Страница:
Аналогичные явления были продемонстрированы и на других позвоночных. Деафферентированные головастики, жабы и рыбы сохраняли способность плавать, нормально координируя движения, до тех пор, пока у них оставался неразрушенным хотя бы один чувствительный нерв. Следовательно, для поддержания плавательных движений необходим минимальный уровень афферентной импульсации, которая способствует поддержанию уровня возбуждения в соответствующих нервных центрах (или «специфической энергии действия» по Лоренцу).
Большое значение в этом плане имели работы Рёдера, который и обнаружил недостаточность рефлекторной теории для объяснения некоторых сторон поведения насекомых.
В последующие десятилетия эти данные о возможности спонтанной активности в цнс насекомых получили широкое экспериментальное подтверждение. По представлениям Рёдера, нервные элементы, лежащие в основе спонтанного и рефлекторного поведения, различаются, в частности, по порогу возбудимости. Порог раздражения первых может быть настолько низким, что инстинктивная реакция способна осуществляться помимо видимых внешних раздражителей.
Влияние уровня возбуждения в соответствующих нервных центрах на характер протекания инстинктивных реакций было подтверждено работами немецких ученых Э. фон Хольста и У. Сент-Поль (1960). В зависимости от локализации раздражаемой током структуры ствола мозга кур они наблюдали либо элементарные поведенческие реакции (клевание, повороты головы), либо сложные акты поведения (ухаживание за отсутствующими самками). Сложные реакции протекали менее константно, чем простые. Они зависели от состояния животного, от предшествующих стимулов от параметров тока и всей совокупности действующих на птицу стимулов. Например, при слабом токе петух бросался на чучело хорька, на которое до этого не обращал внимания, а при усилении тока такая же реакция проявлялась на неспецифический раздражитель — кулак.
Накопление «специфической энергии действия» представлено в модели в виде постепенного накопления воды в резервуаре.
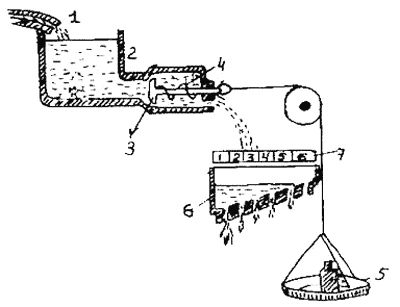
1 — кран, 2 — резервуар, 3 — клапан, 4 — пружина, 5 — чашка весов, 6 — лоток 7 — шкала
Давление жидкости внутри него все время повышается. Отток жидкости из резервуара (аналогизируемый с активностью животного) возможен по трубам, которые блокируются клапаном. Этот клапан может открыться при достаточно большом давлении (накоплении жидкости в резервуаре), а также при воздействии извне — при действии тяжести гири, приложенной к клапану. Такое внешнее воздействие аналогизируется с влиянием внешних раздражителей, специфичных для данного поведенческого акта. Нарастающее давление и действие внешней силы направлены одинаково и могут суммироваться. Чем больше жидкости набралось в резервуаре, тем меньше усилий нужно приложить извне, чтобы открыть клапан, а иногда клапан может открыться и без внешнего воздействия. Этот момент моделирует появление активности «вхолостую», возникающей без видимых внешних сигналов. Эта аналогия может быть продолжена. Если клапан открылся лишь немного, жидкость будет вытекать слабо и попадет только в область нижнего отверстия специального наклонного лотка. Если же клапан откроется сильнее — жидкость потечет сильнее и попадет и на следующие отверстия. Этот лоток с отверстиями аналогизируется с разными типами двигательной активности, относящейся к данному поведенческому акту. Самое нижнее отверстие соответствует активности с наиболее низким порогом — например, некоторым проявлением поискового поведения. Другие отверстия соответствуют иным формам активности, обладающим более высоким порогом. Если резервуар опорожняется, т. е. иссякает; «специфическая энергия действия», — данное поведение больше продолжаться не может.
Анализ применимости модели Лоренца к описанию поведения в сопоставлении с другой моделью, предложенной Дейчем, можно найти в книге Меннинга (1982). Плодотворность применения модели Лоренца мы иллюстрируем в гл.5 на примере анализа поведения кошек, выполненного Лейхаузеном.
Современная оценка концепции Лоренца о врожденном разрешающем механизме опирается на большое количество экспериментальных фактов, полученных в последние годы. Подробный обзор современных взглядов по этим вопросам приводится в книге Меннинга.
Очевидно, что и «специфическая энергия действия» и ключевые стимулы — это понятия, которые в переводе на язык современной нейрофизиологии выражаются такими терминами, как «специфическое побуждение», активация той или иной мотивационной системы, а также видоспецифическая избирательность перцепторного аппарата.
Развитие концепции К. Лоренца в работах Тинбергена
Иерархическая теория инстинкта
Конфликтное поведение
Разработка проблем эволюции поведения
Функциональный анализ признаков поведения
Большое значение в этом плане имели работы Рёдера, который и обнаружил недостаточность рефлекторной теории для объяснения некоторых сторон поведения насекомых.
В последующие десятилетия эти данные о возможности спонтанной активности в цнс насекомых получили широкое экспериментальное подтверждение. По представлениям Рёдера, нервные элементы, лежащие в основе спонтанного и рефлекторного поведения, различаются, в частности, по порогу возбудимости. Порог раздражения первых может быть настолько низким, что инстинктивная реакция способна осуществляться помимо видимых внешних раздражителей.
