Страница:
На этом я, пожалуй, завершу рассмотрение тренировочных методов, воздействующих на факторы общей выносливости организма, и перейду к рассмотрению, собственно, силовой выносливости мышц.
Способность мышц сокращаться с требуемым усилием определяется, прежде всего, насыщенностью мышц энергией. И хотя основной причиной снижения силы сокращения мышц является вовсе не отсутствие АТФ, а снижение АТФазной активности миозина и нарушения в механизме передачи возбуждения с нерва вглубь волокна, причиной упомянутых нарушений являются метаболические факторы утомления (молочная кислота, ортофосфорная кислота, АДФ и др.), а их появление в мышце связано как раз с доступностью энергии. Недостаток АТФ, производимой окислительным путем, приводит к активизации гликолиза и появлению в мышце большого количества молочной кислоты (лактата), недостаток энергии, производимой путем гликолиза, приводит к истощению запасов креатинфосфата и, соответственно, увеличению в мышце концентрации ортофосфата.
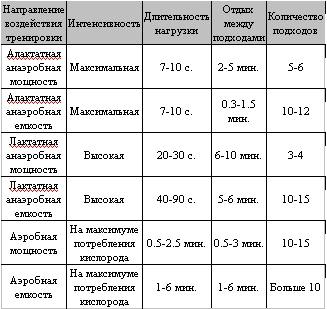
По Н.И. Волкову при рассмотрении факторов работоспособности мышц, в зависимости от основного механизма энергообеспечения, следует различать аэробную (окисление) и анаэробную работоспособность, а анаэробная работоспособность, в свою очередь, делится на лактатную (гликолиз) и алактатную (креатинфосфат). В качестве главных критериев оценки механизмов энергообеспечения мышечной деятельности принято выделять максимальную мощность, время удержания максимальной мощности, и общую емкость механизма. Максимальная мощность – это наибольшая скорость образования АТФ в данном метаболическом процессе. От мощности механизма энергообеспечения зависит возможная сила сокращения мышц в данном режиме работы. Под емкостью понимается общее количество энергии, которое можно получить за счет данного механизма ресинтеза АТФ.
Из вышесказанного следует, что для запуска адаптационных процессов, направленных на увеличение максимальной гликолитической мощности, длительность нагрузки должна соответствовать времени удержания максимальной мощности данного метаболического процесса, что составляет 30-60 секунд. Отдых между подходами должен быть достаточно длительным, для обеспечения вывода продуктов метаболизма из мышцы и развития высокой мощности гликолиза в следующем подходе. Устойчивость рН среды мышечных волокон к выбросу молочной кислоты и устойчивость ключевых ферментов к снижению рН вырабатывается в ходе тренировок, сопровождающихся максимальным накоплением лактата в мышцах. Это могут быть нагрузки высокой интенсивности, длительностью 1-1.5 минуты до наступления отказа мышц, вызванного сильным закислением, либо более короткие нагрузки, длительностью 20-40 секунд, со столь же коротким интервалом отдыха, приводящие к кумулятивному накоплению лактата в мышцах.
Гликолитическая емкость определяется главным образом запасами гликогена в мышцах, гликоген печени для процессов гликолиза не обладает достаточной мобильностью. О методах накопления мышечного гликогена, как и гликогена печени, я уже рассказывал при рассмотрении факторов общей работоспособности организма.
В заключение хочу привести сводную таблицу тренировочного воздействия на работоспособность мышц в различных режимах работы, почерпнутую мной из диссертации М. Хосни, посвященной изучению биохимических основ интервальной тренировки. Для развития соответствующих качеств Хосни рекомендует следующие методические приемы:
 На этом я заканчиваю изложение основ тренировки работоспособности мышц и перехожу к анализу основных факторов, определяющих мышечные объемы спортсмена.
На этом я заканчиваю изложение основ тренировки работоспособности мышц и перехожу к анализу основных факторов, определяющих мышечные объемы спортсмена.
Часть 6: Рассмотрение тренировочных методик.
Способность мышц сокращаться с требуемым усилием определяется, прежде всего, насыщенностью мышц энергией. И хотя основной причиной снижения силы сокращения мышц является вовсе не отсутствие АТФ, а снижение АТФазной активности миозина и нарушения в механизме передачи возбуждения с нерва вглубь волокна, причиной упомянутых нарушений являются метаболические факторы утомления (молочная кислота, ортофосфорная кислота, АДФ и др.), а их появление в мышце связано как раз с доступностью энергии. Недостаток АТФ, производимой окислительным путем, приводит к активизации гликолиза и появлению в мышце большого количества молочной кислоты (лактата), недостаток энергии, производимой путем гликолиза, приводит к истощению запасов креатинфосфата и, соответственно, увеличению в мышце концентрации ортофосфата.
