Страница:
Регуляция силы и скорости мышечного сокращения.
Как вы знаете на собственном опыте, человек обладает возможностью произвольно регулировать силу и скорость мышечного сокращения. Реализуется эта возможность несколькими способами. С одним из них вы уже знакомы – это управление частотой нервной импульсации. Подавая волокну одиночные команды на сокращение, можно добиться в нем легкого напряжения. Так, например, поддерживающие позу мышцы слегка напряжены, даже когда человек отдыхает. Повышая частоту импульсов можно увеличивать силу сокращения до максимально возможной для данного волокна в данных условиях работы, которая достигается при слиянии отдельных импульсов в тетанус.Сила, развиваемая волокном в состоянии тетануса, не всегда одинакова и зависит от характера и скорости движения. При статическом напряжении (когда длина волокна остается постоянной) сила, развиваемая волокном, больше чем при сокращении волокна, и чем быстрее сокращается волокно, тем меньшую силу оно способно развить. Максимальную же силу волокно развивает в условиях негативного движения, то есть при удлинении волокна.
При отсутствии внешней нагрузки волокно сокращается с максимальной скоростью. При увеличении нагрузки скорость сокращения волокна уменьшается и по достижении определенного уровня нагрузки падает до нуля, при дальнейшем увеличении нагрузки волокно удлиняется.
Причину в различии силы волокна при различных направлениях движения легко понять, рассмотрев уже приведенный ранее пример с гребцами и веслами. Дело в том, что после завершения «гребка» миозиновый мостик некоторое время находится в состоянии сцепления с нитью актина, представьте, что весло после гребка так же не сразу извлекается из воды, а находится погруженным еще некоторое время. В случае, когда гребцы плывут вперед (позитивное движение), весла, остающиеся погруженными в воду после завершения гребка, тормозят движение и мешают плыть, в то же время, если лодка буксируется назад, а гребцы сопротивляются этому движению, то погруженные весла так же мешают движению, и буксиру приходится прилагать большие усилия. То есть, при сокращении волокна, сцепленные мостики мешают движению и ослабляют силу волокна, при негативном движении – удлинении мышцы – не отцепленные мостики так же мешают движению, но в этом случае они как бы поддерживают опускающийся вес, что и позволяет волокну развивать большую силу. Легче всего понять различия между статическим напряжением, позитивным и негативным движением взглянув на Рис.5.
Итак, мы рассмотрели основные факторы, влияющие на силу и скорость сокращения отдельного волокна. Сила же сокращения целой мышцы зависит от количества волокон в данный момент времени вовлеченных в работу.
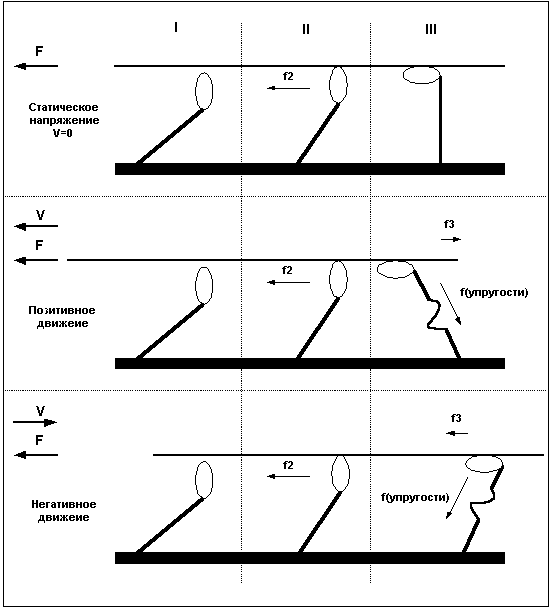
Рис.5
Вовлечение волокон в работу.
При поступлении от ЦНС (центральной нервной системы) к мотонейронам (расположенным в спинном мозге), возбуждающего сигнала (запускающий импульс), мембрана мотонейрона поляризуется, и он генерирует серию импульсов, направляемых по аксону к волокнам. Чем сильнее воздействие на мотонейрон (поляризация мембраны), тем выше частота генерируемого им импульса – от небольшой стартовой частоты (4-5 Гц), до максимально возможной, для данного мотонейрона, частоты (50 Гц и более). Быстрые мотонейроны способны генерировать гораздо более высокочастотный импульс, чем медленные, поэтому сила сокращения быстрых волокон гораздо больше подвержена частотной регуляции, чем сила медленных.В то же время имеется и обратная связь с мышцей, от которой поступают тормозящие сигналы, уменьшающие поляризацию мембраны мотонейрона и снижающие его ответ.