Влияние уровня возбуждения в соответствующих нервных центрах на характер протекания инстинктивных реакций было подтверждено работами немецких ученых Э. фон Хольста и У. Сент-Поль (1960). В зависимости от локализации раздражаемой током структуры ствола мозга кур они наблюдали либо элементарные поведенческие реакции (клевание, повороты головы), либо сложные акты поведения (ухаживание за отсутствующими самками). Сложные реакции протекали менее константно, чем простые. Они зависели от состояния животного, от предшествующих стимулов от параметров тока и всей совокупности действующих на птицу стимулов. Например, при слабом токе петух бросался на чучело хорька, на которое до этого не обращал внимания, а при усилении тока такая же реакция проявлялась на неспецифический раздражитель — кулак.
Гидравлическая модель.
Лоренц предложил гипотетическую модель осуществления реакций типа завершающих актов, общие принципы которой он заимствовал из гидравлики. В настоящее время гидравлическую модель можно считать глубоким анахронизмом в этологии, представляющим лишь исторический интерес. Ввиду этого мы остановимся на ней очень кратко (рис.1).Накопление «специфической энергии действия» представлено в модели в виде постепенного накопления воды в резервуаре.
Рис. 1. Схема гидравлической модели для объяснения регуляции поведения (см. Меннинг, 1982)
1 — кран, 2 — резервуар, 3 — клапан, 4 — пружина, 5 — чашка весов, 6 — лоток 7 — шкала
Давление жидкости внутри него все время повышается. Отток жидкости из резервуара (аналогизируемый с активностью животного) возможен по трубам, которые блокируются клапаном. Этот клапан может открыться при достаточно большом давлении (накоплении жидкости в резервуаре), а также при воздействии извне — при действии тяжести гири, приложенной к клапану. Такое внешнее воздействие аналогизируется с влиянием внешних раздражителей, специфичных для данного поведенческого акта. Нарастающее давление и действие внешней силы направлены одинаково и могут суммироваться. Чем больше жидкости набралось в резервуаре, тем меньше усилий нужно приложить извне, чтобы открыть клапан, а иногда клапан может открыться и без внешнего воздействия. Этот момент моделирует появление активности «вхолостую», возникающей без видимых внешних сигналов. Эта аналогия может быть продолжена. Если клапан открылся лишь немного, жидкость будет вытекать слабо и попадет только в область нижнего отверстия специального наклонного лотка. Если же клапан откроется сильнее — жидкость потечет сильнее и попадет и на следующие отверстия. Этот лоток с отверстиями аналогизируется с разными типами двигательной активности, относящейся к данному поведенческому акту. Самое нижнее отверстие соответствует активности с наиболее низким порогом — например, некоторым проявлением поискового поведения. Другие отверстия соответствуют иным формам активности, обладающим более высоким порогом. Если резервуар опорожняется, т. е. иссякает; «специфическая энергия действия», — данное поведение больше продолжаться не может.
Современная оценка гидравлической модели Лоренца.
В определенных пределах предложенная Лоренцем модель хорошо описывает феноменологию инстинктивных действий, а также циклические изменения, происходящие в реактивности нервной системы животного к внешним раздражителям — снижение порога совершения действия, если оно долго не выполнялось, восстановление готовности к инстинктивным действиям после перерыва и возможность появления реакций на неспецифические раздражители. В течение некоторого времени она являлась стимулом к развитию новых исследований. Однако наряду с этим модель Лоренца неоднократно подвергалась критике, отчасти необоснованной, но отчасти и справедливой (например, механистичность построения). При создании модели принимаются в расчет лишь самые общие черты, отражающие схему работы моделируемой системы, а также ее соответствие известным из опыта фактам. Другое дело, что предложенная Лоренцем модель, хорошо объясняя очень многие из известных фактов, объяснить другие была не в состоянии. Однако это свидетельствовало лишь о том, что представление о запасании «специфической энергии действия» имеет ограниченное применение, и соответствующие термины и понятия не универсальны.Анализ применимости модели Лоренца к описанию поведения в сопоставлении с другой моделью, предложенной Дейчем, можно найти в книге Меннинга (1982). Плодотворность применения модели Лоренца мы иллюстрируем в гл.5 на примере анализа поведения кошек, выполненного Лейхаузеном.
Современная оценка концепции Лоренца о врожденном разрешающем механизме опирается на большое количество экспериментальных фактов, полученных в последние годы. Подробный обзор современных взглядов по этим вопросам приводится в книге Меннинга.
Очевидно, что и «специфическая энергия действия» и ключевые стимулы — это понятия, которые в переводе на язык современной нейрофизиологии выражаются такими терминами, как «специфическое побуждение», активация той или иной мотивационной системы, а также видоспецифическая избирательность перцепторного аппарата.
Развитие концепции К. Лоренца в работах Тинбергена
Представления Лоренца, заложившего основы этологии, развил голландский ученый Н. Тиберген. В 50-е годы Тинберген был приглашен в Оксфордский университет, где и была проведена большая часть его исследований. Работы Тинбергена и его учеников образовали особое направление, которое известно как английская школы этологии.