По Н.И. Волкову при рассмотрении факторов работоспособности мышц, в зависимости от основного механизма энергообеспечения, следует различать аэробную (окисление) и анаэробную работоспособность, а анаэробная работоспособность, в свою очередь, делится на лактатную (гликолиз) и алактатную (креатинфосфат). В качестве главных критериев оценки механизмов энергообеспечения мышечной деятельности принято выделять максимальную мощность, время удержания максимальной мощности, и общую емкость механизма. Максимальная мощность – это наибольшая скорость образования АТФ в данном метаболическом процессе. От мощности механизма энергообеспечения зависит возможная сила сокращения мышц в данном режиме работы. Под емкостью понимается общее количество энергии, которое можно получить за счет данного механизма ресинтеза АТФ.
Алактатная работоспособность мышц
Максимальная алактатная мощность, с одной стороны, зависит от концентрации и активности фермента креатинкиназа (переносящего фосфатную группу с креатинфосфата на АДФ) и собственно креатинфосфата, с другой стороны мощность данной реакции зависит от потребности мышц в энергии, соответственно, определяется максимальной скоростью расхода АТФ развиваемой мышцами. Максимальная длительность удержания алактатной мощности составляет 6-12 секунд. Алактатная емкость зависит от запасов креатинфосфата в мышце. О методах тренировки алактатной мощности и емкости я уже рассказывал ранее, рассматривая методы развития максимальной силы, и сейчас не буду подробно останавливаться на этом вопросе.Лактатная работоспособность мышц
Максимальная лактатная мощность определяется главным образом концентрацией и активностью ключевых ферментов гликолиза. Время удержания максимальной мощности данного метаболического процесса составляет 30-60 секунд, и определяется, с одной стороны, устойчивостью ферментов гликолиза к понижению рН среды (повышение кислотности среды ингибирует активность гликолитических ферментов, что подавляет энергопроизводство), и устойчивостью кислотно-щелочного равновесия внутренней среды мышц, в условиях усиленной выработки лактата. С другой стороны, время удержания максимальной гликолитической мощности лимитируется факторами утомления мышцы, снижающими интенсивность сокращения.Из вышесказанного следует, что для запуска адаптационных процессов, направленных на увеличение максимальной гликолитической мощности, длительность нагрузки должна соответствовать времени удержания максимальной мощности данного метаболического процесса, что составляет 30-60 секунд. Отдых между подходами должен быть достаточно длительным, для обеспечения вывода продуктов метаболизма из мышцы и развития высокой мощности гликолиза в следующем подходе. Устойчивость рН среды мышечных волокон к выбросу молочной кислоты и устойчивость ключевых ферментов к снижению рН вырабатывается в ходе тренировок, сопровождающихся максимальным накоплением лактата в мышцах. Это могут быть нагрузки высокой интенсивности, длительностью 1-1.5 минуты до наступления отказа мышц, вызванного сильным закислением, либо более короткие нагрузки, длительностью 20-40 секунд, со столь же коротким интервалом отдыха, приводящие к кумулятивному накоплению лактата в мышцах.
Гликолитическая емкость определяется главным образом запасами гликогена в мышцах, гликоген печени для процессов гликолиза не обладает достаточной мобильностью. О методах накопления мышечного гликогена, как и гликогена печени, я уже рассказывал при рассмотрении факторов общей работоспособности организма.
Аэробная работоспособность мышц
Максимальная аэробная мощность зависит главным образом от плотности митохондрий в мышечных волокнах, концентрации и активности окислительных ферментов, скорости поступления кислорода вглубь волокна. Объем кислорода доступного для окислительных реакций лимитируется, как факторами общей работоспособности организма, которые я уже ранее рассматривал, так и рядом локальных внутримышечных факторов, среди которых можно выделить капиляризацию мышц, концентрацию миоглобина, диаметр мышечного волокна (чем меньше диаметр волокна, тем лучше оно снабжается кислородом и тем выше его относительная аэробная мощность). Скорость производства АТФ за счет окисления достигает максимальных значений на 2-3-й минуте работы, что связано с необходимостью развертывания множества процессов, обеспечивающих доставку кислорода к митохондриям. Время удержания максимальной аэробной мощности составляет примерно 6 минут, в дальнейшем аэробная мощность снижается по причине усталости всех активно работающих систем организма. Соответственно, для повышения аэробной мощности мышц тренировочная нагрузка должна длиться не менее 2 минут (для выхода скорости энергопроизводства на максимум). Не имеет смысла и затягивать нагрузку дольше чем на 6 минут, при тренировке именно мощности, так как далее идет ее (мощности) снижение. Эффективным оказывается многократное повторение таких нагрузок.В заключение хочу привести сводную таблицу тренировочного воздействия на работоспособность мышц в различных режимах работы, почерпнутую мной из диссертации М. Хосни, посвященной изучению биохимических основ интервальной тренировки. Для развития соответствующих качеств Хосни рекомендует следующие методические приемы:
Часть 6: Рассмотрение тренировочных методик.
