Страница:
От магнитного момента зависят многие др. свойства ферромагнетика, например электрическое сопротивление, механическая деформация. Изменение магнитного момента вызывает изменение и этих свойств. Соответственно наблюдается, например, гальваномагнитный Г., магнитострикционный Г.
Диэлектрический Г. наблюдается обычно в сегнетоэлектриках , например титанате бария. Зависимость поляризации Рот напряжённости электрического поля Ев сегнетоэлектриках ( рис. 3 ) подобна зависимости Мот Нв ферромагнетиках и объясняется наличием спонтанной электрической поляризации, электрических доменов и трудностью перестройки доменной структуры. Гистерезисные потери составляют большую часть диэлектрических потерь в сегнетоэлектриках.
Поскольку с поляризацией связаны др. характеристики сегнетоэлектриков, например деформация, то с диэлектрическим Г. связаны др. виды Г., например пьезоэлектрический Г. ( рис. 4 ), Г. электрооптического эффекта . В некоторых случаях наблюдаются двойные петли диэлектрического Г. ( рис. 5 ). Это объясняется тем, что под влиянием электрического поля в образце происходит фазовый переход с перестройкой кристаллической структуры. Такого рода диэлектрический Г. тесно связан с Г. при фазовых переходах.
Упругий Г., т. е. гистерезисная зависимость деформации иот механического напряжения s, наблюдается в любых реальных материалах при достаточно больших напряжениях ( рис. 6 ). Упругий Г. возникает всякий раз, когда имеет место пластическая (неупругая) деформация (см. Пластичность ). Пластическая деформация обусловлена перемещением дефектов, например дислокаций , всегда присутствующих в реальных материалах. Примеси, включения и др. дефекты, а также сама кристаллическая решётка стремятся удержать дислокацию в определенных положениях в кристалле. Поэтому требуются напряжения достаточной величины, чтобы сдвинуть дислокацию. Механическая обработка и введение примесей приводят к закреплению дислокаций, в результате чего происходит упрочнение материала, пластическая деформация и упругий Г. наблюдаются при больших напряжениях. Энергия, теряемая в образце за один цикл, идёт в конечном счёте на нагревание образца. Потери на упругий Г. дают вклад во внутреннее трение . В случае упругих деформаций, помимо гистерезисных, есть и др. потери, например обусловленные вязкостью . Величина этих потерь, в отличие от гистерезисных, зависит от частоты изменения s(или и). Иногда понятие «упругий Г.» употребляется шире — говорят о динамической петле упругого Г., включающей все потери на данной частоте.
Лит.:Киренский Л. В., Магнетизм, 2 изд., М., 1967; Вонсовский С. В., Современное учение о магнетизме, М. — Л., 1952; Бозорт Р., Ферромагнетизм, пер. с англ., М., 1956; Иона Ф., Ширане Д., Сегнетоэлектрические кристаллы, пер. с англ., М., 1965; Постников В. С., Внутреннее трение в металлах, М., 1969; Физический энциклопедический словарь, т. 1, М., 1960.
А. П. Леванюк, Д. Г. Санников.
 Рис. 2. Влияние механической и термической обработки на форму петли магнитного гистерезиса пермалоя: 1 — после наклёпа; 2 — после отжига; 3 — кривая мягкого железа (для сравнения).
Рис. 2. Влияние механической и термической обработки на форму петли магнитного гистерезиса пермалоя: 1 — после наклёпа; 2 — после отжига; 3 — кривая мягкого железа (для сравнения).
 Рис. 5. Двойная петля диэлектрического гистерезиса.
Рис. 5. Двойная петля диэлектрического гистерезиса.
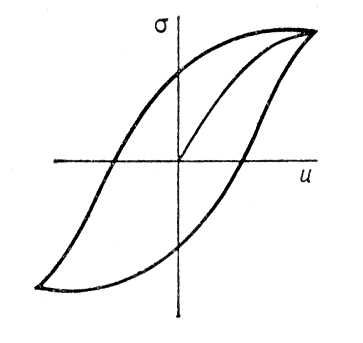 Рис. 6. Петля упругого гистерезиса: s — механическое напряжение; u — деформация.
Рис. 6. Петля упругого гистерезиса: s — механическое напряжение; u — деформация.
 Рис. 1. Петля магнитного гистерезиса для ферромагнетика: Н — напряжённость магнитного поля; М — магнитный момент образца; Н
с— коэрцитивное поле; M
r— остаточный магнитный момент; M
s— магнитный момент насыщения. Пунктиром показана непредельная петля гистерезиса. Схематически приведена доме'нная структура образца для некоторых точек петли.
