Страница:
Вне пищеварения, натощак, содержимое двенадцатиперстной кишки имеет слабощелочную реакцию (pH 7,2–8,0). При переходе в нее порций кислого содержимого из желудка реакция дуоденального содержимого также становится кислой, но затем быстро происходит ее изменение, так как соляная кислота желудочного сока здесь нейтрализуется желчью, соком поджелудочной железы, а также дуоденальных (бруннеровых) желез и кишечных крипт (либеркюновы железы). При этом действие желудочного пепсина прекращается. Чем выше кислотность дуоденального содержимого, тем больше выделяется сока поджелудочной железы и желчи и тем больше замедляется эвакуация содержимого желудка в двенадцатиперстную кишку. В гидролизе питательных веществ в двенадцатиперстной кишке особенно велика роль ферментов сока поджелудочной железы, желчи.
Пищеварение в тонкой кишке – наиболее важный этап пищеварительного процесса в целом. Оно обеспечивает деполимеризацию питательных веществ до стадии мономеров, которые всасываются из кишечника в кровь и лимфу. Пищеварение в тонкой кишке происходит сначала в ее полости (полостное пищеварение), а затем в зоне щеточной каймы кишечного эпителия при помощи ферментов, встроенных в мембрану микроворсинок кишечных клеток, а также фиксированных в гликокаликсе (мембранное пищеварение). Полостное и мембранное пищеварение осуществляется ферментами, поступающими с соком поджелудочной железы, а также собственно кишечными ферментами (мембранными, или трансмембранными) (см. табл. 2.1). Важную роль в расщеплении липидов играет желчь.
Для человека наиболее характерна комбинация полостного и мембранного пищеварения. Начальные этапы гидролиза осуществляются за счет полостного пищеварения. Большинство надмолекулярных комплексов и крупных молекул (белки и продукты их неполного гидролиза, углеводы, жиры) расщепляются в полости тонкой кишки в нейтральной и слабощелочной средах, главным образом под действием эндогидролаз, секретируемых клетками поджелудочной железы. Часть этих ферментов может адсорбироваться на структурах слизи или слизистых наложениях. Пептиды, образующиеся в проксимальном отделе кишки и состоящие из 2–6 аминокислотных остатков, дают 60–70 % α-аминоазота, а в дистальной части кишки – до 50 %.
Углеводы (полисахариды, крахмал, гликоген) расщепляются α-амилазой поджелудочного сока до декстринов, три– и дисахаридов без значительного накопления глюкозы. Жиры подвергаются гидролизу в полости тонкой кишки панкреатической липазой, которая поэтапно отщепляет жирные кислоты, что приводит к образованию ди– и моноглицеридов, свободных жирных кислот и глицерина. В гидролизе жиров существенную роль играет желчь.
Образующиеся в полости тонкой кишки продукты частичного гидролиза, благодаря кишечной моторике, поступают из полости тонкой кишки в зону щеточной каймы, чему способствует их перенос в потоках растворителя (воды), возникающих благодаря всасыванию ионов натрия и воды. Именно на структурах щеточной каймы и происходит мембранное пищеварение. При этом промежуточные этапы гидролиза биополимеров реализуются панкреатическими ферментами, адсорбированными на структурах апикальной поверхности энтероцитов (гликокаликса), а заключительные – собственно кишечными мембранными ферментами (мальтазой, сахаразой, α-амилазой, изомальтазой, трегалазой, аминопептидазой, три– и дипептидазами, щелочной фосфатазой, моноглицеридлипазой и др.)> встроенными в мембрану энтероцитов, покрывающую микроворсинки щеточной каймы. Некоторые ферменты (α-амилаза и аминопептидаза) гидролизуют и высокополимеризованные продукты.
Пептиды, поступающие в область щеточной каймы кишечных клеток, расщепляются до олигопептидов, дипептидов и аминокислот, способных к всасыванию. Пептиды, состоящие более чем из трех аминокислотных остатков, гидролизуются преимущественно ферментами щеточной каймы, а три– и дипептиды – как ферментами щеточной каймы, так и внутриклеточно ферментами цитоплазмы. Глицилглицин и некоторые дипептиды, содержащие остатки пролина и оксипролина и не имеющие существенного нутритивного значения, всасываются частично или полностью в нерасщепленном виде. Дисахариды, поступающие с пищей (например, сахароза), а также образующиеся при расщеплении крахмала и гликогена, гидролизуются собственно кишечными гликозидазами до моносахаридов, которые транспортируются через кишечный барьер во внутреннюю среду организма. Триглицериды расщепляются не только под действием панкреатической липазы, но и под влиянием кишечной моноглицеридлипазы.
Секреция
Всасывание
Моторика
Защитные системы
Пищеварение в тонкой кишке – наиболее важный этап пищеварительного процесса в целом. Оно обеспечивает деполимеризацию питательных веществ до стадии мономеров, которые всасываются из кишечника в кровь и лимфу. Пищеварение в тонкой кишке происходит сначала в ее полости (полостное пищеварение), а затем в зоне щеточной каймы кишечного эпителия при помощи ферментов, встроенных в мембрану микроворсинок кишечных клеток, а также фиксированных в гликокаликсе (мембранное пищеварение). Полостное и мембранное пищеварение осуществляется ферментами, поступающими с соком поджелудочной железы, а также собственно кишечными ферментами (мембранными, или трансмембранными) (см. табл. 2.1). Важную роль в расщеплении липидов играет желчь.