Каждый мотонейрон имеет свой порог возбудимости. Если сумма возбуждающих и тормозящих сигналов, превышает этот порог и на мембране достигается необходимый уровень поляризации, то мотонейрон вовлекается в работу. Медленные мотонейроны имеют, как правило, низкий порог возбудимости, а быстрые – высокий. Мотонейроны же целой мышцы имеют широкий спектр значений этого параметра. Таким образом, при повышении силы сигнала ЦНС, активируется все большее число мотонейронов, а мотонейроны с низким порогом возбудимости увеличивают частоту генерируемого импульса.
Когда требуется легкое усилие, например, при ходьбе или беге трусцой, активируется небольшое число медленных мотонейронов и соответствующее число медленных волокон, ввиду высокой выносливости этих волокон такая работа может поддерживаться очень долго. По мере увеличения нагрузки ЦНС приходится посылать все более сильный сигнал, и большее число мотонейронов (а, следовательно, волокон), вовлекается в работу, а те, что уже работали, увеличивают силу сокращения, по причине увеличения частоты импульсации, поступающей от мотонейронов. По мере увеличения нагрузки в работу включаются быстрые окислительные волокна, а по достижения определенного уровня нагрузки (20%-25% от максимума), например, во время подъема в гору или финального спурта, силы окислительных волокон становится недостаточно, и посылаемый ЦНС сигнал включает в работу быстрые – гликолитические волокна. Быстрые волокна значительно повышают силу сокращения мышцы, но, в свою очередь, быстро утомляются, и в работу вовлекается все большее их количество. Если уровень внешней нагрузки не уменьшается, работу в скором времени приходится останавливать из-за усталости, в результате накопления молочной кислоты.
Скорость движения мышцы зависит от соотношения внешней нагрузки и количества вовлеченных в движение волокон. Мобилизация относительно небольшого количества волокон, приведет к медленному движению, как только совместная сила сокращающихся волокон превысит уровень внешней силы. Мобилизация все большего количества волокон при той же внешней нагрузке приведет к увеличению скорости сокращения, а увеличение внешней нагрузки, или падение силы волокон в результате усталости, при неизменном количестве мобилизованных волокон, приведет к падению скорости сокращения.
При предельных нагрузках, например, при подъеме максимального веса или подъеме относительно небольшого веса, но с максимальной скоростью, сокращается сразу максимально возможное для данного индивида число волокон.
По вопросу вовлечения волокон при предельных нагрузках в литературе приводятся противоречивые сведения. Так ряд авторов утверждает, что даже у тренированных спортсменов при предельных нагрузках активно не более 85% от общего количества двигательных единиц в мышце. С другой стороны, Гурфинкель и Левин приводят данные, что большая часть двигательных единиц рекрутируется уже при нагрузках до 50% от максимума, и в дальнейшем, при повышении силы, подключается лишь небольшая часть (около 10%) самых крупных двигательных единиц, а рост силы от 75% до 100% происходит уже не за счет вовлечения новых двигательных единиц, а за счет повышения частоты импульса, генерируемого мотонейронами. Независимо от того, чья точка зрения является правильной, важно одно – сила мышцы зависит от силы импульса ЦНС, а чем обусловлен рост силы при нагрузках свыше 75% от максимальной только ли ростом частоты, генерируемой мотонейронами, или еще и вовлечением в работу новых мотонейронов не принципиально. Важно так же и то, что в мышцу встроен ограничительный механизм контроля над развиваемым напряжением, осуществляемый через сухожильные органы Гольджи. Сухожильные рецепторы регистрируют напряжение и при превышении критического значения оказывают тормозящее воздействие на мотонейроны. Считается, что при отключении контроля над напряжением проявляется «сверхсила» человека, регистрируемая в экстремальных ситуациях.
Часть 2: Теория тренировки.
Первую часть статьи я посвятил краткому изложению основ физиологии мышечной деятельности и, пожалуй, пора вознаградить терпение читателей и приступить к рассмотрению самого главного вопроса, интересующего всех приверженцев железного спорта, – что заставляет наши мышцы увеличивать объем и силу? Но, прежде чем изложить собственные взгляды по этому предмету, я хотел бы ознакомить читателя с основами классической теорией тренировки.
Организм – саморегулируемая система, стремящаяся к поддержанию постоянства внутренней среды. Физическая нагрузка оказывает выраженное воздействие на внутреннюю среду мышц и организма в целом, смещая многие биохимические константы от уровня, характерного для гомеостаза покоя, к состоянию, соответствующему гомеостазу деятельности. Степень этих изменений зависит от характера и интенсивности физической нагрузки и индивидуальной реакции на нее организма, отражающей уровень тренированности. Сразу после прекращения нагрузки, в организме начинаются процессы, стремящиеся восстановить исходное состояние, соответствующее гомеостазу покоя, в ходе этих процессов закрепляются изменения, позволяющие в дальнейшем минимизировать возмущение внутренней среды при аналогичных нагрузках.