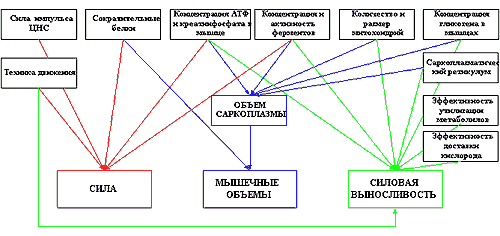
Ну что же, основные методы тренировок, способствующих развитию силы и силовой выносливости мышц, нами уже рассмотрены. Настало время приступить к рассмотрению тренировочных методик, в полной мере способствующих гипертрофии мышц, для чего следует определить тканевые и внутриклеточные структуры, от развития которых зависят мышечные объемы спортсмена. Я уже затрагивал немного этот вопрос во второй части, теперь остановимся на нем чуть подробнее. Как вы помните, объем мышцы, прежде всего, определяется количеством мышечных волокон (клеток) в теле мышцы, размером самих этих волокон, а так же объемом межклеточного вещества, представленного, главным образом, кровеносными сосудами и соединительной тканью, отделяющей друг от друга отдельные волокна и их пучки. Немаловажное значение для визуальных объемов спортсмена имеют и запасы жира в организме, однако, вклад жиров уже трудно назвать вкладом в «мышечные» объемы, а стандарты соревновательного бодибилдинга требует минимизации такового вклада, поэтому методы тренировок, приводящих к увеличению жировой составляющей объемов спортсмена, я рассматривать не буду, они и так хорошо всем известны. Увеличение количества мышечных волокон у человека ни разу достоверно не фиксировалось в экспериментах, хотя, как я уже говорил ранее, гиперплазия не кажется мне столь уж невероятным явлением, после того так она была зафиксирована у животных, но, дабы не прослыть пустым фантазером, я не буду включать гиперплазию в причины мышечной гипертрофии, до появления достоверных экспериментов, зафиксировавших увеличение количества мышечных волокон у человека. И так, нам остается уповать только на капилляризацию мышц, увеличение в объеме мышечных волокон и рост соединительной ткани. Объем мышечных волокон контролируется, прежде всего, количеством мышечных ядер в волокне. Именно от количества ядер, при прочих равных условиях, зависит общий объем белка, синтезируемого мышечным волокном в единицу времени. И этот фактор незаслуженно игнорируется многими специалистами при рассмотрении причин мышечной гипертрофии под действием тренировок. Как вы помните, к увеличению количества мышечных ядер приводит деление клеток сателлит, инициируемое факторами, появляющимися в мышечном волокне при его повреждении. Но ядра служат первопричиной, а увеличивают объем волокна иные клеточные структуры, такие как миофибриллы, саркоплазма, митохондрии, и др. Вот какие данные о потенциале роста мышц, за счет различных клеточных и межклеточных структур приводит в Ф. Хетфилд:
Факторы. Примерный вклад в увеличение размеров мышцы, %:
Капилляризация 3-5
Митохондрии 15-25
Саркоплазма (клеточная жидкость) 20-30
Соединительные ткани 2-3
Мышечные фибриллы 20-30
Гликоген 2-5
Как вы видите, существенный вклад в объем мышц вносит количество и поперечное сечение миофибрилл в мышечном волокне. Сравнимое влияние на размеры мышц оказывает объем саркоплазмы и расположенных в ней митохондрий. Таким образом, следует различать миофибриллярную и саркоплазматическую гипертрофию. На первый взгляд потенциал саркоплазматической гипертрофии (клеточная жидкость + митохондрии + гликоген) даже превышает потенциал роста за счет сократительных структур, но при внимательном рассмотрении становится ясно, что саркоплазматическая гипертрофия находятся в подчиненном отношении к миофибриллярной. Каждая миофибрилла требует наличия в клетке определенного объема саркоплазмы и митохондрий, призванных обеспечивать их (миофибрилл) функционирование. Рост миофибриллярных структур автоматически приведет к соответствующему увеличению саркоплазматических структур. Более того, процентные отношения, приведенные Хетфилдом вызывают некоторые сомнения, тем более что автор не указывает источник своей информации. Так, например, в учебнике биологической химии, за авторством Т.Т. Березова и Б.Ф. Коровкина приводятся несколько иные сведения. Химический анализ мышечной ткани показывает, что 70-80% мышечной массы приходится на воду и 20-30 это сухой остаток, состоящий из белков, липидов и углеводов. Процентное соотношение белков обнаруживаемых в сухом остатке следующее: сократительные белки – 35%, белки саркоплазмы – 45% и белки стромы (соединительной ткани) – 20%. То есть, процентные отношения белков получаются близкие к приведенным Хетфилдом, однако не следует забывать, что это отношения массы, а не объема. По данным этого же источника, миофибриллы занимают около 80% объема мышечного волокна, то есть на все остальные структуры помимо собственно миофибрилл, суммарно приходится не более 20% объема клетки. Соответственно, соотношение между миофибриллярной и саркоплазматической гипертрофией получается уже несколько иное, чем следует из данных приведенных Хетфилдом: миофибриллярная гипертрофия может дать до 80% прироста объемов волокна, а саркоплазматическая гипертрофия только 20%. Но для человека, стремящегося к максимальному развитию мускулатуры, не следует пренебрегать и этими 20-тью процентами.