Для человека наиболее характерна комбинация полостного и мембранного пищеварения. Начальные этапы гидролиза осуществляются за счет полостного пищеварения. Большинство надмолекулярных комплексов и крупных молекул (белки и продукты их неполного гидролиза, углеводы, жиры) расщепляются в полости тонкой кишки в нейтральной и слабощелочной средах, главным образом под действием эндогидролаз, секретируемых клетками поджелудочной железы. Часть этих ферментов может адсорбироваться на структурах слизи или слизистых наложениях. Пептиды, образующиеся в проксимальном отделе кишки и состоящие из 2–6 аминокислотных остатков, дают 60–70 % α-аминоазота, а в дистальной части кишки – до 50 %.
Углеводы (полисахариды, крахмал, гликоген) расщепляются α-амилазой поджелудочного сока до декстринов, три– и дисахаридов без значительного накопления глюкозы. Жиры подвергаются гидролизу в полости тонкой кишки панкреатической липазой, которая поэтапно отщепляет жирные кислоты, что приводит к образованию ди– и моноглицеридов, свободных жирных кислот и глицерина. В гидролизе жиров существенную роль играет желчь.
Образующиеся в полости тонкой кишки продукты частичного гидролиза, благодаря кишечной моторике, поступают из полости тонкой кишки в зону щеточной каймы, чему способствует их перенос в потоках растворителя (воды), возникающих благодаря всасыванию ионов натрия и воды. Именно на структурах щеточной каймы и происходит мембранное пищеварение. При этом промежуточные этапы гидролиза биополимеров реализуются панкреатическими ферментами, адсорбированными на структурах апикальной поверхности энтероцитов (гликокаликса), а заключительные – собственно кишечными мембранными ферментами (мальтазой, сахаразой, α-амилазой, изомальтазой, трегалазой, аминопептидазой, три– и дипептидазами, щелочной фосфатазой, моноглицеридлипазой и др.)> встроенными в мембрану энтероцитов, покрывающую микроворсинки щеточной каймы. Некоторые ферменты (α-амилаза и аминопептидаза) гидролизуют и высокополимеризованные продукты.
Пептиды, поступающие в область щеточной каймы кишечных клеток, расщепляются до олигопептидов, дипептидов и аминокислот, способных к всасыванию. Пептиды, состоящие более чем из трех аминокислотных остатков, гидролизуются преимущественно ферментами щеточной каймы, а три– и дипептиды – как ферментами щеточной каймы, так и внутриклеточно ферментами цитоплазмы. Глицилглицин и некоторые дипептиды, содержащие остатки пролина и оксипролина и не имеющие существенного нутритивного значения, всасываются частично или полностью в нерасщепленном виде. Дисахариды, поступающие с пищей (например, сахароза), а также образующиеся при расщеплении крахмала и гликогена, гидролизуются собственно кишечными гликозидазами до моносахаридов, которые транспортируются через кишечный барьер во внутреннюю среду организма. Триглицериды расщепляются не только под действием панкреатической липазы, но и под влиянием кишечной моноглицеридлипазы.
Секреция
В слизистой оболочке тонкой кишки имеются железистые клетки, расположенные на ворсинках, которые продуцируют пищеварительные секреты, выделяющиеся в кишку. Это бруннеровы железы двенадцатиперстной кишки, либеркюновы крипты тощей кишки, бокаловидные клетки. Эндокринными клетками вырабатываются гормоны, которые поступают в межклеточное пространство, а откуда транспортируются в лимфу и кровь. Здесь же локализованы выделяющие белковый секрет клетки с ацидофильными гранулами в цитоплазме (клетки Панета). Объем кишечного сока (в норме до 2,5 л) может возрастать при местном воздействии некоторых пищевых или токсических субстанций на слизистую оболочку кишечника. Прогрессирующая дистрофия и атрофия слизистой оболочки тонкой кишки сопровождаются уменьшением секреции кишечного сока.
Железистые клетки образуют и накапливают секрет и на определенной стадии своей деятельности отторгаются в просвет кишки, где, распадаясь, отдают этот секрет в окружающую жидкость. Сок можно разделить на жидкую и плотную части, соотношение между которыми меняется в зависимости от силы и характера раздражения кишечных клеток. В жидкой части сока содержится около 20 г/л сухого вещества, состоящего частично из содержимого десквамированных клеток, поступающих из крови органических (слизь, белки, мочевина и др.) и неорганических веществ – примерно 10 г/л (таких как бикарбонаты, хлориды, фосфаты). Плотная часть кишечного сока имеет вид слизистых комков и состоит из неразрушенных десквамированных эпителиальных клеток, их фрагментов и слизи (секрета бокаловидных клеток).
У здоровых людей периодическая секреция характеризуется относительной качественной и количественной стабильностью, способствующей поддержанию гомеостаза энтеральной среды, какой является в первую очередь химус.
По некоторым расчетам у взрослого человека с пищеварительными соками поступает в пищу до 140 г белка в сутки, еще 25 г белковых субстратов образуется в результате десквамации кишечного эпителия. Не трудно представить значительность белковых потерь, которые могут происходить при длительной и тяжелой диарее, при любых формах нарушения пищеварения, патологических состояниях, связанных с энтеральной недостаточностью – усилением тонкокишечной секреции и нарушением обратного всасывания (реабсорбции).
Слизь, синтезируемая бокаловидными клетками тонкой кишки, является важным компонентом секреторной активности. Количество бокаловидных клеток в составе ворсинок больше, чем в криптах (приблизительно до 70 %), и увеличивается в дистальных отделах тонкой кишки. По-видимому, это отражает важность непищеварительных функций слизи. Установлено, что клеточный эпителий тонкой кишки покрыт сплошным гетерогенным слоем толщиной до 50-кратной высоты энтероцита. В этом надэпителиальном слое слизистых наложений содержится значительное количество адсорбированных панкреатических и незначительное количество кишечных ферментов, реализующих пищеварительную функцию слизи. Слизистый секрет богат кислыми и нейтральными мукополисахаридами, но беден белками. Это обеспечивает цитопротективную состоятельность слизистого геля, механическую, химическую защиту слизистой оболочки, предотвращение проникновения в глубинные структуры ткани крупномолекулярных соединений и антигенных агрессоров.