Спортивную тренировку можно рассматривать как процесс направленного приспособления организма (адаптации) к воздействию тренировочных нагрузок.
Различают срочную и долговременную адаптацию.
Срочная адаптация – это ответ организма на однократное воздействие тренировочной нагрузки, выраженный в «аварийном» приспособлении к изменившемуся состоянию внутренней среды и сводящийся, преимущественно, к изменениям в энергетическом обмене и к активации высших нервных центров, ответственных за регуляцию энергетического обмена.
Долговременная адаптация развивается постепенно на основе многократной реализации срочной адаптации путем суммирования следов повторяющихся нагрузок.
В развитии процессов адаптации различают специфическую компоненту и общую адаптационную реакцию.
Процессы специфической адаптация затрагивают внутриклеточный энергетический и пластический обмен и связанные с ним функции вегетативного обслуживания, которые специфически реагируют на данный вид воздействия пропорционально его силе.
Общая адаптационная реакция развивается в ответ на самые различные раздражители не зависимо от их природы, в случае, если сила этих раздражителей превышает определенный пороговый уровень. Реализуется общая адаптационная реакция благодаря возбуждению симпато-адреалиновой и гипофизарно-адренокортикальной систем, в результате активации которых в крови и тканях повышается содержание катехоламинов и глюкокортикоидов, что способствует мобилизации энергетических и пластических резервов организма. Такая неспецифическая реакция на раздражение получила название «синдром стресса», а раздражители, вызывающие эту реакцию, обозначают как стресс-факторы. Общий адаптационный синдром сам по себе не является основой адаптации к тренировочным нагрузкам, а лишь призван на системном уровне обеспечивать протекание специфических адаптационных реакций, которые и реализуют приспособление организма к конкретным видам нагрузки.
Не смотря на различную природу процессов специфической адаптации можно выделить общие закономерности их протекания. Основу специфической адаптации составляют процессы восстановления сниженных во время мышечной работы энергетических ресурсов, разрушенных структур клеток, смещенного водно-электролитического баланса и др. Наглядно проследить закономерности протекания восстановительных процессов можно на примере восстановления энергетических ресурсов организма, так как, при физических нагрузках наиболее выраженные изменения обнаруживаются именно в сфере энергетического обмена.
Мышечная работа, в зависимости от интенсивности и длительности, приводит к снижению уровня креатинфосфата в мышцах, истощению запасов внутримышечного гликогена и гликогена печени, резервов жиров. Интенсивно протекающие после прекращения нагрузки процессы восстановления приводят к тому, что в определенный момент отдыха после работы уровень энергетических веществ превышает исходный дорабочий уровень. Это явление получило название суперкомпенсация, или сверхвосстановление (рис. 1).
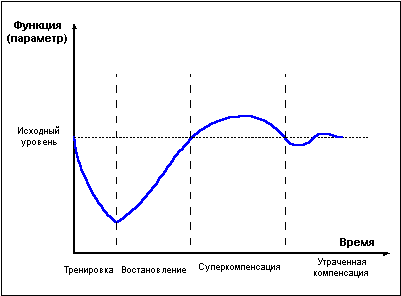
Похожим образом развивается восстановление не только энергетических, но и пластических ресурсов организма, и даже целых тренируемых функций. Напряжение в ходе физической нагрузки систем, ответственных за реализацию той или иной функции, приводит к снижению функциональных возможностей организма, затем во время отдыха достигается состояние суперкомпенсации тренируемой функции, длящееся определенное ограниченное время, далее, при отсутствии повторных нагрузок, уровень тренируемой функции вновь снижается, – наступает фаза утраченной суперкомпенсации (рис 1).
Развитие долговременной адаптации становится возможным только в случае, если достигаемый срочный и отставленный тренировочный эффект от каждой тренировки будет суммироваться по определенным правилам.
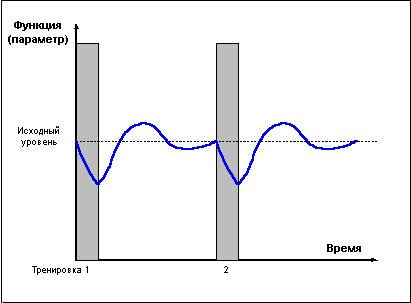
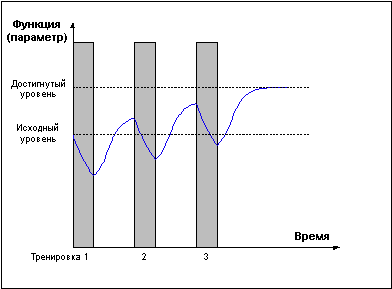
Проведение повторных тренировок в фазе утраченной суперкомпенсации (слишком редкие тренировки) (Рис. 2) не сможет привести к закреплению тренировочного эффекта, так как каждая последующая тренировка проводится после возвращения функциональных возможностей организма к исходному уровню.
