, включая Гавайские. Делится на области: Гвинео– Конголезскую (побережье Гвинейского залива и бассейн р. Конго), Судано-Замбезийскую (почти вся Африка, кроме Гвиано-Конголезской области, Сахары и крайнего юга), Карру-Намибийская (крайний юг-запад Африки), островов св. Елены и Вознесения (упомянутые острова в Атлантическом океане), Мадагаскарскую (о. Мадагаскар с прилегающими островами), Индийскую (п-ов Индостан), Индокитайскую (п-ов Индокитай, южный Китай и северная часть Малаккского п-ва), Малезийскую (южная часть Малаккского п-ва, острова Филиппинские и Зондские, включая Новую Гвинею с прилегающими островами), Фиджийскую (о-ва Фиджи и прилегающие), Гавайскую (Гавайские о-ва), Полинезийскую (острова между Фиджи и Гавайскими), Новокаледонскую (о. Новая Каледония и прилегающие).
3. Неотропическое царство. Занимает Флориду, Панамский перешеек, острова Карибского моря и Южную Америку на юг до устья Ла-Платы. Делится на области: Карибскую (острова Карибского моря, Панамский перешеек, Флорида и крайний северо-запад Южной Америки), Амазонскую (большая часть бассейнов Амазонки и Ориноко), Гвианского нагорья (небольшой участок на северо-западе Амазонской области), Бразильскую (к югу от Амазонской до устья Ла-Платы и до побережья Атлантического океана) и Андийскую (горы Анды на юг немного южнее южного тропика).
4. Капское царство. Самый юг Африки, между Драконовыми горами и океаном.
5. Австралийское царство. Континент Австралии и остров Тасмания. Делится на области: Северо-Восточную (северное и восточное побережья Австралии и о. Тасмания), Юго-Западную (крайний юго– запад континента) и Центральную (большая часть Австралии, центр континента).
6. Голантарктическое царство. Делится на области: Островов Хуан-Фернандес (упомянутые острова), Чилийско-Патагонскую (Южная Америка к югу от устья Ла-Платы), Новозеландскую (Новая Зеландия) и субантарктических островов (мелкие острова по периферии Антарктиды).
Зоологи также выделяют шесть зоогеографических царств (по В. Г. Гептнеру).
6.1. Голарктическое царство. Включает всю северную Америку (с Флоридой), всю Евразию кроме крайнего юга и юго-востока и север Африки. Делится на области: Арктическую (тундры и лесотундры обоих континентов и острова Северного Ледовитого океана), Канадскую (большая часть Северной Америки кроме южной части континента), Европейско-Сибирскую (лесная и степная зоны Евразии), Средиземноморскую (побережье Средиземного моря, Сахара, Аравия, Кавказ, Ближний и Средний Восток до гор Средней Азии, на восток до Индии), Центральноазиатскую (пустыни и горы Средней Азии, Гоби и Тибет), Маньчжурско-Китайскую (Япония, Корея, Восточный Китай, российское Приморье, Сахалин), Сонорскую (южная часть Северной Америки).
6.2. Неотропическое царство. Острова Карибского моря, Панамский перешеек и вся Южная Америка. Делится на области: Центральноамериканскую (Панамский перешеек), Антильскую (острова Карибского моря), Бразильскую (северная часть материка, бассейны Амазонки и Ориноко, до Атлантического океана), Чилийскую (Анды и южная часть континента, к югу от устья Ла-Платы).
6.3. Эфиопское царство. Африка к югу от Сахары и Мадагаскар. Делится на области: Восточноафриканскую (побережье Гвинейского залива и бассейн р. Конго), Западноафриканскую (остальная часть Африки кроме юга), Южноафриканскую (юг континента), Мадагаскарскую (Мадагаскар).
6.4. Восточное (Индо-Малайсоке) царство. Индостан и Индокитай, Филиппинские и Зондские острова (до Новой Гвинеи). Делится на области: Индийскую (полуостров Индостан и Цейлон), Индомалайскую (Индокитай, южный Китай, Филиппинские и Зондские острова до Новой Гвинеи).
6.5. Австралазийское царство. Включает Австралию и все острова Тихого океана вплоть до Гавайских. Делится на области: Австралийскую (вся Австралия, кроме самого севера), Папуасскую (север Австралии, Новая Гвинея с прилегающими островами), Новозеландскую (Новая Зеландия), Полинезийскую (острова в Тихом океане, кроме Гавайских), Гавайскую (Гавайские острова).
6.6. Антарктическое царство. Мелкие острова вдоль побережья Антарктиды.
