Страница:
Скорость роста численности в S-образной модели определяет дифференциальное уравнение
dN/dτ = rN(1 – N/K), (4.2)
где K – поддерживающая емкость среды, т. е. максимальный размер популяции, которая может существовать в данных условиях, удовлетворяя свои потребности неопределенно долго.
Если N >K, скорость роста отрицательна. Если N <K, скорость роста положительна и величина популяции N стремится к K, т. е. приводится в соответствие с поддерживающей емкостью среды. Если N = K, скорость роста популяции равна нулю. При нулевом росте популяция стабильна, т. е. ее размеры не меняются, хотя отдельные организмы по-прежнему растут, размножаются и отмирают. Происходящее размножение уравновешивается смертностью.
В специализированной литературе J– и S-образные модели роста численности часто называют соответственно экспоненциальной и логистической.
Поддерживающая емкость играет решающую роль не только при росте популяции по S-образной, но также и по J-образной модели, ибо в некоторый момент времени все же наступает исчерпание какого-либо ресурса среды, т. е. он (или даже несколько одновременно) становится лимитирующим. Развитие дальнейших событий показано на рис. 4.6, а, б. После бума с внезапным выходом J-образной кривой за пределы уровня K происходит крах популяции,[24] т. е. катастрофа, приводящая к резкому снижению численности. Причиной краха часто бывает внезапное резкое изменение условий окружающей среды (экологических факторов), понижающее поддерживающую емкость среды. Тогда огромное число особей, не способных эмигрировать, погибает.
При наиболее благоприятном для популяции стечении обстоятельств новый уровень численности соответствует поддерживающей емкости среды или, иначе говоря, кривая роста превращается из J-образной в S-образную (рис. 4.6, б). Однако исчерпание пищевых ресурсов может привести также к появлению и других трудностей для популяции, например к развитию болезней. Тогда численность снижается до уровня значительно более низкого, чем поддерживающая емкость среды (рис. 4.6, а), а в пределе популяция может даже быть обречена на вымирание.
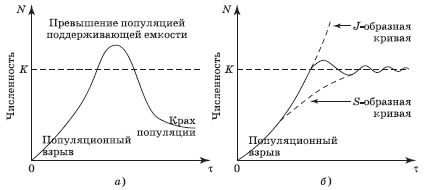 Рис. 4.6. Варианты завершения роста популяции по J-образной модели: а – для дафнии на питательной среде; б – преобразование в S-образный вид
Рис. 4.6. Варианты завершения роста популяции по J-образной модели: а – для дафнии на питательной среде; б – преобразование в S-образный вид
Для S-образной модели в случаях отставания действия регулирующих механизмов по каким-либо причинам, например, в связи с затратами времени на воспроизводство или по иным причинам временное запаздывание учитывает дифференциальное уравнение
N/dτ = rNK – rN 2(τ– T)/K, (4.3)
где T – время, необходимое системе для реакции на внешнее воздействие.
Вычитаемое в правой части уравнения, содержащее N2, позволяет предсказать момент выхода системы из состояния равновесия в случаях, когда время запаздывания относительно велико по сравнению с временем релаксации (1/r) системы. В итоге при увеличении в системе времени запаздывания вместо асимптотического приближения к состоянию равновесия происходит колебание численности организмов относительно теоретической S-образной кривой. В случаях, когда пищевые ресурсы ограничены, популяция не достигает устойчивого равновесия, ибо численность одного поколения зависит от численности другого, что отражается на скорости репродукции и приводит к хищничеству и каннибализму. Колебания численности популяции, для которой характерны большие значения r, малое время воспроизводства τ и несложный регулирующий механизм, могут быть весьма значительными.
Описанные модели роста популяции и дифференциальные уравнения предполагают, что все организмы сходны между собой, имеют равную вероятность погибнуть и равную способность к размножению, так что скорость роста популяции в экспоненциальной фазе зависит только от ее численности и не ограничена условиями среды, которые остаются постоянными. Они точно описывают процессы роста и взаимодействия особей в большинстве искусственных и некоторых естественных популяциях. «Идеальность» всех экологических факторов в исходных условиях предопределила то, что рассматриваемые модели называют идеальными.