Железистые клетки образуют и накапливают секрет и на определенной стадии своей деятельности отторгаются в просвет кишки, где, распадаясь, отдают этот секрет в окружающую жидкость. Сок можно разделить на жидкую и плотную части, соотношение между которыми меняется в зависимости от силы и характера раздражения кишечных клеток. В жидкой части сока содержится около 20 г/л сухого вещества, состоящего частично из содержимого десквамированных клеток, поступающих из крови органических (слизь, белки, мочевина и др.) и неорганических веществ – примерно 10 г/л (таких как бикарбонаты, хлориды, фосфаты). Плотная часть кишечного сока имеет вид слизистых комков и состоит из неразрушенных десквамированных эпителиальных клеток, их фрагментов и слизи (секрета бокаловидных клеток).
У здоровых людей периодическая секреция характеризуется относительной качественной и количественной стабильностью, способствующей поддержанию гомеостаза энтеральной среды, какой является в первую очередь химус.
По некоторым расчетам у взрослого человека с пищеварительными соками поступает в пищу до 140 г белка в сутки, еще 25 г белковых субстратов образуется в результате десквамации кишечного эпителия. Не трудно представить значительность белковых потерь, которые могут происходить при длительной и тяжелой диарее, при любых формах нарушения пищеварения, патологических состояниях, связанных с энтеральной недостаточностью – усилением тонкокишечной секреции и нарушением обратного всасывания (реабсорбции).
Слизь, синтезируемая бокаловидными клетками тонкой кишки, является важным компонентом секреторной активности. Количество бокаловидных клеток в составе ворсинок больше, чем в криптах (приблизительно до 70 %), и увеличивается в дистальных отделах тонкой кишки. По-видимому, это отражает важность непищеварительных функций слизи. Установлено, что клеточный эпителий тонкой кишки покрыт сплошным гетерогенным слоем толщиной до 50-кратной высоты энтероцита. В этом надэпителиальном слое слизистых наложений содержится значительное количество адсорбированных панкреатических и незначительное количество кишечных ферментов, реализующих пищеварительную функцию слизи. Слизистый секрет богат кислыми и нейтральными мукополисахаридами, но беден белками. Это обеспечивает цитопротективную состоятельность слизистого геля, механическую, химическую защиту слизистой оболочки, предотвращение проникновения в глубинные структуры ткани крупномолекулярных соединений и антигенных агрессоров.
Всасывание
Под всасыванием понимается совокупность процессов, в результате которых компоненты пищи, содержащиеся в пищеварительных полостях, переносятся через клеточные слои и межклеточные пути во внутренние циркуляторные среды организма – кровь и лимфу. Главным органом всасывания служит тонкая кишка, хотя некоторые пищевые компоненты могут всасываться в толстой кишке, желудке и даже ротовой полости. Пищевые вещества, поступающие из тонкой кишки, с током крови и лимфы разносятся по всему организму и далее участвуют в интермедиарном (промежуточном) обмене. В сутки в желудочно-кишечном тракте всасывается до 8–9 л жидкости. Из них приблизительно 2,5 л поступает с пищей и питьем, остальное – жидкость секретов пищеварительного аппарата.
Всасывание большинства пищевых веществ происходит после их ферментативной обработки и деполимеризации, которые происходят как в полости тонкой кишки, так и на ее поверхности за счет мембранного пищеварения. Уже через 3–7 ч после приема пищи все ее основные компоненты исчезают из полости тонкой кишки. Интенсивность всасывания пищевых веществ в различных отделах тонкой кишки неодинакова и зависит от топографии соответствующих ферментативных и транспортных активностей вдоль кишечной трубки (рис. 2.4).
Различают два типа транспорта через кишечный барьер во внутреннюю среду организма. Это – трансмембранный (трансцеллюлярный, через клетку) и парацеллюлярный (шунтирующий, идущий через межклеточные пространства).
Основным типом транспорта является трансмембранный. Условно можно выделить два вида трансмембранного переноса веществ через биологические мембраны – это макромолекулярный и микромолекулярный. Под макромолекулярным транспортом понимается перенос крупных молекул и молекулярных агрегатов через клеточные слои. Этот транспорт прерывист и реализуется преимущественно посредством пино– и фагоцитоза, объединяемых названием «эндоцитоз». За счет этого механизма в организм могут поступать белки, в том числе антитела, аллергены и некоторые другие соединения, значимые для организма.
Микромолекулярный транспорт служит основным типом, в результате которого из кишечной среды во внутреннюю среду организма переносятся продукты гидролиза пищевых веществ, преимущественно мономеры, различные ионы, лекарственные препараты и другие соединения, обладающие небольшой молекулярной массой. Транспорт углеводов через плазматическую мембрану кишечных клеток происходит в виде моносахаридов (глюкозы, галактозы, фруктозы и т. д.), белков – преимущественно в виде аминокислот, жиров – в виде глицерина и жирных кислот.
Во время трансмембранного движения вещество пересекает мембрану микроворсинок щеточной каймы кишечных клеток, поступает в цитоплазму, затем через базолатеральную мембрану – в лимфатические и кровеносные сосуды кишечных ворсинок и далее в общую систему циркуляции. Цитоплазма кишечных клеток служит компартментом, образующим градиент между щеточной каймой и базолатеральной мембраной.