Иногда Голарктическое царство делят на два подцарства: Heap ктику (Северная Америка) и Палеарктику (Евразия). Также иногда выделяют лишь четыре царства: Арктогею (Голарктическое царство), Неогею (Неотропическое царство), Палеогею (Эфиопское и Восточное царства, делят на три области: Эфиопскую, Мадагаскарскую и Восточную). Нотогея (Австралазийское царство). Антарктическая область не входит по этой классификации, ни в одно из царств.
Из изложенного видно, что довольно трудно совместить зоогеографическое и флористическое районирование в единое биогеографическое. По А. Г. Воронову, Н. Н. Дроздову и В. В. Второву, выделяется восемь биогеографических царств: Голарктическое, Неотропическое, Эфиопское, Восточное, Австралийское, Антарктическое, Мадагаскарское и Капское. В основном принято зоогеографическое деление, только в последнем случае приоритет отдан флористическому.
Биогеографическое разделение Мирового Океана не столь четко. Флористы выделяют здесь три царства: Бореальное (к северу от северного тропика), Тропическое (от северного тропика до южного) и Южное (к югу от южного тропика). Зоогеографы выделяют пять царств: Арктическое, Атлантическое, Тихоокеанское, Тропическое (Индийский океан и тропические зоны Тихого и Атлантического океанов) и Антарктическое (южные части всех трех океанов).
Часть 2
Экологическая характеристика основных природных зон земного шара
Глава 11
Экосистемы тундры и лесотундры
Ландшафтно-географическая зона тундры характеризуется наиболее простым строением основных экосистем, бедным видовым составом живых организмов, простотой и неустойчивостью биоценотических связей, малой биомассой и низкой продуктивностью.
Тундра занимает материковый север Евразии и Северной Америки, а также прилегающие к материкам острова. Далеко отстоящие острова заняты арктическими пустынями. На южной границе тундры располагается переходный биом – лесотундра. По горам тундровые экосистемы проникают далеко на юг вглубь Полярного Урала, гор Восточной Сибири, Аляски, Скандинавии, смыкаясь с высокогорной тундрой, которая, собственно, относится уже к другому типу экосистем – к горным.
Рельеф тундровой зоны однообразен. Здесь нет сколько‑нибудь высоких гор, но снеговая линия лежит очень низко. Так, горная страна Бырранга на Таймыре имеет наибольшую высоту 1 146 м, но уже на высоте 800 м начинаются вечные снега, имеются постоянные ледники. Равнинные тундровые экосистемы здесь непосредственно переходят в горнотундровые. Речные долины в тундрах – это в основном низовья очень крупных рек, текущих с юга (Печора, Обь, Енисей, Лена, Макензи, Коппермайн и др.), имеются поймы очень большой ширины, много островов, т. к. в низовьях преобладают аккумулятивные процессы – нанос обломочного материала, формирующего острова и мели (дельта Лены). Если пойма рек прорезает более возвышенный берег, образуются сравнительно узкие и длинные заливы, глубоко врезающиеся в материк – губы (Обско-Тазовская губа), а небольшие реки в гористых берегах образуют очень глубокие и сравнительно узкие заливы – фьорды (в Скандинавии).
Между долин крупных рек располагаются волнистые равнины с многочисленными озерами. Крупные озера – в основном реликты отступающих ледников, котловины, заполненные талыми водами последних (оз. Таймыр), мелкие озера – в основном термокарстовые воронки, заполненные снеговой водой. Вблизи крупных рек располагаются соры – озера, образовавшиеся путем подпруживания вод притоков водами главной реки.
Климат суровый. Зима продолжительная, морозная, средняя температура января -20 °C —30 °C, длится 7–9 месяцев, абсолютные минимумы температуры ниже -50 °C. Короткое лето с температурой самого теплого месяца – июля – не более +10 °C, причем даже в июле возможны заморозки и снегопады. Сильные ветра увеличивают суровость климата. Вечная мерзлота препятствует произрастанию древесной растительности. Хотя осадков в тундре выпадает очень мало – от 100 до 300 мм в год, причем до 90 % осадков выпадет летом – низкая температура делает испарение минимальным. А так как мерзлота препятствует просачиванию воды вглубь, а равнинный рельеф приводит к тому, что поверхностный сток невелик, большая часть поверхности тундры заболочена. Причем болота находятся не только в низинах, но и на склонах и даже на возвышенных местах рельефа. Мерзлота же обусловливает сильное развитие таких форм рельефа, как бугры вспучивания и морозобойные трещины.