Понятно, что относительный объем саркоплазмы мышечной клетки зависит и от активности использования миофибрилл, то есть, от объема регулярно производимой мышцами работы. Связь между концентрацией митохондрий в клетке с ее энергетическими потребностями, я думаю, не вызывает вопросов, а вот почему увеличение энергозатрат увеличивает объем саркоплазмы мышечной клетки, стоит разъяснить. Саркоплазма – это не только клеточная жидкость (вода), это и миллионы молекул различных веществ, взвешенных и растворенных в ней. Это, прежде всего, крупные молекулы белков-ферментов, призванные обеспечивать протекание множества жизненно важных химических реакций, в том числе и энергообеспечивающих. Это запасы органического топлива – АТФ, креатинфосфата, гликогена, жирных кислот и аминокислот. Это молекулы миоглобина. Это, в конце концов, всевозможные ионы (К+, Са++, Na+, Mg++ и др.). Но основной объем саркоплазмы создают даже не сами перечисленные вещества, а вода их окружающая. Вещества, растворенные и взвешенные в саркоплазме, одним своим наличием связывают, и задерживают в клетке определенное количество молекул воды. Накопление в клетке перечисленных выше веществ пропорционально увеличивает объем саркоплазмы. С эффектом резкого увеличения объема саркоплазмы мышечных волокон во время тренировки, за счет жидкости, нахлынувшей в клетки из межклеточного пространства и плазмы крови, вы хорошо знакомы. При гликолизе, активизирующемся во время мышечной деятельности, глюкоза распадается на молочную кислоту в соотношении 1:2 (одна молекула глюкозы – две молекулы молочной кислоты). Так как две молекулы кислоты связывают больше молекул воды, чем одна молекула глюкозы, то активизация гликолиза увеличивает потребность клетки в жидкости, и вода устремляется внутрь мышечных волокон, что приводит к их разбуханию и заметному увеличению мышц в объеме. Однако, не следует путать такое временное увеличение объемов с мышечным ростом, как только молочная кислота будет выведена из мышц, объем клеточной жидкости вернется к норме. Гипертрофией саркоплазмы можно считать лишь устойчивое увеличение ее объемов, фиксируемое в состоянии покоя.
Интересно, что увеличение объема саркоплазмы может происходить не только благодаря простому накоплению в ней перечисленных выше веществ. Саркоплазма мышечных волокон несколько отличается от саркоплазмы иных клеток, связно это с наличием в мышечных волокнах таких структур как миофибриллы. Каждая миофибрилла окружена плотной сетью саркоплазматического ретикулума, состоящей из терминальных цистерн с ионами Ca++ (ионы выбрасываются в саркоплазму при сокращении), и переплетения так называемых Т-трубочек, связывающих терминальные цистерны с сарколеммой (оболочкой волокна) и обеспечивающих поступление сигнала к сокращению. То есть, каждая миофибрилла жестко окружена определенным объемом саркоплазматических структур. Объем этих структур пропорционален площади поверхности миофибрилл в волокне. Соответственно, чем больше диаметр отдельных миофибрилл, тем меньше объем окружающей миофибриллу саркоплазмы по отношению к объему сократительных белков внутри этой миофибриллы (выше доля сократительных белков в волокне). Но, чем больше объем каждой миофибриллы, тем труднее обеспечивать ее энергопотребности, так как длиннее путь переноса энергии от поверхности миофибриллы (где расположены основные источники энергии – митохондрии) вглубь. Соответственно, при активизации мышечной деятельности адаптация волокон к изменению условий жизнедеятельности может быть направлена на расщепление крупных миофибрилл на несколько мелких. В случае расщепления миофибрилл их масса остается неизменной, однако, возрастает их количество и, соответственно, увеличивается площадь поверхности миофибрилл, что неминуемо должно сопровождаться увеличением объема саркоплазматического ретикулума. То есть, происходит гипертрофия мышечной клетки без увеличения объема сократительных белков, – наблюдается саркоплазматическая гипертрофия. Имея в виду, что объем саркоплазмы мышечного волокна может быть увеличен как благодаря накоплению различных веществ, ответственных за энергопроизводство клетки, так и благодаря расщеплению миофибрилл в процессе эргономического приспособления к увеличивающемуся объему работы, можно сказать, что саркоплазматическая гипертрофия является адаптационной реакцией мышц на увеличение объема работы, регулярно выполняемого мышцами.