Поэтому в классической спортивной школе годичный (и даже многолетний) период тренировок разбивают на микро и макроциклы в ходе которых ставятся задачи по развитию определенных тренируемых качеств, чередование тренировочных занятий в ходе микроциклов осуществляется таким образом, что бы физические нагрузки, направленные на развитие определенного двигательного качества задавались через промежутки времени, обеспечивающие суперкомпенсацию ведущей функции, а нагрузки иной направленности, применяемые в этот период, не оказывали отрицательного влияния на восстановление основной функции. Однако такой метод срабатывает только при развитии взаимонезависимых функций или параметров. В случае если определенное двигательное качество в равной мере зависит от развития нескольких функций или параметров, испытывающих одновременное напряжение в ходе одного тренировочного занятия и имеющих разное время восстановления, в течение микроцикла приходиться варьировать интенсивностью и объемом тренировок, накладывая волны восстановления различных параметров друг на друга таким образом, что бы получить суперкомпенсацию основных тренируемых функций к моменту завершения микроцикла.
Как вы видите, классическая теория тренировки оперирует такими понятиями как двигательные качества, функциональные возможности, и основывается на изучение процессов, приводящих к росту работоспособности мышц и организма в целом в различных режимах работы. Основой практически всех видов спорта является именно работоспособность, и цель планируемых адаптационных изменений в организме спортсмена, как правило, – выход на новый уровень работоспособности. Гипертрофия мышц вовсе не является целью тренинга в классическом спорте, и воспринимается лишь как побочный продукт развития основных двигательных качеств, более того, в некоторых случаях, гипертрофия мышц может оказывать даже отрицательное влияние на достижение стоящих перед спортсменом целей. Посетителей же тренажерных залов, за редким исключением, в большей степени интересует именно достижение гипертрофии мышц, нежели развитие двигательных качеств и повышения работоспособности. Безусловно, развитие работоспособности мышц способствует гипертрофии мышечной ткани.
Процессы, направленные на улучшение доставки кислорода к мышцам существенно развивают капиллярную сеть, что способствует общей гипертрофии мышц.
Тренировка окислительной активности мышц приводит к значительному росту в саркоплазме мышечных волокон количества и объема митохондрий – энергетических станций клетки.
Последовательные процессы суперкомпенсации внутримышечных запасов гликогена приводят к значительному его накоплению, что, в свою очередь, увеличивает объем саркоплазмы мышечного волокна.
Накопление иных веществ, ответственных за энергообеспечение, мышечной деятельности, таких, например, как креатинфосфат, так же увеличивает объем саркоплазмы, и даже не столько за счет объемов самих этих веществ, сколько за счет сопутствующего увеличения объема внутриклеточной жидкости.
То есть, тренировка работоспособности мышц приводит к гипертрофии мышечных волокон в основном за счет увеличения объема саркоплазмы.
Но самый существенный вклад в рост объемов и силы сокращения волокна вносит гипертрофия миофибрилл, все остальные компоненты клетки призваны лишь обеспечивать их сократительную активность. Как размер топливных баков самолета зависит от мощности его турбин, так и объем саркоплазмы мышечного волокна зависит от развития сократительного аппарата клетки (ну и конечно от объема работы, регулярно выполняемой мышцами).
Как я упоминал в первой части, миофибриллы представляют из себя белковые нити, поэтому увеличение количества и поперечного сечения миофибрилл в волокне напрямую связано с интенсивностью синтеза белка клеткой. Сейчас является неопровержимым фактом то, что тренировка интенсифицирует синтез белка, но вот вопрос, – как и почему это происходит?
Синтез белка очень сложный процесс и его интенсивность зависит от огромного количества факторов.
Прежде всего, считывание мРНК в ядре начинается под воздействием стероидных гормонов, вырабатываемых железами внутренней секреции и поступающими в кровь. Молекула гормона из крови проникает в клетку, где, с помощью белка рецептора, доставляется в ядро и разблокирует участок цепочки ДНК, ответственный за тот или иной белок, после чего становится возможной транскрипция мРНК.
Для запуска транскрипции РНК, необходимо так же развернуть спираль ДНК, для чего используется фермент РНК-полимераза.
На синтез белка сильнейшее влияние оказывает гормон роста (СТГ или соматотропин). По химическому составу соматотропин это белок, поэтому он не может свободно проникать в клетку, в отличие от стероидных гормонов, а воздействует на рецепторы, расположенные на поверхности клетки. Механизм действия гормона роста до конца не изучен, но точно известно, что он стимулирует деятельность РНК-полимераз и рибосомного аппарата клетки.
Ну и конечно для сборки белка требуется наличие в клетке достаточного количества аминокислот и запасов энергии. Без аминокислот не из чего будет строить белок, а энергия нужна для сборки молекулы.