Тинбергену принадлежит разработка иерархической модели поведения, которая в большей степени учитывала физиологические данные, чем исходная модель Лоренца. На базе этой модели он выделил некоторые формы конфликтного поведения и высказал гипотезу о их механизмах.
Тинберген и его ученики в течение многих лет систематически исследовали в природных условиях поведение ряда видов насекомых и птиц. Классическим объектом их лабораторных исследований стала трехиглая колюшка — легко содержащийся в неволе вид пресноводных рыб. Репродуктивное поведение колюшки послужило моделью для выявления многих важных принципов организации поведения животных. Особое значение приобрели их работы по колониальным морским птицам. Они явились основой многих современных представлений о сообществах животных и факторах, регулирующих их структуру, а также о коммуникативном значении ряда поведенческих актов. Кроме того, они способствовали изучению проблемы многообразных форм приспособления животных к борьбе с хищниками, которая накладывает отпечаток практически на все стороны поведения. Впоследствии эта проблема изучалась и на других животных, Последовательное применение сравнительного метода и остроумные полевые эксперименты способствовали выделению приспособительного значения поведенческих признаков и их эволюционного происхождения. Рассмотрим основные направления работ Тинбергена.
Тинбергену принадлежит разработка иерархической модели поведения, которая в большей степени учитывала физиологические данные, чем исходная модель Лоренца. На базе этой модели он выделил некоторые формы конфликтного поведения и высказал гипотезу о их механизмах.
Тинберген и его ученики в течение многих лет систематически исследовали в природных условиях поведение ряда видов насекомых и птиц. Классическим объектом их лабораторных исследований стала трехиглая колюшка — легко содержащийся в неволе вид пресноводных рыб. Репродуктивное поведение колюшки послужило моделью для выявления многих важных принципов организации поведения животных. Особое значение приобрели их работы по колониальным морским птицам. Они явились основой многих современных представлений о сообществах животных и факторах, регулирующих их структуру, а также о коммуникативном значении ряда поведенческих актов. Кроме того, они способствовали изучению проблемы многообразных форм приспособления животных к борьбе с хищниками, которая накладывает отпечаток практически на все стороны поведения. Впоследствии эта проблема изучалась и на других животных, Последовательное применение сравнительного метода и остроумные полевые эксперименты способствовали выделению приспособительного значения поведенческих признаков и их эволюционного происхождения. Рассмотрим основные направления работ Тинбергена.
Иерархическая теория инстинкта
Основой для разработанной Тинбергеном модели поведения послужили следующие факты. Известно, что между различными стереотипными двигательными реакциями существует ряд закономерных отношений. В некоторых ситуациях группы инстинктивных движений появляются совместно — они характеризуют определенное внутреннее состояние животного и проявляют общие флюктуации порога поведенческой реакции. Повышение порога реакции А поднимает порог реакции В (и наоборот), а это свидетельствует о том, что обе они зависят от общего функционального «центра». Наблюдая за сложными поведенческими комплексами действий, можно видеть некоторую регулярность в последовательности проявления тех или иных действий. В качестве примера можно привести агрессивные столкновения рыб за раздел территории. У многих костистых рыб, в том числе и у цихлид, им почти всегда предшествует демонстрация запугивания. Причем у одних видов эти столкновения следуют за очень коротким периодом запугивания; а у других — за весьма разнообразными демонстрациями запугивания, серьезное агрессивное столкновение с ранениями следует лишь в том случае, если силы обоих самцов равны. Наконец, у третьей группы видов настоящие драки уже не наблюдается, и крайне ритуализированная церемония запугивания выполняется до полного истощения одного из соперников, что и решает спор.
В такого рода ритуализованных столкновениях имеется специфическая последовательность движений: они начинаются с демонстрации боковых поверхностей тела, за которой следует подъем вертикальных плавников. Затем следуют удары хвостом, которые через посредство боковой линии, воспринимающей изменение давления воды, могут, вероятно, сообщить о силе противника. После этого противники встают друг перед другом, вслед за чем начинаются взаимные толчки с открытой пастью, а у других видов — укусы в открытый рот. Они продолжаются до тех пор, пока один из соперников не устанет, окраска его бледнеет и в конце концов он уплывает.
Такие ритуализованные драки и агрессивные столкновения — прекрасные примеры специфической последовательности стереотипных двигательных реакций: удары хвостом не начнутся до подъема спинного плавника, а толчки отмечаются только после многих ударов хвостом. По интенсивности демонстрации запугивания и ударов хвоста, опытный наблюдатель может определить, кто победит, и начнутся ли толчки «с открытой пастью» вообще, или же один из соперников просто сбежит до начала «серьезной драки».