Из вышеприведенного краткого анализа становится ясно, что никаких особых специфических методов тренировки, направленных исключительно на увеличение объемов мышц, не существует. Гипертрофии мышц, в той или иной мере, способствуют рассмотренные ранее тренировочные методики, направленные на развитие силы (за счет развития сократительных структур) и силовой выносливости мышц (саркоплазматическая гипертрофия). Точнее, развитие ряда клеточных структур, может вносить свой вклад в развитие таких мышечных качеств как сила, силовая выносливость и объем (см. рис.1)
Снижение тренируемой функции под действием тренировки —> восстановление (компенсация) —> сверхвосстановление (суперкомпенсация) —> утраченное сверхвосстановление (декомпенсация).
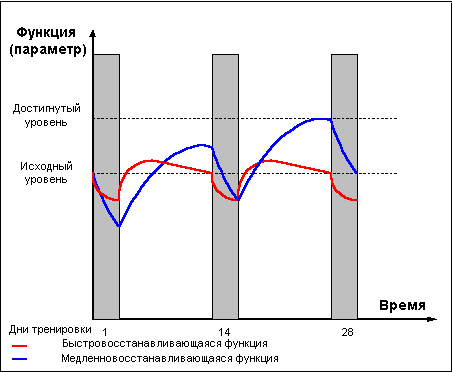
Положительное взаимодействие тренировочных эффектов наблюдается лишь при задании новой тренировочной нагрузки в состоянии сверхвосстановления (возросших функциональных возможностей). Слишком большой перерыв между тренировками приводит к воздействию на тренируемую функцию в состоянии утраченной компенсации и не может привести к закреплению адаптационных изменений, вызванных предыдущими тренировками. Недостаточный отдых между тренировками приводит к тому, что нагрузка на тренируемую функцию задается еще до того, как функция восстановится после предыдущей тренировки, что, при длительном повторении такой ситуации, может стать причиной перетренированности. Поэтому тренировочный процесс, по возможности, строится таким образом, что бы в период восстановления одной тренируемой функции задаваемая нагрузка воздействовала на иную систему организма и не оказывала отрицательного воздействия на восстанавливаемую функцию. Например, после объемной тренировки аэробной направленности восстановление энергетического потенциала мышц может занять около трех дней, в это время вполне возможно проведение небольших по объему тренировочных занятий анаэробного характера. Но не всегда тренировки можно так четко разделить по направленности тренировочного воздействия. При одновременном напряжении различных функциональных систем, обладающие свойством гетерохронизма восстановительных процессов, указать оптимальный перерыв между тренировочными нагрузками становится невозможно. Какое бы время отдыха вы не выбрали, оно будет оптимальным для одних функциональных систем, но часть функций окажется либо в состоянии недовосстановления, либо в состоянии утраченной компенсации. В той или иной мере любая тренировка воздействует на системы ответственные и за алактатные, и за гликолитические, и за аэробные процессы в мышцах и организме, более того, достаточно интенсивная тренировка в анаэробном режиме, как правило, оказывает разрушающее воздействие на сократительные структуры мышечных волокон, а интенсивная аэробная тренировка может оказать аналогичное разрушающее воздействие на митохондрии мышечных клеток. Восстановление клеточных структур требует больше времени, чем необходимо для достижения суперкомпенсации энергетических компонент, в свою очередь, различные энергетические компоненты так же имеют различные сроки достижения состояния суперкомпенсации. Как же быть?
С самым простым решением проблемы вы уже знакомы. Достаточно проводить тренировки с частотой, обеспечивающей достижение состояния суперкомпенсации самой долговосстанавливающейся функциональной системы. Такие тренировки обеспечат планомерный рост данной функции и позволят гарантировано избежать состояния перетренированности, так как ни одна функциональная система не будет нагружаться в состоянии недовосстановления. В то же время, ряд функциональных систем к моменту новой тренировки будет в состоянии декомпенсации, что не может привести к развитию долговременной адаптации данных систем (см. рис. 2). Как вы поняли, я веду речь о тренировках в стиле «Супертренинга» Майка Ментцера. Данный метод подходит только для развития силы и массы мышц за счет роста сократительных структур клетки (именно восстановление и суперкомпенсация сократительных белков клетки, разрушенных в ходе тренировки, требует наибольшего времени). Но хотя развитие миофибриллярного аппарата мышечных волокон является определяющим для силы и массы мышц, добиться максимально возможного развития силы, существенной саркоплазматической гипертрофии, значительного повышения силовой выносливости и общей работоспособности организма таким методом тренировки невозможно.