И так, для успешного синтеза белка требуются, как минимум, следующие условия:
– высокий уровень анаболических гормонов в крови (тестостерона и соматотропина)
– наличие в клетке белков-рецепторов тестостерона
– активность ферментов и факторов транскрипции РНК (РНК-полимераз и др.)
– достаточное количество аминокислот в клетке
– запас энергии в клетке
Теперь осталось только ответить на вопрос – как именно тренировка влияет на синтез белка?
Должен разочаровать читателя детально объяснить механизм этого влияния на сегодняшнем уровне развития науки невозможно. Если о том, как происходит регуляция синтеза белка в простейших одноклеточных организмах, когда в клетке может идти строительство всех белков, закодированных в ДНК, ученые имеют определенное представление, то, как осуществляется регуляция синтеза белка в многоклеточных организмах, когда, теоретически, каждая клетка может синтезировать все возможные белки, закодированные в ДНК, но синтезирует лишь набор белков, присущий данному типу клеток, остается пока не известным. Да, гормоно-рецепторный комплекс разблокирует участок ДНК, в котором закодирован определенный белок, но как гормон узнает, какой именно ген в данный момент необходим клетке – миозин быстрого волокна или миозин медленного волокна, а может быть миоглобин? Генетикам предстоит еще долгий путь, прежде чем раскроются все тайны синтеза белка. А как же быть до тех пор?
Организм – саморегулируемая система, стремящаяся к поддержанию постоянства внутренней среды. Физическая нагрузка оказывает выраженное воздействие на внутреннюю среду мышц и организма в целом, смещая многие биохимические константы от уровня, характерного для гомеостаза покоя, к состоянию, соответствующему гомеостазу деятельности. Степень этих изменений зависит от характера и интенсивности физической нагрузки и индивидуальной реакции на нее организма, отражающей уровень тренированности. Сразу после прекращения нагрузки, в организме начинаются процессы, стремящиеся восстановить исходное состояние, соответствующее гомеостазу покоя, в ходе этих процессов закрепляются изменения, позволяющие в дальнейшем минимизировать возмущение внутренней среды при аналогичных нагрузках.
Спортивную тренировку можно рассматривать как процесс направленного приспособления организма (адаптации) к воздействию тренировочных нагрузок.
Различают срочную и долговременную адаптацию.
Срочная адаптация – это ответ организма на однократное воздействие тренировочной нагрузки, выраженный в «аварийном» приспособлении к изменившемуся состоянию внутренней среды и сводящийся, преимущественно, к изменениям в энергетическом обмене и к активации высших нервных центров, ответственных за регуляцию энергетического обмена.
Долговременная адаптация развивается постепенно на основе многократной реализации срочной адаптации путем суммирования следов повторяющихся нагрузок.
В развитии процессов адаптации различают специфическую компоненту и общую адаптационную реакцию.
Процессы специфической адаптация затрагивают внутриклеточный энергетический и пластический обмен и связанные с ним функции вегетативного обслуживания, которые специфически реагируют на данный вид воздействия пропорционально его силе.
Общая адаптационная реакция развивается в ответ на самые различные раздражители не зависимо от их природы, в случае, если сила этих раздражителей превышает определенный пороговый уровень. Реализуется общая адаптационная реакция благодаря возбуждению симпато-адреалиновой и гипофизарно-адренокортикальной систем, в результате активации которых в крови и тканях повышается содержание катехоламинов и глюкокортикоидов, что способствует мобилизации энергетических и пластических резервов организма. Такая неспецифическая реакция на раздражение получила название «синдром стресса», а раздражители, вызывающие эту реакцию, обозначают как стресс-факторы. Общий адаптационный синдром сам по себе не является основой адаптации к тренировочным нагрузкам, а лишь призван на системном уровне обеспечивать протекание специфических адаптационных реакций, которые и реализуют приспособление организма к конкретным видам нагрузки.
Не смотря на различную природу процессов специфической адаптации можно выделить общие закономерности их протекания. Основу специфической адаптации составляют процессы восстановления сниженных во время мышечной работы энергетических ресурсов, разрушенных структур клеток, смещенного водно-электролитического баланса и др. Наглядно проследить закономерности протекания восстановительных процессов можно на примере восстановления энергетических ресурсов организма, так как, при физических нагрузках наиболее выраженные изменения обнаруживаются именно в сфере энергетического обмена.
Мышечная работа, в зависимости от интенсивности и длительности, приводит к снижению уровня креатинфосфата в мышцах, истощению запасов внутримышечного гликогена и гликогена печени, резервов жиров. Интенсивно протекающие после прекращения нагрузки процессы восстановления приводят к тому, что в определенный момент отдыха после работы уровень энергетических веществ превышает исходный дорабочий уровень. Это явление получило название суперкомпенсация, или сверхвосстановление (рис. 1).