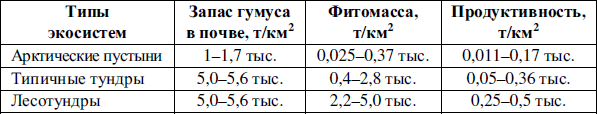
В холодном климате тундр микроорганизмы малоактивны и микробное разложение отмершей органики замедлено. Растительный покров подстилается толстой губчатой подушкой из неразложившейся мертвой растительности, сильно пропитанной водой. На возвышенных местах может скапливаться войлокообразная масса из неперегнившей высохшей растительности. Все это приводит к бедности тундровых почв гумусом, а также к низкой продуктивности тундровых сообществ и их сравнительно небольшой биомассе (таблица 4), причем более половины этой биомассы – 55 % – приходится на корневую систему.
Таблица 4
Запасы гумуса в почве, фптомасса и продуктивность различных тундровых экосистем
Растительность тундр своеобразна. На островах Северного Ледовитого океана (кроме приближенных к материкам) развиты арктические пустыни. Большая часть таких островов покрыта льдом, а свободные от ледников участки почти полностью покрыты щебенкой, и лишь около 30 % их поверхности заняты растительностью. Это небольшие «садики» на южных, лучше прогреваемых склонах, где‑нибудь в расщелинах между камней, где есть укрытие от ветров. Здесь растут мхи, лишайники. Некоторые цветковые – полярный мак (Papaver radicatum), полемониум (Polemonium humile) и нек. др. На ледниках встречаются одноклеточные сине-зеленые водоросли (Ancilonema nordenscheldii), окрашивающие значительные площади ледовых полей в красно-бурый цвет.
Материковые тундры имеют молодые растительные сообщества, образовавшиеся в конце Плейстоцена – начале Голоцена. Для них характерно господство хамефитов и гемикриптофитов. Очень большой процент тундровых растений вечнозеленые, имеющие весной уже готовый аппарат для фотосинтеза. Многие растения низкорослы – сверху рост ограничивается сильными ветрами, которые иссушают и обламывают все, что поднимается зимой выше уровня снежного покрова, снизу – вечной мерзлотой (глубина ее обычно 15–20 см, часто меньше). По этой же причине в тундре нет древесной растительности. Растения прижимаются к почве, температура которой летом часто бывает выше температуры воздуха, а зимой скрываются под снегом. Снеговой покров глубиной до 0,5 м при наружной температуре -55 °C сохраняет на уровне поверхности почвы всего лишь -7 °C.
Флористически тундры бедны. На северных островах Земли Франца-Иосифа отмечено 38 видов растений, в наиболее богатых видами материковых евразийских и канадских тундрах – до 450 видов.
Наиболее многочисленны мхи и лишайники, образующие два основных ландшафтных типа тундр. Наиболее распространены моховые тундры. Они часто заболочены, состоят из сильно переплетенных моховых дернин из нескольких видов зеленых мхов (Aulacomnium sp., Camtothecium sp., Dicranum sp. и др.), над которыми располагается сильно разреженный травяной ярус из мятлика (Роа arctica), осок (Carex sp.), горца (Polygonum viviparum) и кустарничковый из дриады (Dryas punctata), голубики (Vaccinium uliginosum), брусники (V. vitis idaea), карликовой березки (Betula nana), нескольких видов карликовых ив (Salix sp.). В южной части тундры кустарничковый ярус становится сплошным, преобладает карликовая березка. В таких местах бывает много грибов (подберезовики, Leccinum scabram), шляпки которых возвышаются над стелющимися во мху стволиками карликовых березок и ив.
Тундра занимает материковый север Евразии и Северной Америки, а также прилегающие к материкам острова. Далеко отстоящие острова заняты арктическими пустынями. На южной границе тундры располагается переходный биом – лесотундра. По горам тундровые экосистемы проникают далеко на юг вглубь Полярного Урала, гор Восточной Сибири, Аляски, Скандинавии, смыкаясь с высокогорной тундрой, которая, собственно, относится уже к другому типу экосистем – к горным.
Рельеф тундровой зоны однообразен. Здесь нет сколько‑нибудь высоких гор, но снеговая линия лежит очень низко. Так, горная страна Бырранга на Таймыре имеет наибольшую высоту 1 146 м, но уже на высоте 800 м начинаются вечные снега, имеются постоянные ледники. Равнинные тундровые экосистемы здесь непосредственно переходят в горнотундровые. Речные долины в тундрах – это в основном низовья очень крупных рек, текущих с юга (Печора, Обь, Енисей, Лена, Макензи, Коппермайн и др.), имеются поймы очень большой ширины, много островов, т. к. в низовьях преобладают аккумулятивные процессы – нанос обломочного материала, формирующего острова и мели (дельта Лены). Если пойма рек прорезает более возвышенный берег, образуются сравнительно узкие и длинные заливы, глубоко врезающиеся в материк – губы (Обско-Тазовская губа), а небольшие реки в гористых берегах образуют очень глубокие и сравнительно узкие заливы – фьорды (в Скандинавии).