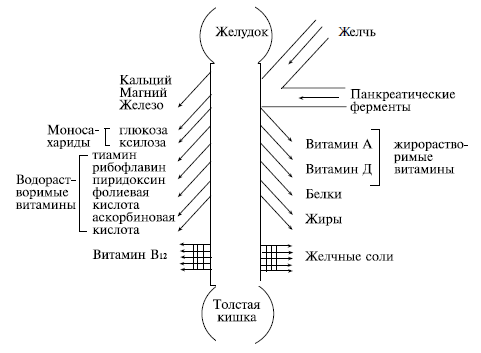 Рис. 2.4. Распределение резорбтивных функций вдоль тонкой кишки (по: С. D. Booth, 1967, с изменениями).
Рис. 2.4. Распределение резорбтивных функций вдоль тонкой кишки (по: С. D. Booth, 1967, с изменениями).
В микромолекулярном транспорте в свою очередь принято выделять пассивный и активный транспорт. Пассивный транспорт может происходить благодаря диффузии веществ через мембрану или водные поры по концентрационному градиенту, осмотическому или гидростатическому давлению. Он ускоряется благодаря движущимся через поры потокам воды, изменениям градиента pH, а также транспортерам в мембране (в случае облегченной диффузии их работа осуществляется без затраты энергии). Обменная диффузия обеспечивает микроциркуляцию ионов между периферией клетки и окружающей ее микросредой. Облегченная диффузия реализуется с помощью особых транспортеров – специальных белковых молекул (специфических транспортных белков), способствующих без затраты энергии проникновению субстанций через мембрану клеток за счет концентрационного градиента.
Активно транспортируемое вещество перемещается через апикальную мембрану кишечной клетки против своего электромеханического градиента с участием специальных транспортных систем, функционирующих по типу мобильных или конформационных транспортеров (переносчиков) с затратой энергии. Этим активный транспорт резко отличается от облегченной диффузии.
Транспорт большинства органических мономеров через мембрану щеточной каймы кишечных клеток зависит от ионов натрия. Это справедливо для глюкозы, галактозы, лактата, большинства аминокислот, некоторых конъюгированных желчных кислот, а также для ряда других соединений. Движущей силой такого транспорта служит градиент концентрации Na+. Однако в клетках тонкой кишки существует не только Ма+-зависимая транспортная система, но и Ма+-независимая, которая свойственна некоторым аминокислотам.
Вода всасывается из кишечника в кровь и поступает обратно по законам осмоса, но большая часть – из изотонических растворов кишечного химуса, так как в кишечнике гипер– и гипотонические растворы быстро разводятся или концентрируются.
Всасывание ионов натрия в кишечнике происходит как через базолатеральную мембрану в межклеточное пространство и далее в кровь, так и трансцеллюлярным путем. За сутки в пищеварительный тракт человека поступает с пищей 5–8 г натрия, 20–30 г этого иона секретируется с пищеварительными соками (т. е. всего 25–35 г). Часть ионов натрия всасывается вместе с ионами хлора, а также во время противоположно направленного транспорта ионов калия за счет Na+, К+-АТФазы.
Всасывание двухвалентных ионов (Са2+, Mg2+, Zn2+, Fe2+) происходит по всей длине желудочно-кишечного тракта, а Си2+ – главным образом в желудке. Двухвалентные ионы всасываются очень медленно. Всасывание Са2+ наиболее активно происходит в двенадцатиперстной и тощей кишках с участием механизмов простой и облегченной диффузии, активируется витамином D, соком поджелудочной железы, желчью и рядом других соединений.
Углеводы всасываются в тонкой кишке в виде моносахаридов (глюкозы, фруктозы, галактозы). Всасывание глюкозы происходит активно с затратой энергии. В настоящее время уже известна молекулярная структура №+-зависимого транспортера глюкозы. Это белковый олигомер с высокой молекулярной массой и экстрацеллюлярными петлями, обладающий центрами связывания глюкозы и натрия.
Белки всасываются через апикальную мембрану кишечных клеток преимущественно в виде аминокислот и в значительно меньшей мере в виде дипептидов и трипептидов. Как и в случае с моносахаридами, энергия для транспорта аминокислот обеспечивается натриевым котранспортером.
В щеточной кайме энтероцитов существует по меньшей мере шесть Na+-зависимых транспортных систем для различных аминокислот и три – независимых от натрия. Пептидный (или аминокислотный) транспортер, как и транспортер глюкозы, представляет собой олигомерный гликозилированный белок с экстрацеллюлярной петлей.
Что касается всасывания пептидов, или так называемого пептидного транспорта, то в ранние сроки постнатального развития в тонкой кишке имеет место всасывание интактных белков. В настоящее время принято, что вообще всасывание интактных белков – процесс физиологический, необходимый для отбора антигенов субэпителиальными структурами. Однако на фоне общего поступления белков пищи преимущественно в виде аминокислот этот процесс имеет весьма малое нутритивное значение. Ряд дипептидов может поступать в цитоплазму трансмембранным путем, как и некоторые трипептиды, и расщепляться внутриклеточно.
Транспорт липидов осуществляется по-другому. Образовавшиеся при гидролизе жиров пищи длинноцепочечные жирные кислоты и глицерин практически пассивно переносятся через апикальную мембрану в энтероцит, где ресинтезируются в триглицериды и заключаются в липопротеиновую оболочку, белковый компонент которой синтезируется в энтероците. Тем самым образуется хиломикрон, который транспортируется в центральный лимфатический сосуд кишечной ворсинки и по системе грудного лимфатического протока затем поступает в кровь. Среднецепочечные и короткоцепочечные жирные кислоты поступают в кровоток сразу, без ресинтеза триглицеридов.