Рис.1
Фаза суперкомпенсации длится не вечно, постепенно уровень энергетических веществ возвращается к норме, испытывая некоторые колебания возле состояния равновесия. Чем больше расход энергии при работе, тем интенсивнее идет восстановление и тем значительнее превышение исходного уровня в фазе суперкомпенсации. Однако это правило применимо лишь в ограниченных пределах. При истощающих нагрузках, приводящих к большому накоплению продуктов распада, скорость восстановительных процессов замедляется, фаза суперкомпенсации откладывается во времени и оказывается выраженной в меньшей степени.Похожим образом развивается восстановление не только энергетических, но и пластических ресурсов организма, и даже целых тренируемых функций. Напряжение в ходе физической нагрузки систем, ответственных за реализацию той или иной функции, приводит к снижению функциональных возможностей организма, затем во время отдыха достигается состояние суперкомпенсации тренируемой функции, длящееся определенное ограниченное время, далее, при отсутствии повторных нагрузок, уровень тренируемой функции вновь снижается, – наступает фаза утраченной суперкомпенсации (рис 1).
Развитие долговременной адаптации становится возможным только в случае, если достигаемый срочный и отставленный тренировочный эффект от каждой тренировки будет суммироваться по определенным правилам.
Проведение повторных тренировок в фазе утраченной суперкомпенсации (слишком редкие тренировки) (Рис. 2) не сможет привести к закреплению тренировочного эффекта, так как каждая последующая тренировка проводится после возвращения функциональных возможностей организма к исходному уровню.
Рис. 2.
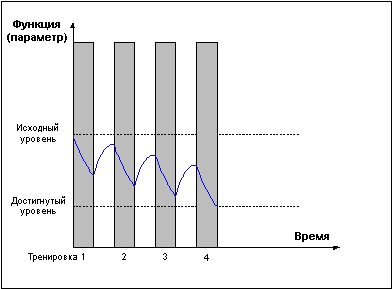
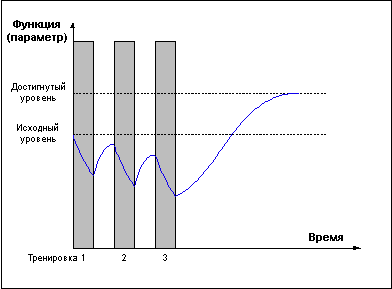
Слишком частые тренировки, прерывающие стадию восстановления до достижения эффекта суперкомпенсации (Рис. 3) приводят к отрицательному взаимодействию тренировочных эффектов и снижению функциональных возможностей организма.Рис. 3.
И только проведение повторных тренировок в фазе суперкомпенсации (Рис. 4) приводит к положительному взаимодействию тренировочных эффектов, закреплению следов срочной адаптации, росту тренируемой функции и формированию долговременной адаптации.Рис. 4.
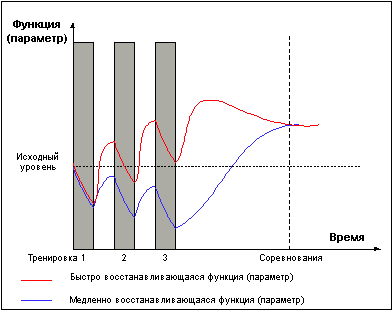
Однако не следует воспринимать приведенные выше правила слишком конкретно. Требование задавать нагрузку только в стадии суперкомпенсации справедливо лишь в долгосрочной перспективе. В рамках одного тренировочного микроцикла возможны серии тренировок в стадии недовосстановления (Рис. 5), приводящие к более глубокому истощению тренируемой функции, что может быть использовано для получения более мощного роста функциональных возможностей в стадии суперкомпенсации, либо для вызванного тактической необходимостью переноса во времени эффекта суперкомпенсации.Рис. 5.