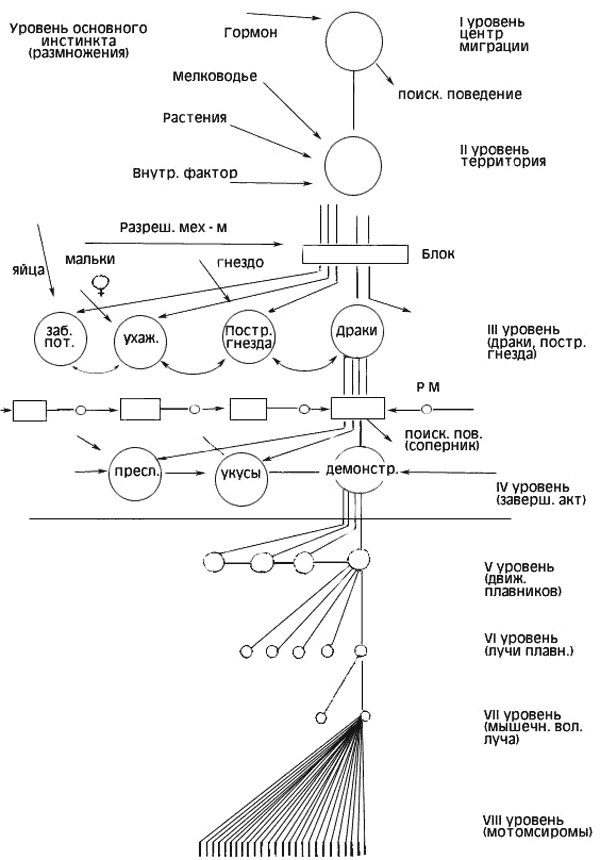
Интерпретируя подобные явления, Тинберген выдвинул гипотезу об иерархии центров, управляющих отдельными поведенческими реакциями (рис.2). Согласно Тинбергену, инстинкт представляет собой завершенную иерархическую организацию поведенческих актов, реагирующую на определенный раздражитель четко координированным комплексом действий. Нужно отметить, что понятие «центр» у Тинбергена является главным образом функциональным и не может быть полностью отождествлено с анатомическим понятием нервного центра, хотя Тинберген и ссылается на известные опыты Хесса. В этих опытах при электрораздражении промежуточного мозга кошек наблюдалось развитие некоторых целостных форм поведения (драк, угроз, сна, пищевого, поведения).
Согласно представлениям Тинбергена, изменение возбудимости центров под влиянием внешних и внутренних воздействий происходит в определенной последовательности. Сначала повышается возбудимость «центра» поисковой фазы поведения, и голодное животное начинает поиск пищи. Когда пища будет найдена, произойдет «разрядка» центра, стоящего на более низком уровне иерархии, и контролирующего осуществления завершающего акта (поедание пищи).
Схему иерархии центров, управляющих поведением самца колюшки в период размножения, Тинберген представляет следующим образом.
Высший центр репродуктивного поведения самца активизируется увеличенной длиной дня, гормональными и другими факторами. Импульсы из этого центра снимают блок с центра поискового поведения. Разрядка этого центра выражается в поисках условий для постройки гнезда. Когда такие условия (подходящая территория, температура, необходимый грунт, мелководье, растительность) найдены, происходит разрядка центров следующего уровня иерархии и благодаря этому становится возможной постройка гнезда.
Если на территорию данного самца проникает соперник, то возбудимость центра агрессивного поведения повышается. Результат этого центра агрессивного поведения — преследования и драки с самцом-соперником. Наконец, при появлении самки повышается возбудимость центра полового поведения, и начинается ухаживание за самкой, представляющее собой комплекс фиксированных действий.
В дальнейшем вопросы иерархической организации поведения изучал Хайнд (1975). Он показал, что хотя в принципе комплекс фиксированных действий большой синицы можно расположить в иерархическую схему, однако не всегда удается сделать это полностью, так как некоторые из движений характерны для двух и более видов инстинктов. Иногда эти движения являются завершающими актами, а иногда — просто средством создания условий, в которых можно осуществить завершающее действие.
У молодых животных иерархия поведения часто еще не сформирована. У птенцов, например, сначала появляются на первый взгляд бессмысленные изолированные двигательные акты, и только позже они интегрируются в сложный функциональный комплекс движений, связанных с полетом.
Расчленение иерархии поведения на элементы часто можно наблюдать во время игры, когда отдельные поведенческие акты, связанные с различными функциями, свободно комбинируются в сочетания не характерных для нормального поведения.
Существенно, что модель Тинбергена предусматривает возможность взаимодействия между «центрами» различных видов поведения. Дело в том, что случаи, когда животное в каждый данный момент занято каким-то одним видом деятельности, являются скорее исключением, чем правилом. Обычно одни виды активности сменяют другие. Наиболее простой пример такого взаимодействия — подавление одних центров другими. Например, если у самца чайки во время ухаживания за самками усиливается голод, то он может прекратить брачные демонстрации и отправиться на поиски корма. В данном случае поведение определяется не присутствием внешнего раздражителя, а соответствующим внутренним побуждением.
В такого рода ритуализованных столкновениях имеется специфическая последовательность движений: они начинаются с демонстрации боковых поверхностей тела, за которой следует подъем вертикальных плавников. Затем следуют удары хвостом, которые через посредство боковой линии, воспринимающей изменение давления воды, могут, вероятно, сообщить о силе противника. После этого противники встают друг перед другом, вслед за чем начинаются взаимные толчки с открытой пастью, а у других видов — укусы в открытый рот. Они продолжаются до тех пор, пока один из соперников не устанет, окраска его бледнеет и в конце концов он уплывает.