Скорость всасывания в тонкой кишке зависит от уровня ее кровоснабжения (влияет на процессы активного транспорта), уровня внутрикишечного давления (влияет на процессы фильтрации из просвета кишки) и топографии всасывания. Сведения об этой топографии позволяют представить себе особенности дефицита всасывания при энтеральной патологии, при пострезекционных синдромах и других нарушениях желудочно-кишечного тракта. На рис. 2.5 представлена схема контроля за процессами, происходящими в желудочно-кишечном тракте.
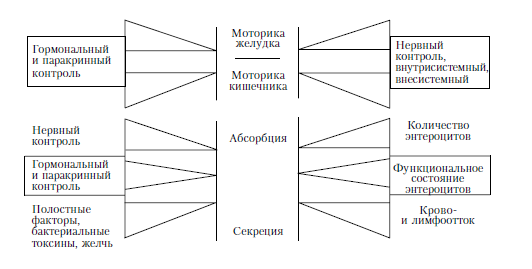
Рис. 2.5. Факторы, влияющие на процессы секреции и абсорбции в тонкой кишке (по: R. J. Levin, 1982, с изменениями).
Всасывание большинства пищевых веществ происходит после их ферментативной обработки и деполимеризации, которые происходят как в полости тонкой кишки, так и на ее поверхности за счет мембранного пищеварения. Уже через 3–7 ч после приема пищи все ее основные компоненты исчезают из полости тонкой кишки. Интенсивность всасывания пищевых веществ в различных отделах тонкой кишки неодинакова и зависит от топографии соответствующих ферментативных и транспортных активностей вдоль кишечной трубки (рис. 2.4).
Различают два типа транспорта через кишечный барьер во внутреннюю среду организма. Это – трансмембранный (трансцеллюлярный, через клетку) и парацеллюлярный (шунтирующий, идущий через межклеточные пространства).
Основным типом транспорта является трансмембранный. Условно можно выделить два вида трансмембранного переноса веществ через биологические мембраны – это макромолекулярный и микромолекулярный. Под макромолекулярным транспортом понимается перенос крупных молекул и молекулярных агрегатов через клеточные слои. Этот транспорт прерывист и реализуется преимущественно посредством пино– и фагоцитоза, объединяемых названием «эндоцитоз». За счет этого механизма в организм могут поступать белки, в том числе антитела, аллергены и некоторые другие соединения, значимые для организма.
Микромолекулярный транспорт служит основным типом, в результате которого из кишечной среды во внутреннюю среду организма переносятся продукты гидролиза пищевых веществ, преимущественно мономеры, различные ионы, лекарственные препараты и другие соединения, обладающие небольшой молекулярной массой. Транспорт углеводов через плазматическую мембрану кишечных клеток происходит в виде моносахаридов (глюкозы, галактозы, фруктозы и т. д.), белков – преимущественно в виде аминокислот, жиров – в виде глицерина и жирных кислот.
Во время трансмембранного движения вещество пересекает мембрану микроворсинок щеточной каймы кишечных клеток, поступает в цитоплазму, затем через базолатеральную мембрану – в лимфатические и кровеносные сосуды кишечных ворсинок и далее в общую систему циркуляции. Цитоплазма кишечных клеток служит компартментом, образующим градиент между щеточной каймой и базолатеральной мембраной.
В микромолекулярном транспорте в свою очередь принято выделять пассивный и активный транспорт. Пассивный транспорт может происходить благодаря диффузии веществ через мембрану или водные поры по концентрационному градиенту, осмотическому или гидростатическому давлению. Он ускоряется благодаря движущимся через поры потокам воды, изменениям градиента pH, а также транспортерам в мембране (в случае облегченной диффузии их работа осуществляется без затраты энергии). Обменная диффузия обеспечивает микроциркуляцию ионов между периферией клетки и окружающей ее микросредой. Облегченная диффузия реализуется с помощью особых транспортеров – специальных белковых молекул (специфических транспортных белков), способствующих без затраты энергии проникновению субстанций через мембрану клеток за счет концентрационного градиента.
Активно транспортируемое вещество перемещается через апикальную мембрану кишечной клетки против своего электромеханического градиента с участием специальных транспортных систем, функционирующих по типу мобильных или конформационных транспортеров (переносчиков) с затратой энергии. Этим активный транспорт резко отличается от облегченной диффузии.
Транспорт большинства органических мономеров через мембрану щеточной каймы кишечных клеток зависит от ионов натрия. Это справедливо для глюкозы, галактозы, лактата, большинства аминокислот, некоторых конъюгированных желчных кислот, а также для ряда других соединений. Движущей силой такого транспорта служит градиент концентрации Na+. Однако в клетках тонкой кишки существует не только Ма+-зависимая транспортная система, но и Ма+-независимая, которая свойственна некоторым аминокислотам.
Вода всасывается из кишечника в кровь и поступает обратно по законам осмоса, но большая часть – из изотонических растворов кишечного химуса, так как в кишечнике гипер– и гипотонические растворы быстро разводятся или концентрируются.
Всасывание ионов натрия в кишечнике происходит как через базолатеральную мембрану в межклеточное пространство и далее в кровь, так и трансцеллюлярным путем. За сутки в пищеварительный тракт человека поступает с пищей 5–8 г натрия, 20–30 г этого иона секретируется с пищеварительными соками (т. е. всего 25–35 г). Часть ионов натрия всасывается вместе с ионами хлора, а также во время противоположно направленного транспорта ионов калия за счет Na+, К+-АТФазы.
Всасывание двухвалентных ионов (Са2+, Mg2+, Zn2+, Fe2+) происходит по всей длине желудочно-кишечного тракта, а Си2+ – главным образом в желудке. Двухвалентные ионы всасываются очень медленно. Всасывание Са2+ наиболее активно происходит в двенадцатиперстной и тощей кишках с участием механизмов простой и облегченной диффузии, активируется витамином D, соком поджелудочной железы, желчью и рядом других соединений.