На первый взгляд может показаться, что составление эффективных тренировочных программ является делом не сложным. Достаточно определить уровень нагрузки, необходимый для достижения максимальной суперкомпенсации той или иной функции, и время наступления фазы суперкомпенсации, а далее задавать повторные нагрузки с необходимой частотой, постоянно получая положительное суммирование тренировочных эффектов. На самом деле построить тренировку по такому принципу невозможно. Дело в том, что различные параметры и функции, вносящие свой вклад в общую тренированность, необходимую для того или иного вида спорта, имеют разное время восстановления и достижения суперкомпенсации и разную длительность фазы суперкомпенсации. Так, фаза суперкомпенсации креатинфосфата достигается через несколько минут отдыха после нагрузки, приводящей к существенному снижению его уровня. Для достижения выраженной суперкомпенсации содержания гликогена в мышцах требуется не менее 2-3 суток, к этому моменту уровень креатинфосфата уже вступит в фазу утраченной суперкомпенсации. А вот для восстановления структур клеток, разрушенных в ходе тренировок, может потребоваться еще больший период времени, в течение которого уровень гликогена в мышцах уже может вернуться к исходному уровню. В этой связи хочу сразу обратить внимание читателей на тот факт, что заявления многих «гуру» бодибилдинга о том, что время восстановления мышцы после тренировки должно составлять n часов/дней, без указания о восстановлении какой ведущей функции идет речь, кажутся довольно сомнительными. Задать определенный период отдыха между тренировками, позволяющий получать развитие всех тренируемых функций одновременно, невозможно.Поэтому в классической спортивной школе годичный (и даже многолетний) период тренировок разбивают на микро и макроциклы в ходе которых ставятся задачи по развитию определенных тренируемых качеств, чередование тренировочных занятий в ходе микроциклов осуществляется таким образом, что бы физические нагрузки, направленные на развитие определенного двигательного качества задавались через промежутки времени, обеспечивающие суперкомпенсацию ведущей функции, а нагрузки иной направленности, применяемые в этот период, не оказывали отрицательного влияния на восстановление основной функции. Однако такой метод срабатывает только при развитии взаимонезависимых функций или параметров. В случае если определенное двигательное качество в равной мере зависит от развития нескольких функций или параметров, испытывающих одновременное напряжение в ходе одного тренировочного занятия и имеющих разное время восстановления, в течение микроцикла приходиться варьировать интенсивностью и объемом тренировок, накладывая волны восстановления различных параметров друг на друга таким образом, что бы получить суперкомпенсацию основных тренируемых функций к моменту завершения микроцикла.
Рис. 6.
На рисунке 6 представлен простейший вариант построения микроцикла для двух тренируемых функций, имеющих разное время восстановления. В течение микроцикла одна из функций испытывает последовательное положительное суммирование тренировочных эффектов, в то время как другая последовательно вводится в стадию истощения и достигает суперкомпенсации только во время отдыха, либо снижения нагрузки к концу микроцикла. Реальная картина тренировок еще гораздо сложнее, представьте, что тренируемых параметров и функций не две, а десятки. Но общие правила и закономерности, я думаю, вам уже стали понятны.Как вы видите, классическая теория тренировки оперирует такими понятиями как двигательные качества, функциональные возможности, и основывается на изучение процессов, приводящих к росту работоспособности мышц и организма в целом в различных режимах работы. Основой практически всех видов спорта является именно работоспособность, и цель планируемых адаптационных изменений в организме спортсмена, как правило, – выход на новый уровень работоспособности. Гипертрофия мышц вовсе не является целью тренинга в классическом спорте, и воспринимается лишь как побочный продукт развития основных двигательных качеств, более того, в некоторых случаях, гипертрофия мышц может оказывать даже отрицательное влияние на достижение стоящих перед спортсменом целей. Посетителей же тренажерных залов, за редким исключением, в большей степени интересует именно достижение гипертрофии мышц, нежели развитие двигательных качеств и повышения работоспособности. Безусловно, развитие работоспособности мышц способствует гипертрофии мышечной ткани.
Процессы, направленные на улучшение доставки кислорода к мышцам существенно развивают капиллярную сеть, что способствует общей гипертрофии мышц.
Тренировка окислительной активности мышц приводит к значительному росту в саркоплазме мышечных волокон количества и объема митохондрий – энергетических станций клетки.
Последовательные процессы суперкомпенсации внутримышечных запасов гликогена приводят к значительному его накоплению, что, в свою очередь, увеличивает объем саркоплазмы мышечного волокна.
Накопление иных веществ, ответственных за энергообеспечение, мышечной деятельности, таких, например, как креатинфосфат, так же увеличивает объем саркоплазмы, и даже не столько за счет объемов самих этих веществ, сколько за счет сопутствующего увеличения объема внутриклеточной жидкости.
То есть, тренировка работоспособности мышц приводит к гипертрофии мышечных волокон в основном за счет увеличения объема саркоплазмы.
Но самый существенный вклад в рост объемов и силы сокращения волокна вносит гипертрофия миофибрилл, все остальные компоненты клетки призваны лишь обеспечивать их сократительную активность. Как размер топливных баков самолета зависит от мощности его турбин, так и объем саркоплазмы мышечного волокна зависит от развития сократительного аппарата клетки (ну и конечно от объема работы, регулярно выполняемой мышцами).
Как я упоминал в первой части, миофибриллы представляют из себя белковые нити, поэтому увеличение количества и поперечного сечения миофибрилл в волокне напрямую связано с интенсивностью синтеза белка клеткой. Сейчас является неопровержимым фактом то, что тренировка интенсифицирует синтез белка, но вот вопрос, – как и почему это происходит?