Такие ритуализованные драки и агрессивные столкновения — прекрасные примеры специфической последовательности стереотипных двигательных реакций: удары хвостом не начнутся до подъема спинного плавника, а толчки отмечаются только после многих ударов хвостом. По интенсивности демонстрации запугивания и ударов хвоста, опытный наблюдатель может определить, кто победит, и начнутся ли толчки «с открытой пастью» вообще, или же один из соперников просто сбежит до начала «серьезной драки».
Интерпретируя подобные явления, Тинберген выдвинул гипотезу об иерархии центров, управляющих отдельными поведенческими реакциями (рис.2). Согласно Тинбергену, инстинкт представляет собой завершенную иерархическую организацию поведенческих актов, реагирующую на определенный раздражитель четко координированным комплексом действий. Нужно отметить, что понятие «центр» у Тинбергена является главным образом функциональным и не может быть полностью отождествлено с анатомическим понятием нервного центра, хотя Тинберген и ссылается на известные опыты Хесса. В этих опытах при электрораздражении промежуточного мозга кошек наблюдалось развитие некоторых целостных форм поведения (драк, угроз, сна, пищевого, поведения).
Согласно представлениям Тинбергена, изменение возбудимости центров под влиянием внешних и внутренних воздействий происходит в определенной последовательности. Сначала повышается возбудимость «центра» поисковой фазы поведения, и голодное животное начинает поиск пищи. Когда пища будет найдена, произойдет «разрядка» центра, стоящего на более низком уровне иерархии, и контролирующего осуществления завершающего акта (поедание пищи).
Схему иерархии центров, управляющих поведением самца колюшки в период размножения, Тинберген представляет следующим образом.
Высший центр репродуктивного поведения самца активизируется увеличенной длиной дня, гормональными и другими факторами. Импульсы из этого центра снимают блок с центра поискового поведения. Разрядка этого центра выражается в поисках условий для постройки гнезда. Когда такие условия (подходящая территория, температура, необходимый грунт, мелководье, растительность) найдены, происходит разрядка центров следующего уровня иерархии и благодаря этому становится возможной постройка гнезда.
Если на территорию данного самца проникает соперник, то возбудимость центра агрессивного поведения повышается. Результат этого центра агрессивного поведения — преследования и драки с самцом-соперником. Наконец, при появлении самки повышается возбудимость центра полового поведения, и начинается ухаживание за самкой, представляющее собой комплекс фиксированных действий.
В дальнейшем вопросы иерархической организации поведения изучал Хайнд (1975). Он показал, что хотя в принципе комплекс фиксированных действий большой синицы можно расположить в иерархическую схему, однако не всегда удается сделать это полностью, так как некоторые из движений характерны для двух и более видов инстинктов. Иногда эти движения являются завершающими актами, а иногда — просто средством создания условий, в которых можно осуществить завершающее действие.
У молодых животных иерархия поведения часто еще не сформирована. У птенцов, например, сначала появляются на первый взгляд бессмысленные изолированные двигательные акты, и только позже они интегрируются в сложный функциональный комплекс движений, связанных с полетом.
Расчленение иерархии поведения на элементы часто можно наблюдать во время игры, когда отдельные поведенческие акты, связанные с различными функциями, свободно комбинируются в сочетания не характерных для нормального поведения.
Существенно, что модель Тинбергена предусматривает возможность взаимодействия между «центрами» различных видов поведения. Дело в том, что случаи, когда животное в каждый данный момент занято каким-то одним видом деятельности, являются скорее исключением, чем правилом. Обычно одни виды активности сменяют другие. Наиболее простой пример такого взаимодействия — подавление одних центров другими. Например, если у самца чайки во время ухаживания за самками усиливается голод, то он может прекратить брачные демонстрации и отправиться на поиски корма. В данном случае поведение определяется не присутствием внешнего раздражителя, а соответствующим внутренним побуждением.
Конфликтное поведение
Как особый случай проявления взаимодействия «центров» можно рассматривать так называемое конфликтное поведение, когда у животного наблюдается одновременно несколько тенденций к различным (часто противоположным) типам поведения. Одним из примеров конфликтного поведения — поведение самцов территориальных видов, описанное Тинбергеном в результате наблюдений за трехиглой колюшкой и за различными видами чаек.
Например, если самец А вторгается на территорию самца В, то последний нападает на него и преследует, а самец А спасается бегством. То же самое произойдет, если самец Б вторгнется на территорию самца А. Если же столкновение произойдет на границе этих двух территорий, то поведение обоих самцов будет выглядеть иначе: у обоих самцов элементы реакций нападения и бегства будут чередоваться. Причем элементы нападения будут выражены тем сильнее, чем ближе самец к центру своей территории. Напротив, по мере удаления от центра будут сильнее выражены элементы бегства.
Как показали наблюдения за озерной чайкой Larus redibundus, угрожающее поведение самцов на границе двух территорий включает пять поз, характер и последовательность которых зависят от реакций противника. Каждая из поз отражает определенную степень конфликта между противоположными внутренними побуждениями: агрессивностью — стремлением напасть на противника и страхом — стремлением убежать от него.