Между долин крупных рек располагаются волнистые равнины с многочисленными озерами. Крупные озера – в основном реликты отступающих ледников, котловины, заполненные талыми водами последних (оз. Таймыр), мелкие озера – в основном термокарстовые воронки, заполненные снеговой водой. Вблизи крупных рек располагаются соры – озера, образовавшиеся путем подпруживания вод притоков водами главной реки.
Климат суровый. Зима продолжительная, морозная, средняя температура января -20 °C —30 °C, длится 7–9 месяцев, абсолютные минимумы температуры ниже -50 °C. Короткое лето с температурой самого теплого месяца – июля – не более +10 °C, причем даже в июле возможны заморозки и снегопады. Сильные ветра увеличивают суровость климата. Вечная мерзлота препятствует произрастанию древесной растительности. Хотя осадков в тундре выпадает очень мало – от 100 до 300 мм в год, причем до 90 % осадков выпадет летом – низкая температура делает испарение минимальным. А так как мерзлота препятствует просачиванию воды вглубь, а равнинный рельеф приводит к тому, что поверхностный сток невелик, большая часть поверхности тундры заболочена. Причем болота находятся не только в низинах, но и на склонах и даже на возвышенных местах рельефа. Мерзлота же обусловливает сильное развитие таких форм рельефа, как бугры вспучивания и морозобойные трещины.
В холодном климате тундр микроорганизмы малоактивны и микробное разложение отмершей органики замедлено. Растительный покров подстилается толстой губчатой подушкой из неразложившейся мертвой растительности, сильно пропитанной водой. На возвышенных местах может скапливаться войлокообразная масса из неперегнившей высохшей растительности. Все это приводит к бедности тундровых почв гумусом, а также к низкой продуктивности тундровых сообществ и их сравнительно небольшой биомассе (таблица 4), причем более половины этой биомассы – 55 % – приходится на корневую систему.
Таблица 4
Запасы гумуса в почве, фптомасса и продуктивность различных тундровых экосистем
Растительность тундр своеобразна. На островах Северного Ледовитого океана (кроме приближенных к материкам) развиты арктические пустыни. Большая часть таких островов покрыта льдом, а свободные от ледников участки почти полностью покрыты щебенкой, и лишь около 30 % их поверхности заняты растительностью. Это небольшие «садики» на южных, лучше прогреваемых склонах, где‑нибудь в расщелинах между камней, где есть укрытие от ветров. Здесь растут мхи, лишайники. Некоторые цветковые – полярный мак (Papaver radicatum), полемониум (Polemonium humile) и нек. др. На ледниках встречаются одноклеточные сине-зеленые водоросли (Ancilonema nordenscheldii), окрашивающие значительные площади ледовых полей в красно-бурый цвет.
Материковые тундры имеют молодые растительные сообщества, образовавшиеся в конце Плейстоцена – начале Голоцена. Для них характерно господство хамефитов и гемикриптофитов. Очень большой процент тундровых растений вечнозеленые, имеющие весной уже готовый аппарат для фотосинтеза. Многие растения низкорослы – сверху рост ограничивается сильными ветрами, которые иссушают и обламывают все, что поднимается зимой выше уровня снежного покрова, снизу – вечной мерзлотой (глубина ее обычно 15–20 см, часто меньше). По этой же причине в тундре нет древесной растительности. Растения прижимаются к почве, температура которой летом часто бывает выше температуры воздуха, а зимой скрываются под снегом. Снеговой покров глубиной до 0,5 м при наружной температуре -55 °C сохраняет на уровне поверхности почвы всего лишь -7 °C.
Флористически тундры бедны. На северных островах Земли Франца-Иосифа отмечено 38 видов растений, в наиболее богатых видами материковых евразийских и канадских тундрах – до 450 видов.
Наиболее многочисленны мхи и лишайники, образующие два основных ландшафтных типа тундр. Наиболее распространены моховые тундры. Они часто заболочены, состоят из сильно переплетенных моховых дернин из нескольких видов зеленых мхов (Aulacomnium sp., Camtothecium sp., Dicranum sp. и др.), над которыми располагается сильно разреженный травяной ярус из мятлика (Роа arctica), осок (Carex sp.), горца (Polygonum viviparum) и кустарничковый из дриады (Dryas punctata), голубики (Vaccinium uliginosum), брусники (V. vitis idaea), карликовой березки (Betula nana), нескольких видов карликовых ив (Salix sp.). В южной части тундры кустарничковый ярус становится сплошным, преобладает карликовая березка. В таких местах бывает много грибов (подберезовики, Leccinum scabram), шляпки которых возвышаются над стелющимися во мху стволиками карликовых березок и ив.