Факторы. Примерный вклад в увеличение размеров мышцы, %:
Капилляризация 3-5
Митохондрии 15-25
Саркоплазма (клеточная жидкость) 20-30
Соединительные ткани 2-3
Мышечные фибриллы 20-30
Гликоген 2-5
Как вы видите, существенный вклад в объем мышц вносит количество и поперечное сечение миофибрилл в мышечном волокне. Сравнимое влияние на размеры мышц оказывает объем саркоплазмы и расположенных в ней митохондрий. Таким образом, следует различать миофибриллярную и саркоплазматическую гипертрофию. На первый взгляд потенциал саркоплазматической гипертрофии (клеточная жидкость + митохондрии + гликоген) даже превышает потенциал роста за счет сократительных структур, но при внимательном рассмотрении становится ясно, что саркоплазматическая гипертрофия находятся в подчиненном отношении к миофибриллярной. Каждая миофибрилла требует наличия в клетке определенного объема саркоплазмы и митохондрий, призванных обеспечивать их (миофибрилл) функционирование. Рост миофибриллярных структур автоматически приведет к соответствующему увеличению саркоплазматических структур. Более того, процентные отношения, приведенные Хетфилдом вызывают некоторые сомнения, тем более что автор не указывает источник своей информации. Так, например, в учебнике биологической химии, за авторством Т.Т. Березова и Б.Ф. Коровкина приводятся несколько иные сведения. Химический анализ мышечной ткани показывает, что 70-80% мышечной массы приходится на воду и 20-30 это сухой остаток, состоящий из белков, липидов и углеводов. Процентное соотношение белков обнаруживаемых в сухом остатке следующее: сократительные белки – 35%, белки саркоплазмы – 45% и белки стромы (соединительной ткани) – 20%. То есть, процентные отношения белков получаются близкие к приведенным Хетфилдом, однако не следует забывать, что это отношения массы, а не объема. По данным этого же источника, миофибриллы занимают около 80% объема мышечного волокна, то есть на все остальные структуры помимо собственно миофибрилл, суммарно приходится не более 20% объема клетки. Соответственно, соотношение между миофибриллярной и саркоплазматической гипертрофией получается уже несколько иное, чем следует из данных приведенных Хетфилдом: миофибриллярная гипертрофия может дать до 80% прироста объемов волокна, а саркоплазматическая гипертрофия только 20%. Но для человека, стремящегося к максимальному развитию мускулатуры, не следует пренебрегать и этими 20-тью процентами.
Понятно, что относительный объем саркоплазмы мышечной клетки зависит и от активности использования миофибрилл, то есть, от объема регулярно производимой мышцами работы. Связь между концентрацией митохондрий в клетке с ее энергетическими потребностями, я думаю, не вызывает вопросов, а вот почему увеличение энергозатрат увеличивает объем саркоплазмы мышечной клетки, стоит разъяснить. Саркоплазма – это не только клеточная жидкость (вода), это и миллионы молекул различных веществ, взвешенных и растворенных в ней. Это, прежде всего, крупные молекулы белков-ферментов, призванные обеспечивать протекание множества жизненно важных химических реакций, в том числе и энергообеспечивающих. Это запасы органического топлива – АТФ, креатинфосфата, гликогена, жирных кислот и аминокислот. Это молекулы миоглобина. Это, в конце концов, всевозможные ионы (К+, Са++, Na+, Mg++ и др.). Но основной объем саркоплазмы создают даже не сами перечисленные вещества, а вода их окружающая. Вещества, растворенные и взвешенные в саркоплазме, одним своим наличием связывают, и задерживают в клетке определенное количество молекул воды. Накопление в клетке перечисленных выше веществ пропорционально увеличивает объем саркоплазмы. С эффектом резкого увеличения объема саркоплазмы мышечных волокон во время тренировки, за счет жидкости, нахлынувшей в клетки из межклеточного пространства и плазмы крови, вы хорошо знакомы. При гликолизе, активизирующемся во время мышечной деятельности, глюкоза распадается на молочную кислоту в соотношении 1:2 (одна молекула глюкозы – две молекулы молочной кислоты). Так как две молекулы кислоты связывают больше молекул воды, чем одна молекула глюкозы, то активизация гликолиза увеличивает потребность клетки в жидкости, и вода устремляется внутрь мышечных волокон, что приводит к их разбуханию и заметному увеличению мышц в объеме. Однако, не следует путать такое временное увеличение объемов с мышечным ростом, как только молочная кислота будет выведена из мышц, объем клеточной жидкости вернется к норме. Гипертрофией саркоплазмы можно считать лишь устойчивое увеличение ее объемов, фиксируемое в состоянии покоя.