Подобный же анализ позволил объяснить и механизм так называемых «замещающих движений» (displacement activity), которые иногда также обнаруживаются у животных в конфликтных ситуациях. Например, в пограничной между двумя участками зоне два самца серебристой чайки (Larus argentatus), стоящие друг перед другом в угрожающих позах, могут внезапно начать чистить перья, белые гуси (Ansercoerulescens) на земле совершают те же движения, что и при купании, серые гуси (Anser anser) в этих ситуациях отряхиваются, а петухи клюют траву и все, что находится поблизости. Эти реакции являются, как выяснилось, врожденными, так как проявляются без соответствующего индивидуального опыта.
В других случаях конфликт страха и агрессивности приводит к тому, что животное нападает не на противника, а на более слабую особь (как это наблюдал Лоренц у серых гусей), или даже на неодушевленный предмет (чайки при этом клюют листья или землю). Такая «переадресованная» активность, как и «замещающие» действия, проявляется в тех случаях, когда агрессивность и страх уравновешиваются, уступая место другим видам активности, не связанным непосредственно с данной ситуацией.
Таким образом; иерархическая теория инстинктов Тинбергена может объяснить вышеперечисленные явления, и поведение в ситуации конфликта, и замещающие действия, и переадресованную активность.
Работы, начатые Тинбергеном и его сотрудниками, впоследствии, были продолжены и расширены. Накопленный огромный фактический материал (см. например, Хайнд, 1975) показал плодотворность такого подхода и позволил проанализировать многие виды демонстративного поведения. Результаты этих исследований частично соответствовали основным положениям схемы Тинбергена, частично требовали ее усовершенствования. Они как бы продемонстрировали пределы ее применимости и наметили направления ее дальнейшего развития.
Например, если самец А вторгается на территорию самца В, то последний нападает на него и преследует, а самец А спасается бегством. То же самое произойдет, если самец Б вторгнется на территорию самца А. Если же столкновение произойдет на границе этих двух территорий, то поведение обоих самцов будет выглядеть иначе: у обоих самцов элементы реакций нападения и бегства будут чередоваться. Причем элементы нападения будут выражены тем сильнее, чем ближе самец к центру своей территории. Напротив, по мере удаления от центра будут сильнее выражены элементы бегства.
Как показали наблюдения за озерной чайкой Larus redibundus, угрожающее поведение самцов на границе двух территорий включает пять поз, характер и последовательность которых зависят от реакций противника. Каждая из поз отражает определенную степень конфликта между противоположными внутренними побуждениями: агрессивностью — стремлением напасть на противника и страхом — стремлением убежать от него.
Подобный же анализ позволил объяснить и механизм так называемых «замещающих движений» (displacement activity), которые иногда также обнаруживаются у животных в конфликтных ситуациях. Например, в пограничной между двумя участками зоне два самца серебристой чайки (Larus argentatus), стоящие друг перед другом в угрожающих позах, могут внезапно начать чистить перья, белые гуси (Ansercoerulescens) на земле совершают те же движения, что и при купании, серые гуси (Anser anser) в этих ситуациях отряхиваются, а петухи клюют траву и все, что находится поблизости. Эти реакции являются, как выяснилось, врожденными, так как проявляются без соответствующего индивидуального опыта.
Рис. 2. Схема организации инстинкта по Тинбергену
В других случаях конфликт страха и агрессивности приводит к тому, что животное нападает не на противника, а на более слабую особь (как это наблюдал Лоренц у серых гусей), или даже на неодушевленный предмет (чайки при этом клюют листья или землю). Такая «переадресованная» активность, как и «замещающие» действия, проявляется в тех случаях, когда агрессивность и страх уравновешиваются, уступая место другим видам активности, не связанным непосредственно с данной ситуацией.
Таким образом; иерархическая теория инстинктов Тинбергена может объяснить вышеперечисленные явления, и поведение в ситуации конфликта, и замещающие действия, и переадресованную активность.
Работы, начатые Тинбергеном и его сотрудниками, впоследствии, были продолжены и расширены. Накопленный огромный фактический материал (см. например, Хайнд, 1975) показал плодотворность такого подхода и позволил проанализировать многие виды демонстративного поведения. Результаты этих исследований частично соответствовали основным положениям схемы Тинбергена, частично требовали ее усовершенствования. Они как бы продемонстрировали пределы ее применимости и наметили направления ее дальнейшего развития.
Разработка проблем эволюции поведения
Изучение эволюционных аспектов поведения составило одно из основных и наиболее важных направлений в работе Тинбергена и его учеников. Параллельно с Лоренцем они развивали выдвинутые Дарвином идеи об эволюции поведения, последовательно применяя введенный им сравнительный метод и распространяя его на более широкий круг явлений.
Основные работы этологов в этом направлении условно можно подразделить на две основные группы:
1. Выяснение таксономического значения поведенческих признаков. (Работы школы Лоренца).
2. Исследование адаптивного значения поведенческих признаков и роли естественного отбора в их формировании.
Уже согласно определению, поведение является формой приспособления организма к внешней среде. В более узком смысле под адаптивностью поведения понимают те преимущества, которые данный поведенческий признак дает виду в его борьбе за существование. Именно такой анализ и стоял в центре внимания Тинбергена и его учеников. Тинберген писал, что задача этолога состоит не в том, чтобы просто описывать те или иные формы поведения, а в том, чтобы понять, как именно они способствуют борьбе животного за существование.