Углеводы всасываются в тонкой кишке в виде моносахаридов (глюкозы, фруктозы, галактозы). Всасывание глюкозы происходит активно с затратой энергии. В настоящее время уже известна молекулярная структура №+-зависимого транспортера глюкозы. Это белковый олигомер с высокой молекулярной массой и экстрацеллюлярными петлями, обладающий центрами связывания глюкозы и натрия.
Белки всасываются через апикальную мембрану кишечных клеток преимущественно в виде аминокислот и в значительно меньшей мере в виде дипептидов и трипептидов. Как и в случае с моносахаридами, энергия для транспорта аминокислот обеспечивается натриевым котранспортером.
В щеточной кайме энтероцитов существует по меньшей мере шесть Na+-зависимых транспортных систем для различных аминокислот и три – независимых от натрия. Пептидный (или аминокислотный) транспортер, как и транспортер глюкозы, представляет собой олигомерный гликозилированный белок с экстрацеллюлярной петлей.
Что касается всасывания пептидов, или так называемого пептидного транспорта, то в ранние сроки постнатального развития в тонкой кишке имеет место всасывание интактных белков. В настоящее время принято, что вообще всасывание интактных белков – процесс физиологический, необходимый для отбора антигенов субэпителиальными структурами. Однако на фоне общего поступления белков пищи преимущественно в виде аминокислот этот процесс имеет весьма малое нутритивное значение. Ряд дипептидов может поступать в цитоплазму трансмембранным путем, как и некоторые трипептиды, и расщепляться внутриклеточно.
Транспорт липидов осуществляется по-другому. Образовавшиеся при гидролизе жиров пищи длинноцепочечные жирные кислоты и глицерин практически пассивно переносятся через апикальную мембрану в энтероцит, где ресинтезируются в триглицериды и заключаются в липопротеиновую оболочку, белковый компонент которой синтезируется в энтероците. Тем самым образуется хиломикрон, который транспортируется в центральный лимфатический сосуд кишечной ворсинки и по системе грудного лимфатического протока затем поступает в кровь. Среднецепочечные и короткоцепочечные жирные кислоты поступают в кровоток сразу, без ресинтеза триглицеридов.
Скорость всасывания в тонкой кишке зависит от уровня ее кровоснабжения (влияет на процессы активного транспорта), уровня внутрикишечного давления (влияет на процессы фильтрации из просвета кишки) и топографии всасывания. Сведения об этой топографии позволяют представить себе особенности дефицита всасывания при энтеральной патологии, при пострезекционных синдромах и других нарушениях желудочно-кишечного тракта. На рис. 2.5 представлена схема контроля за процессами, происходящими в желудочно-кишечном тракте.
Рис. 2.5. Факторы, влияющие на процессы секреции и абсорбции в тонкой кишке (по: R. J. Levin, 1982, с изменениями).
Моторика
Существенное значение для процессов пищеварения в тонкой кишке имеет моторно-эвакуаторная деятельность, которая обеспечивает перемешивание пищевого содержимого с пищеварительными секретами, продвижение химуса по кишке, смену слоя химуса на поверхности слизистой оболочки, повышение внутрикишечного давления, способствующего фильтрации некоторых компонентов химуса из полости кишки в кровь и лимфу. Двигательная активность тонкой кишки состоит из непропульсивных перемешивающих движений и пропульсивной перистальтики. Она зависит от собственной активности гладкомышечных клеток и от влияния вегетативной нервной системы и многочисленных гормонов, в основном желудочно-кишечного происхождения.
Итак, сокращения тонкой кишки происходят в результате координированных движений продольного (наружного) и поперечного (циркуляторного) слоев волокон. Эти сокращения могут быть нескольких типов. По функциональному принципу все сокращения делят на две группы:
1) локальные, которые обеспечивают перемешивание и растирание содержимого тонкой кишки (непропульсивные);
2) направленные на передвижение содержимого кишки (пропульсивные). Выделяют несколько типов сокращений: ритмическую сегментацию, маятникообразные, перистальтические (очень медленные, медленные, быстрые, стремительные), антиперистальтические и тонические.
Ритмическая сегментация обеспечивается преимущественно сокращением циркуляторного слоя мышц. При этом содержимое кишечника разделяется на части. Следующим сокращением образуется новый сегмент кишки, содержимое которого состоит из частей бывшего сегмента. Этим достигаются перемешивание химуса и повышение давления в каждом из образующих сегментов кишки. Маятникообразные сокращения обеспечиваются сокращениями продольного слоя мышц с участием циркуляторного. При этих сокращениях происходит перемещение химуса вперед-назад и слабое поступательное движение в аборальном направлении. В проксимальных отделах тонкой кишки частота ритмических сокращений, или циклов, составляет 9-12, в дистальных – 6–8 в 1 мин.
Перистальтика состоит в том, что выше химуса за счет сокращения циркуляторного слоя мышц образуется перехват, а ниже в результате сокращения продольных мышц – расширение полости кишки. Этот перехват и расширение движутся вдоль кишки, перемещая впереди перехвата порцию химуса. По длине кишки одновременно движется несколько перистальтических волн. При антиперистальтических сокращениях волна движется в обратном (оральном) направлении. В норме тонкая кишка антиперистальтически не сокращается. Тонические сокращения могут иметь небольшую скорость, а иногда вообще не распространяться, значительно суживая просвет кишки на большом протяжении.