Интересно, что увеличение объема саркоплазмы может происходить не только благодаря простому накоплению в ней перечисленных выше веществ. Саркоплазма мышечных волокон несколько отличается от саркоплазмы иных клеток, связно это с наличием в мышечных волокнах таких структур как миофибриллы. Каждая миофибрилла окружена плотной сетью саркоплазматического ретикулума, состоящей из терминальных цистерн с ионами Ca++ (ионы выбрасываются в саркоплазму при сокращении), и переплетения так называемых Т-трубочек, связывающих терминальные цистерны с сарколеммой (оболочкой волокна) и обеспечивающих поступление сигнала к сокращению. То есть, каждая миофибрилла жестко окружена определенным объемом саркоплазматических структур. Объем этих структур пропорционален площади поверхности миофибрилл в волокне. Соответственно, чем больше диаметр отдельных миофибрилл, тем меньше объем окружающей миофибриллу саркоплазмы по отношению к объему сократительных белков внутри этой миофибриллы (выше доля сократительных белков в волокне). Но, чем больше объем каждой миофибриллы, тем труднее обеспечивать ее энергопотребности, так как длиннее путь переноса энергии от поверхности миофибриллы (где расположены основные источники энергии – митохондрии) вглубь. Соответственно, при активизации мышечной деятельности адаптация волокон к изменению условий жизнедеятельности может быть направлена на расщепление крупных миофибрилл на несколько мелких. В случае расщепления миофибрилл их масса остается неизменной, однако, возрастает их количество и, соответственно, увеличивается площадь поверхности миофибрилл, что неминуемо должно сопровождаться увеличением объема саркоплазматического ретикулума. То есть, происходит гипертрофия мышечной клетки без увеличения объема сократительных белков, – наблюдается саркоплазматическая гипертрофия. Имея в виду, что объем саркоплазмы мышечного волокна может быть увеличен как благодаря накоплению различных веществ, ответственных за энергопроизводство клетки, так и благодаря расщеплению миофибрилл в процессе эргономического приспособления к увеличивающемуся объему работы, можно сказать, что саркоплазматическая гипертрофия является адаптационной реакцией мышц на увеличение объема работы, регулярно выполняемого мышцами.
Из вышеприведенного краткого анализа становится ясно, что никаких особых специфических методов тренировки, направленных исключительно на увеличение объемов мышц, не существует. Гипертрофии мышц, в той или иной мере, способствуют рассмотренные ранее тренировочные методики, направленные на развитие силы (за счет развития сократительных структур) и силовой выносливости мышц (саркоплазматическая гипертрофия). Точнее, развитие ряда клеточных структур, может вносить свой вклад в развитие таких мышечных качеств как сила, силовая выносливость и объем (см. рис.1)
Рис. 1.
Как вы понимаете, для максимального развития силы, выносливости, объемов мышц, следует использовать тренировки, воздействующие на все основные факторы, вносящие свой вклад в развитие соответствующих качеств.Взаимодействие срочных тренировочных эффектов
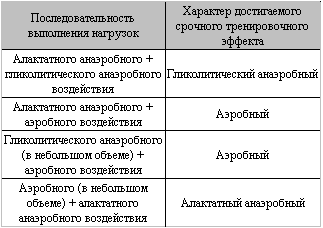
Рассмотрим, каким образом различные тренировочные методики можно объединить в единый тренировочный процесс. Основная проблема здесь состоит в том, что не все методические приемы совместимы в ходе одного тренировочного занятия. Адаптационные изменения в организме могут быть усилены либо ослаблены под влиянием нагрузок разной направленности. Этот феномен принято называть взаимодействием срочных тренировочных эффектов. Взаимодействие тренировочных эффектов разнонаправленных нагрузок может быть положительным, отрицательным и нейтральным. Наиболее выраженные адаптационные изменения в организме формируются при положительном взаимодействии тренировочных эффектов. При неправильном построении тренировок отрицательное взаимодействие нагрузок существенно снижает общий тренировочный эффект и даже может привести к перетренированности. По данным исследований, проведенных Волковым Н.И., положительный эффект от взаимодействия разнонаправленных нагрузок может быть достигнут только в ограниченном числе их сочетаний (см. таблицу).Взаимодействие отставленных тренировочных эффектов
Следующая важная проблема, возникающая при построении тренировочного процесса, кроется в различном времени, требующемся для восстановления нагружаемых в ходе тренировки функциональных систем организма. Хорошо известно, что быстрее всего после тренировки восстанавливаются алактатные резервы организма, чуть дольше происходит восстановление функциональных систем, ответственных за мощность и емкость гликолитических процессов (до 3-х суток), больше всего времени требуется на восстановление разрушенных в ходе тренировки клеточных структур. Данный феномен получил название гетерохронизма восстановительных процессов. Здесь я рекомендую обратиться ко второй части и вспомнить последовательность развития восстановительных процессов в организме:Снижение тренируемой функции под действием тренировки —> восстановление (компенсация) —> сверхвосстановление (суперкомпенсация) —> утраченное сверхвосстановление (декомпенсация).