Свидетельство адаптивности поведения — высокая специализация разных сторон поведенческого акта. О ней говорит прежде всего наличие видоспецифических сложнокоординированных двигательных актов. К ним относятся разнообразные ритуалы, характерные для общественного поведения, движения, связанные с поимкой добычи, строительством гнезд и т. п. реакции, относимые обычно к категории «завершающих актов».
Сенсорные механизмы, обеспечивающие срабатывание реакций типа завершающих актов, также тонко приспособлены к потребностям вида. Завершающий акт выполняется только избирательно в ответ на специфические ключевые раздражители или релизеры. Совершенствование системы сенсорного контроля происходит в двух направлениях. С одной стороны, происходит специализация органов чувств животных, связанных с восприятием ключевых стимулов. Показано, например, что у некоторых животных нейроны слухового анализатора обладают избирательной чувствительностью к специфичным для данного вида звуковым сигналам. Так, например, у пуэрториканкской древесной лягушки Eleutherodcoqulus крик самца состоит из двух слогов — «ко» и «ки». Наблюдения показали, что первый из них «адресован» самцам, являясь, видимо, видоспецифическим территориальным сигналом, второй — самкам. Слияние двух сигналов в один не препятствует существованию раздельных систем их восприятия, напоминающих о соответствии «ключа» «замку». Оказалось, что волоски базиллярной мембраны внутреннего уха самцов и самок этого вида настроены соответственно на разные тоны — либо на восприятие первого, либо на восприятие второго из слогов, составляющих крик самца. С другой стороны, в ряде случаев происходит усовершенствование самих систем, продуцирующих «ключевые раздражители», например звукоиздающего аппарата, а также дифференцированность самих сигналов.
Показано также, что во многих случаях характер онтогенетического развития поведенческого признака также оказывается адаптивным. Там, где поведенческий акт должен быть совершен уже в раннем возрасте (например, взлет бабочки или сосательные движения млекопитающих), жизненно важное значение имеет его «врожденность», и его развитие контролируется главным образом генетическими факторами. В других случаях адаптивность поведения в онтогенезе явно достигается путем обучения методом проб и ошибок. Например, многие птицы обладают врожденной способностью клевать мелкие предметы и поэтому сначала клюют, что попало, и лишь постепенно выучиваются отличать съедобные предметы от несъедобных. Такой тип организации пищевого поведения создает условия для гибкого приспособления к конкретным условиям жизни, когда вид и характер пищи могут варьировать в широких пределах.
Даже такая универсальная характеристика всего живого, как способность к обучению, имеет свою видовую специфику, обеспечивающую наилучшую приспособленность вида, и зачастую представляет собой высокоизбирательный тип взаимодействия с окружающей средой. Например, рабочие пчелы гораздо лучше других насекомых выучивают расположение цветов, на которых они кормятся. Обучение птиц пению происходит, как уже указывалось, в определенные «чувствительные» периоды онтогенеза, причем показано, что птицы предрасположены к усвоению песни своего вида (подробнее см., например, Меннинг, 1982).
В рассмотренных случаях приспособительное значение признаков поведения выступает со всей очевидностью, однако проследить путь их эволюционного развития трудно, или даже невозможно. Особенность работ Тинбергена и его учеников состоит в том, что они, с одной стороны, внесли большой вклад в исследование функционального и приспособительного значения ряда поведенческих признаков, с другой — выделили признаки, на примере которых оказалось возможным показать экспериментально формирующее и стабилизирующее влияние естественного отбора на поведение.
Успехи в выяснения адаптивного значения поведенческих признаков, достигнутые школой Тинбергена, в значительной степени связаны не только с тем, что он четко сформулировал задачу исследований, но и разработал экспериментальные подходы к ее решению. Основным материалом для наблюдений и последующих обобщений послужило поведение нескольких видов морских птиц — чаек, близких в систематическом отношении (одно семейство), но различающихся по ряду особенностей среды обитания, образа жизни, питания и т. п. Основное внимание в этих исследованиях было обращено на изучение поведенческих адаптации, связанных с защитой от хищников, необходимость которой, как выяснилось, накладывает отпечаток практически на все стороны поведения вида.
Необходимый этап в экспериментальном исследовании приспособительного значения акта поведения — его точное описание и выяснение функции, которую он выполняет.
Основные работы этологов в этом направлении условно можно подразделить на две основные группы:
1. Выяснение таксономического значения поведенческих признаков. (Работы школы Лоренца).
2. Исследование адаптивного значения поведенческих признаков и роли естественного отбора в их формировании.
Уже согласно определению, поведение является формой приспособления организма к внешней среде. В более узком смысле под адаптивностью поведения понимают те преимущества, которые данный поведенческий признак дает виду в его борьбе за существование. Именно такой анализ и стоял в центре внимания Тинбергена и его учеников. Тинберген писал, что задача этолога состоит не в том, чтобы просто описывать те или иные формы поведения, а в том, чтобы понять, как именно они способствуют борьбе животного за существование.
Свидетельство адаптивности поведения — высокая специализация разных сторон поведенческого акта. О ней говорит прежде всего наличие видоспецифических сложнокоординированных двигательных актов. К ним относятся разнообразные ритуалы, характерные для общественного поведения, движения, связанные с поимкой добычи, строительством гнезд и т. п. реакции, относимые обычно к категории «завершающих актов».