Выявлена определенная роль моторики в выведении пищеварительных секретов – перистальтика протоков, изменение их тонуса, закрытие и раскрытие их сфинктеров, сокращение и расслабление желчного пузыря. К этому же следует присоединить изменения складчатости слизистой оболочки, микромоторику кишечных ворсинок и микроворсинок тонкой кишки – очень важные явления, оптимизирующие мембранное пищеварение, всасывание нутриентов и других веществ из кишки в кровь и лимфу.
Моторика тонкой кишки регулируется нервными и гуморальными механизмами. Координирующее влияние оказывают интрамуральные (в стенке кишки) нервные образования, а также центральная нервная система. Интрамуральные нейроны обеспечивают координированные сокращения кишки. Особенно велика их роль в перистальтических сокращениях. На интрамуральные механизмы оказывают влияние экстрамуральные, парасимпатические и симпатические нервные механизмы, а также гуморальные факторы.
Моторная активность кишки зависит в том числе от физических и химических свойств химуса. Повышает ее активность грубая пища (черный хлеб, овощи, грубоволокнистые продукты) и жиры. При средней скорости перемещения 1–4 см/мин пища достигает слепой кишки за 2–4 ч. На продолжительность перемещения пищи влияет ее состав, в зависимости от него скорость перемещения уменьшается в ряду: углеводы, белки, жиры.
Гуморальные вещества изменяют моторику кишечника, действуя непосредственно на мышечные волокна и через рецепторы на нейроны интрамуральной нервной системы. Усиливают моторику тонкой кишки вазопрессин, окситоцин, брадикинин, серотонин, гистамин, гастрин, мотилин, холецистокинин-панкреозимин, субстанция Р и ряд других веществ (кислоты, щелочи, соли, продукты переваривания пищевых веществ, особенно жиров).
Итак, сокращения тонкой кишки происходят в результате координированных движений продольного (наружного) и поперечного (циркуляторного) слоев волокон. Эти сокращения могут быть нескольких типов. По функциональному принципу все сокращения делят на две группы:
1) локальные, которые обеспечивают перемешивание и растирание содержимого тонкой кишки (непропульсивные);
2) направленные на передвижение содержимого кишки (пропульсивные). Выделяют несколько типов сокращений: ритмическую сегментацию, маятникообразные, перистальтические (очень медленные, медленные, быстрые, стремительные), антиперистальтические и тонические.
Ритмическая сегментация обеспечивается преимущественно сокращением циркуляторного слоя мышц. При этом содержимое кишечника разделяется на части. Следующим сокращением образуется новый сегмент кишки, содержимое которого состоит из частей бывшего сегмента. Этим достигаются перемешивание химуса и повышение давления в каждом из образующих сегментов кишки. Маятникообразные сокращения обеспечиваются сокращениями продольного слоя мышц с участием циркуляторного. При этих сокращениях происходит перемещение химуса вперед-назад и слабое поступательное движение в аборальном направлении. В проксимальных отделах тонкой кишки частота ритмических сокращений, или циклов, составляет 9-12, в дистальных – 6–8 в 1 мин.
Перистальтика состоит в том, что выше химуса за счет сокращения циркуляторного слоя мышц образуется перехват, а ниже в результате сокращения продольных мышц – расширение полости кишки. Этот перехват и расширение движутся вдоль кишки, перемещая впереди перехвата порцию химуса. По длине кишки одновременно движется несколько перистальтических волн. При антиперистальтических сокращениях волна движется в обратном (оральном) направлении. В норме тонкая кишка антиперистальтически не сокращается. Тонические сокращения могут иметь небольшую скорость, а иногда вообще не распространяться, значительно суживая просвет кишки на большом протяжении.
Выявлена определенная роль моторики в выведении пищеварительных секретов – перистальтика протоков, изменение их тонуса, закрытие и раскрытие их сфинктеров, сокращение и расслабление желчного пузыря. К этому же следует присоединить изменения складчатости слизистой оболочки, микромоторику кишечных ворсинок и микроворсинок тонкой кишки – очень важные явления, оптимизирующие мембранное пищеварение, всасывание нутриентов и других веществ из кишки в кровь и лимфу.
Моторика тонкой кишки регулируется нервными и гуморальными механизмами. Координирующее влияние оказывают интрамуральные (в стенке кишки) нервные образования, а также центральная нервная система. Интрамуральные нейроны обеспечивают координированные сокращения кишки. Особенно велика их роль в перистальтических сокращениях. На интрамуральные механизмы оказывают влияние экстрамуральные, парасимпатические и симпатические нервные механизмы, а также гуморальные факторы.
Моторная активность кишки зависит в том числе от физических и химических свойств химуса. Повышает ее активность грубая пища (черный хлеб, овощи, грубоволокнистые продукты) и жиры. При средней скорости перемещения 1–4 см/мин пища достигает слепой кишки за 2–4 ч. На продолжительность перемещения пищи влияет ее состав, в зависимости от него скорость перемещения уменьшается в ряду: углеводы, белки, жиры.
Гуморальные вещества изменяют моторику кишечника, действуя непосредственно на мышечные волокна и через рецепторы на нейроны интрамуральной нервной системы. Усиливают моторику тонкой кишки вазопрессин, окситоцин, брадикинин, серотонин, гистамин, гастрин, мотилин, холецистокинин-панкреозимин, субстанция Р и ряд других веществ (кислоты, щелочи, соли, продукты переваривания пищевых веществ, особенно жиров).