Положительное взаимодействие тренировочных эффектов наблюдается лишь при задании новой тренировочной нагрузки в состоянии сверхвосстановления (возросших функциональных возможностей). Слишком большой перерыв между тренировками приводит к воздействию на тренируемую функцию в состоянии утраченной компенсации и не может привести к закреплению адаптационных изменений, вызванных предыдущими тренировками. Недостаточный отдых между тренировками приводит к тому, что нагрузка на тренируемую функцию задается еще до того, как функция восстановится после предыдущей тренировки, что, при длительном повторении такой ситуации, может стать причиной перетренированности. Поэтому тренировочный процесс, по возможности, строится таким образом, что бы в период восстановления одной тренируемой функции задаваемая нагрузка воздействовала на иную систему организма и не оказывала отрицательного воздействия на восстанавливаемую функцию. Например, после объемной тренировки аэробной направленности восстановление энергетического потенциала мышц может занять около трех дней, в это время вполне возможно проведение небольших по объему тренировочных занятий анаэробного характера. Но не всегда тренировки можно так четко разделить по направленности тренировочного воздействия. При одновременном напряжении различных функциональных систем, обладающие свойством гетерохронизма восстановительных процессов, указать оптимальный перерыв между тренировочными нагрузками становится невозможно. Какое бы время отдыха вы не выбрали, оно будет оптимальным для одних функциональных систем, но часть функций окажется либо в состоянии недовосстановления, либо в состоянии утраченной компенсации. В той или иной мере любая тренировка воздействует на системы ответственные и за алактатные, и за гликолитические, и за аэробные процессы в мышцах и организме, более того, достаточно интенсивная тренировка в анаэробном режиме, как правило, оказывает разрушающее воздействие на сократительные структуры мышечных волокон, а интенсивная аэробная тренировка может оказать аналогичное разрушающее воздействие на митохондрии мышечных клеток. Восстановление клеточных структур требует больше времени, чем необходимо для достижения суперкомпенсации энергетических компонент, в свою очередь, различные энергетические компоненты так же имеют различные сроки достижения состояния суперкомпенсации. Как же быть?
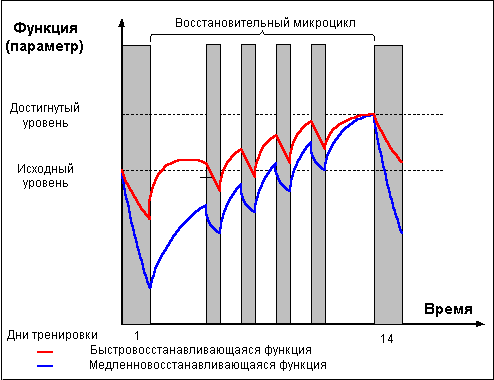
С самым простым решением проблемы вы уже знакомы. Достаточно проводить тренировки с частотой, обеспечивающей достижение состояния суперкомпенсации самой долговосстанавливающейся функциональной системы. Такие тренировки обеспечат планомерный рост данной функции и позволят гарантировано избежать состояния перетренированности, так как ни одна функциональная система не будет нагружаться в состоянии недовосстановления. В то же время, ряд функциональных систем к моменту новой тренировки будет в состоянии декомпенсации, что не может привести к развитию долговременной адаптации данных систем (см. рис. 2). Как вы поняли, я веду речь о тренировках в стиле «Супертренинга» Майка Ментцера. Данный метод подходит только для развития силы и массы мышц за счет роста сократительных структур клетки (именно восстановление и суперкомпенсация сократительных белков клетки, разрушенных в ходе тренировки, требует наибольшего времени). Но хотя развитие миофибриллярного аппарата мышечных волокон является определяющим для силы и массы мышц, добиться максимально возможного развития силы, существенной саркоплазматической гипертрофии, значительного повышения силовой выносливости и общей работоспособности организма таким методом тренировки невозможно.
Рис. 2.
Совместить мифибрилльную гипертрофию с повышением работоспособности мышц и, тем самым, увеличить долю саркоплазматической гипертрофии возможно посредством метода регулирования интенсивности нагрузки. Наибольшим воздействием на сократительные структуры мышечных волокон, как вы помните, обладают высокоинтенсивные тренировки, в ходе которых мышцы развивают относительно высокую скорость расхода энергии. Тренировки, обладающие таким разрушающим мышечные структуры эффектом, и запускающие соответствующие адаптационные процессы, следует проводить только, когда мышцы войдут в состояние суперкомпенсации, после полного восстановления разрушенных интенсивной тренировкой клеточных структур. Как показывает опыт, в зависимости от нагрузки и индивидуальных восстановительных особенностей спортсмена, для полного восстановления и достижения состояния суперкомпенсации мышцам может потребоваться до 2-3-х недель. Но в течение этого периода для обеспечения полноценного восстановления вовсе не обязательно абсолютно исключать нагрузку на мышцы. Для того чтобы мышцы не вошли в состояние декомпенсации энергетических функций, период отдыха между тяжелыми тренировками можно заполнить тренировками меньшей интенсивности, не оказывающими на мышцы разрушающего воздействия (либо сводящими такое воздействие к минимуму), в то же время способствующих закреплению адаптационных изменений в энергобеспечивающей системе мышц и организма в целом. В этом случае тренировочный процесс разбивается на мезоциклы, состоящие из двух микроциклов – ударного и восстановительного (см. рис. 3.).