Рис. 1. Петля магнитного гистерезиса для ферромагнетика: Н — напряжённость магнитного поля; М — магнитный момент образца; Н
с— коэрцитивное поле; M
r— остаточный магнитный момент; M
s— магнитный момент насыщения. Пунктиром показана непредельная петля гистерезиса. Схематически приведена доме'нная структура образца для некоторых точек петли.
 Рис. 3. Петля диэлектрического гистерезиса в сегнетоэлектрике: Р — поляризация образца; Е — напряжённость электрического поля.
Рис. 3. Петля диэлектрического гистерезиса в сегнетоэлектрике: Р — поляризация образца; Е — напряжённость электрического поля.
 Рис. 4. Петля гистерезиса обратного пьезоэлектрического эффекта в титанате бария: U — деформация: Е — напряжённость электрического поля.
Рис. 4. Петля гистерезиса обратного пьезоэлектрического эффекта в титанате бария: U — деформация: Е — напряжённость электрического поля.
синхронный электродвигатель
, у которого вращающий момент возникает за счёт гистерезиса при перемагничивании массивного ротора с сердечником из магнитного материала, имеющего широкую петлю гистерезиса. При мощностях до 100
вти частоте 400
гц. Г. э. обладают несколько лучшими по сравнению с синхронными электродвигателями энергетическими характеристиками. Г. э. надёжны в эксплуатации и долговечны, они бесшумны и способны работать с различной частотой вращения. Г. э. широко применяются в электроприводе малой мощности и в системах автоматизированного управления. В автоматических приборах управления применяются реактивно-гистерезисные синхронные двигатели мощностью 10—15
мквтс частотой вращения, не превышающей несколько
об/мин, и кпд менее 1%.
Диэлектрический Г. наблюдается обычно в сегнетоэлектриках , например титанате бария. Зависимость поляризации Рот напряжённости электрического поля Ев сегнетоэлектриках ( рис. 3 ) подобна зависимости Мот Нв ферромагнетиках и объясняется наличием спонтанной электрической поляризации, электрических доменов и трудностью перестройки доменной структуры. Гистерезисные потери составляют большую часть диэлектрических потерь в сегнетоэлектриках.
Поскольку с поляризацией связаны др. характеристики сегнетоэлектриков, например деформация, то с диэлектрическим Г. связаны др. виды Г., например пьезоэлектрический Г. ( рис. 4 ), Г. электрооптического эффекта . В некоторых случаях наблюдаются двойные петли диэлектрического Г. ( рис. 5 ). Это объясняется тем, что под влиянием электрического поля в образце происходит фазовый переход с перестройкой кристаллической структуры. Такого рода диэлектрический Г. тесно связан с Г. при фазовых переходах.
Упругий Г., т. е. гистерезисная зависимость деформации иот механического напряжения s, наблюдается в любых реальных материалах при достаточно больших напряжениях ( рис. 6 ). Упругий Г. возникает всякий раз, когда имеет место пластическая (неупругая) деформация (см. Пластичность ). Пластическая деформация обусловлена перемещением дефектов, например дислокаций , всегда присутствующих в реальных материалах. Примеси, включения и др. дефекты, а также сама кристаллическая решётка стремятся удержать дислокацию в определенных положениях в кристалле. Поэтому требуются напряжения достаточной величины, чтобы сдвинуть дислокацию. Механическая обработка и введение примесей приводят к закреплению дислокаций, в результате чего происходит упрочнение материала, пластическая деформация и упругий Г. наблюдаются при больших напряжениях. Энергия, теряемая в образце за один цикл, идёт в конечном счёте на нагревание образца. Потери на упругий Г. дают вклад во внутреннее трение . В случае упругих деформаций, помимо гистерезисных, есть и др. потери, например обусловленные вязкостью . Величина этих потерь, в отличие от гистерезисных, зависит от частоты изменения s(или и). Иногда понятие «упругий Г.» употребляется шире — говорят о динамической петле упругого Г., включающей все потери на данной частоте.
Лит.:Киренский Л. В., Магнетизм, 2 изд., М., 1967; Вонсовский С. В., Современное учение о магнетизме, М. — Л., 1952; Бозорт Р., Ферромагнетизм, пер. с англ., М., 1956; Иона Ф., Ширане Д., Сегнетоэлектрические кристаллы, пер. с англ., М., 1965; Постников В. С., Внутреннее трение в металлах, М., 1969; Физический энциклопедический словарь, т. 1, М., 1960.
А. П. Леванюк, Д. Г. Санников.