Защитные системы
Поступление пищи в Ж КТ следует рассматривать не только как способ восполнения энергетических и пластических материалов, но и как аллергическую и токсическую агрессии. Питание связано с опасностью проникновения во внутреннюю среду организма различного рода антигенов и токсических веществ. Особую опасность представляют чужеродные белки. Лишь благодаря сложной системе защиты негативные стороны питания эффективно нейтрализуются. В этих процессах особенно важную роль играет тонкая кишка, осуществляющая несколько жизненно важных функций – пищеварительную, транспортную и барьерную. Именно в тонкой кишке пища подвергается многоступенчатой ферментативной обработке, что необходимо для последующего всасывания и усвоения образующихся продуктов гидролиза пищевых веществ, не имеющих видовой специфичности. Этим организм в определенной мере предохраняет себя от воздействий чужеродных субстанций.
Барьерная, или защитная, функция тонкой кишки зависит от ее макро– и микроструктуры, ферментного спектра, иммунных свойств, слизи, проницаемости и т. д. Слизистая оболочка тонкой кишки участвует в механической, или пассивной, а также в активной защите организма от вредных веществ. Неиммунные и иммунные механизмы защиты тонкой кишки предохраняют внутреннюю среду организма от чужеродных субстанций, антигенов и токсинов. Кислый желудочный сок, пищеварительные ферменты, в том числе протеазы желудочно-кишечного тракта, моторика тонкой кишки, ее микрофлора, слизь, щеточная кайма и гликокаликс апикальной части кишечных клеток относятся к неспецифическим защитным барьерам.
Благодаря ультраструктуре поверхности тонкой кишки, то есть щеточной кайме и гликокаликсу, а также липопротеиновой мембране кишечные клетки служат механическим барьером, препятствующим поступлению антигенов, токсических веществ и других высокомолекулярных соединений из энтеральной среды во внутреннюю. Исключением являются молекулы, подвергающиеся гидролизу ферментами, адсорбированными на структурах гликокаликса. Крупные молекулы и надмолекулярные комплексы не могут проникать в зону щеточной каймы, так как ее поры, или межмикроворсинчатые пространства, чрезвычайно малы. Так, наименьшее расстояние между микроворсинками в среднем составляет 1–2 мкм, а размеры ячеек сети гликокаликса в сотни раз меньше. Таким образом, гликокаликс служит барьером, определяющим проницаемость пищевых веществ, причем апикальная мембрана кишечных клеток благодаря гликокаликсу практически недоступна (или мало доступна) для макромолекул.
К другой механической, или пассивной, системе защиты относятся ограниченная проницаемость слизистой оболочки тонкой кишки для водорастворимых молекул со сравнительно небольшой молекулярной массой и непроницаемость для полимеров, в число которых входят белки, мукополисахариды и другие субстанции, обладающие антигенными свойствами. Однако для клеток пищеварительного аппарата в период раннего постнатального развития характерен эндоцитоз, способствующий поступлению во внутреннюю среду организма макромолекул и чужеродных антигенов. Кишечные клетки взрослых организмов также способны в определенных случаях поглощать крупные молекулы, в том числе нерасщепленные. Кроме того, при прохождении пищи через тонкую кишку образуется значительное количество летучих жирных кислот, одни из которых при всасывании вызывают токсический эффект, а другие – локальное раздражающее действие. Что касается ксенобиотиков, то их образование и всасывание в тонкой кишке варьирует в зависимости от состава, свойств и загрязненности пищи.
Барьерная, или защитная, функция тонкой кишки зависит от ее макро– и микроструктуры, ферментного спектра, иммунных свойств, слизи, проницаемости и т. д. Слизистая оболочка тонкой кишки участвует в механической, или пассивной, а также в активной защите организма от вредных веществ. Неиммунные и иммунные механизмы защиты тонкой кишки предохраняют внутреннюю среду организма от чужеродных субстанций, антигенов и токсинов. Кислый желудочный сок, пищеварительные ферменты, в том числе протеазы желудочно-кишечного тракта, моторика тонкой кишки, ее микрофлора, слизь, щеточная кайма и гликокаликс апикальной части кишечных клеток относятся к неспецифическим защитным барьерам.
Благодаря ультраструктуре поверхности тонкой кишки, то есть щеточной кайме и гликокаликсу, а также липопротеиновой мембране кишечные клетки служат механическим барьером, препятствующим поступлению антигенов, токсических веществ и других высокомолекулярных соединений из энтеральной среды во внутреннюю. Исключением являются молекулы, подвергающиеся гидролизу ферментами, адсорбированными на структурах гликокаликса. Крупные молекулы и надмолекулярные комплексы не могут проникать в зону щеточной каймы, так как ее поры, или межмикроворсинчатые пространства, чрезвычайно малы. Так, наименьшее расстояние между микроворсинками в среднем составляет 1–2 мкм, а размеры ячеек сети гликокаликса в сотни раз меньше. Таким образом, гликокаликс служит барьером, определяющим проницаемость пищевых веществ, причем апикальная мембрана кишечных клеток благодаря гликокаликсу практически недоступна (или мало доступна) для макромолекул.
К другой механической, или пассивной, системе защиты относятся ограниченная проницаемость слизистой оболочки тонкой кишки для водорастворимых молекул со сравнительно небольшой молекулярной массой и непроницаемость для полимеров, в число которых входят белки, мукополисахариды и другие субстанции, обладающие антигенными свойствами. Однако для клеток пищеварительного аппарата в период раннего постнатального развития характерен эндоцитоз, способствующий поступлению во внутреннюю среду организма макромолекул и чужеродных антигенов. Кишечные клетки взрослых организмов также способны в определенных случаях поглощать крупные молекулы, в том числе нерасщепленные. Кроме того, при прохождении пищи через тонкую кишку образуется значительное количество летучих жирных кислот, одни из которых при всасывании вызывают токсический эффект, а другие – локальное раздражающее действие. Что касается ксенобиотиков, то их образование и всасывание в тонкой кишке варьирует в зависимости от состава, свойств и загрязненности пищи.