Как строится белок
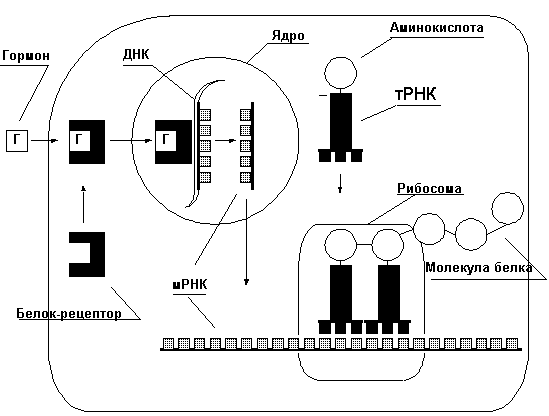
Молекула белка представляет из себя цепочку аминокислот, число звеньев в которой исчисляется от нескольких десятков до нескольких тысяч. Всего в природе насчитывается более 300 видов аминокислот, но для строительства белка используется только 20. Свойства белка определяются последовательностью аминокислот в цепочке, а так же пространственной конфигурацией самой цепочки (вторичная и третичная структура белка). Все белки человека строятся в клетках самостоятельно из аминокислот, поступающих в организм с белковой пищей и синтезируемых самим организмом. Упрощенно процесс синтеза белка изображен на рисунке 7.Рис. 7.
Код каждого белка записан в ДНК ядра клетки в виде цепочки нуклеотидов. Комбинация из трех нуклеотидов, называемая кодоном, кодирует одну аминокислоту. Последовательность кодонов в ДНК определяет последовательность аминокислот в белке. Ген – последовательность нуклеотидов, кодирующая один белок. Эта последовательность считывается с ДНК и записывается в матричной – РНК (м-РНК), этот процесс называется транскрипцией. мРНК – это как бы кусочек ДНК способный выходить из ядра в саркоплазму, где закрепляется на рибосомах. Транспортные РНК (тРНК) доставляют к мРНК аминокислоты. Один конец тРНК узнает на мРНК соответствующий кодон и прикрепляется к нему. Аминокислота, находящаяся на другом конце тРНК, сцепляется с аминокислотой соседней тРНК, таким образом, выстраивается цепочка белка.Синтез белка очень сложный процесс и его интенсивность зависит от огромного количества факторов.
Прежде всего, считывание мРНК в ядре начинается под воздействием стероидных гормонов, вырабатываемых железами внутренней секреции и поступающими в кровь. Молекула гормона из крови проникает в клетку, где, с помощью белка рецептора, доставляется в ядро и разблокирует участок цепочки ДНК, ответственный за тот или иной белок, после чего становится возможной транскрипция мРНК.
Для запуска транскрипции РНК, необходимо так же развернуть спираль ДНК, для чего используется фермент РНК-полимераза.
На синтез белка сильнейшее влияние оказывает гормон роста (СТГ или соматотропин). По химическому составу соматотропин это белок, поэтому он не может свободно проникать в клетку, в отличие от стероидных гормонов, а воздействует на рецепторы, расположенные на поверхности клетки. Механизм действия гормона роста до конца не изучен, но точно известно, что он стимулирует деятельность РНК-полимераз и рибосомного аппарата клетки.
Ну и конечно для сборки белка требуется наличие в клетке достаточного количества аминокислот и запасов энергии. Без аминокислот не из чего будет строить белок, а энергия нужна для сборки молекулы.
И так, для успешного синтеза белка требуются, как минимум, следующие условия:
– высокий уровень анаболических гормонов в крови (тестостерона и соматотропина)
– наличие в клетке белков-рецепторов тестостерона
– активность ферментов и факторов транскрипции РНК (РНК-полимераз и др.)
– достаточное количество аминокислот в клетке
– запас энергии в клетке
Теперь осталось только ответить на вопрос – как именно тренировка влияет на синтез белка?
Должен разочаровать читателя детально объяснить механизм этого влияния на сегодняшнем уровне развития науки невозможно. Если о том, как происходит регуляция синтеза белка в простейших одноклеточных организмах, когда в клетке может идти строительство всех белков, закодированных в ДНК, ученые имеют определенное представление, то, как осуществляется регуляция синтеза белка в многоклеточных организмах, когда, теоретически, каждая клетка может синтезировать все возможные белки, закодированные в ДНК, но синтезирует лишь набор белков, присущий данному типу клеток, остается пока не известным. Да, гормоно-рецепторный комплекс разблокирует участок ДНК, в котором закодирован определенный белок, но как гормон узнает, какой именно ген в данный момент необходим клетке – миозин быстрого волокна или миозин медленного волокна, а может быть миоглобин? Генетикам предстоит еще долгий путь, прежде чем раскроются все тайны синтеза белка. А как же быть до тех пор?