Сенсорные механизмы, обеспечивающие срабатывание реакций типа завершающих актов, также тонко приспособлены к потребностям вида. Завершающий акт выполняется только избирательно в ответ на специфические ключевые раздражители или релизеры. Совершенствование системы сенсорного контроля происходит в двух направлениях. С одной стороны, происходит специализация органов чувств животных, связанных с восприятием ключевых стимулов. Показано, например, что у некоторых животных нейроны слухового анализатора обладают избирательной чувствительностью к специфичным для данного вида звуковым сигналам. Так, например, у пуэрториканкской древесной лягушки Eleutherodcoqulus крик самца состоит из двух слогов — «ко» и «ки». Наблюдения показали, что первый из них «адресован» самцам, являясь, видимо, видоспецифическим территориальным сигналом, второй — самкам. Слияние двух сигналов в один не препятствует существованию раздельных систем их восприятия, напоминающих о соответствии «ключа» «замку». Оказалось, что волоски базиллярной мембраны внутреннего уха самцов и самок этого вида настроены соответственно на разные тоны — либо на восприятие первого, либо на восприятие второго из слогов, составляющих крик самца. С другой стороны, в ряде случаев происходит усовершенствование самих систем, продуцирующих «ключевые раздражители», например звукоиздающего аппарата, а также дифференцированность самих сигналов.
Показано также, что во многих случаях характер онтогенетического развития поведенческого признака также оказывается адаптивным. Там, где поведенческий акт должен быть совершен уже в раннем возрасте (например, взлет бабочки или сосательные движения млекопитающих), жизненно важное значение имеет его «врожденность», и его развитие контролируется главным образом генетическими факторами. В других случаях адаптивность поведения в онтогенезе явно достигается путем обучения методом проб и ошибок. Например, многие птицы обладают врожденной способностью клевать мелкие предметы и поэтому сначала клюют, что попало, и лишь постепенно выучиваются отличать съедобные предметы от несъедобных. Такой тип организации пищевого поведения создает условия для гибкого приспособления к конкретным условиям жизни, когда вид и характер пищи могут варьировать в широких пределах.
Даже такая универсальная характеристика всего живого, как способность к обучению, имеет свою видовую специфику, обеспечивающую наилучшую приспособленность вида, и зачастую представляет собой высокоизбирательный тип взаимодействия с окружающей средой. Например, рабочие пчелы гораздо лучше других насекомых выучивают расположение цветов, на которых они кормятся. Обучение птиц пению происходит, как уже указывалось, в определенные «чувствительные» периоды онтогенеза, причем показано, что птицы предрасположены к усвоению песни своего вида (подробнее см., например, Меннинг, 1982).
В рассмотренных случаях приспособительное значение признаков поведения выступает со всей очевидностью, однако проследить путь их эволюционного развития трудно, или даже невозможно. Особенность работ Тинбергена и его учеников состоит в том, что они, с одной стороны, внесли большой вклад в исследование функционального и приспособительного значения ряда поведенческих признаков, с другой — выделили признаки, на примере которых оказалось возможным показать экспериментально формирующее и стабилизирующее влияние естественного отбора на поведение.
Успехи в выяснения адаптивного значения поведенческих признаков, достигнутые школой Тинбергена, в значительной степени связаны не только с тем, что он четко сформулировал задачу исследований, но и разработал экспериментальные подходы к ее решению. Основным материалом для наблюдений и последующих обобщений послужило поведение нескольких видов морских птиц — чаек, близких в систематическом отношении (одно семейство), но различающихся по ряду особенностей среды обитания, образа жизни, питания и т. п. Основное внимание в этих исследованиях было обращено на изучение поведенческих адаптации, связанных с защитой от хищников, необходимость которой, как выяснилось, накладывает отпечаток практически на все стороны поведения вида.
Необходимый этап в экспериментальном исследовании приспособительного значения акта поведения — его точное описание и выяснение функции, которую он выполняет.
Функциональный анализ признаков поведения
Можно привести много примеров, когда функция поведенческого акта очевидна и не требует специальных исследований. Однако во многих случаях, требуются кропотливые исследования, чтобы выяснить например, функцию отдельных компонентов сложного поведенческого акта. Так, поведение птиц при насиживании имеет вполне очевидную общую функцию. Между тем, для определения конкретной функции отдельных сторон этого поведения, закономерностей их течения во времени и лежащих в их основе регуляторных механизмов потребовались годы интенсивных исследований на птицах разных видов.
В ряде случаев, несмотря на достаточную изученность поведения остаются реакции, функцию которых выявить не удается. Неясно например, почему водные насекомые, прежде чем взлететь, тщательно вытирают поверхность глаз. Не менее загадочной остается описанная у некоторых видов птиц привычка принимать «муравьиные ванны», когда птицы не просто купаются в муравейнике, как в пыли, но могут специально засовывать муравьев себе под перья.
В ряде случаев, несмотря на достаточную изученность поведения остаются реакции, функцию которых выявить не удается. Неясно например, почему водные насекомые, прежде чем взлететь, тщательно вытирают поверхность глаз. Не менее загадочной остается описанная у некоторых видов птиц привычка принимать «муравьиные ванны», когда птицы не просто купаются в муравейнике, как в пыли, но могут специально засовывать муравьев себе под перья.