Для природных популяций принятые допущения чаще всего неверны. В естественных условиях J-и S-образные модели роста популяции преимущественно можно наблюдать в случаях, когда тех или иных животных вселяют или они сами распространяются в новые для них районы. Тем не менее теоретические модели роста позволяют лучше понять процессы, происходящие в естественных условиях. Большинство принципов, используемых для моделирования популяций животных, применимо также и для моделирования популяций растений.
Следует отметить, что при любой модели (как J-, так и S-образной) вначале характерна фаза экспоненциального роста численности популяции (рис. 4.5, 4.6, б). Поэтому при сочетании благоприятных (оптимальных) значений всех факторов среды возникает «популяционный взрыв», т. е. особо быстрый рост популяции того или иного вида.
Миграция или расселение, так же как и внезапное снижение скорости размножения, могут способствовать уменьшению численности популяции. Расселение может быть связано с определенной стадией жизненного цикла, например с образованием семян.
Применительно к условиям реальной природной среды принято использовать понятия биотический потенциал – совокупность всех экологических факторов, способствующих увеличению численности популяции, или видовая способность к размножению при отсутствии ограничений со стороны среды, а также сопротивление среды – сочетание факторов, ограничивающих рост (лимитирующих факторов).
4.4.3. Колебания численности популяции
4.5. Популяции синантропных видов
Контрольные вопросы и задания
ГЛАВА 5
5.1. Трофическая структура биоценозов
5.1.1. Пищевые цепи и сети
5.1.2. Экологические пирамиды
5.1.2.1. Пирамида численности
5.1.2.2. Пирамида биомасс
dN/dτ = rN(1 – N/K), (4.2)
где K – поддерживающая емкость среды, т. е. максимальный размер популяции, которая может существовать в данных условиях, удовлетворяя свои потребности неопределенно долго.
Если N >K, скорость роста отрицательна. Если N <K, скорость роста положительна и величина популяции N стремится к K, т. е. приводится в соответствие с поддерживающей емкостью среды. Если N = K, скорость роста популяции равна нулю. При нулевом росте популяция стабильна, т. е. ее размеры не меняются, хотя отдельные организмы по-прежнему растут, размножаются и отмирают. Происходящее размножение уравновешивается смертностью.
В специализированной литературе J– и S-образные модели роста численности часто называют соответственно экспоненциальной и логистической.
Поддерживающая емкость играет решающую роль не только при росте популяции по S-образной, но также и по J-образной модели, ибо в некоторый момент времени все же наступает исчерпание какого-либо ресурса среды, т. е. он (или даже несколько одновременно) становится лимитирующим. Развитие дальнейших событий показано на рис. 4.6, а, б. После бума с внезапным выходом J-образной кривой за пределы уровня K происходит крах популяции,[24] т. е. катастрофа, приводящая к резкому снижению численности. Причиной краха часто бывает внезапное резкое изменение условий окружающей среды (экологических факторов), понижающее поддерживающую емкость среды. Тогда огромное число особей, не способных эмигрировать, погибает.
При наиболее благоприятном для популяции стечении обстоятельств новый уровень численности соответствует поддерживающей емкости среды или, иначе говоря, кривая роста превращается из J-образной в S-образную (рис. 4.6, б). Однако исчерпание пищевых ресурсов может привести также к появлению и других трудностей для популяции, например к развитию болезней. Тогда численность снижается до уровня значительно более низкого, чем поддерживающая емкость среды (рис. 4.6, а), а в пределе популяция может даже быть обречена на вымирание.
Для S-образной модели в случаях отставания действия регулирующих механизмов по каким-либо причинам, например, в связи с затратами времени на воспроизводство или по иным причинам временное запаздывание учитывает дифференциальное уравнение
N/dτ = rNK – rN 2(τ– T)/K, (4.3)
где T – время, необходимое системе для реакции на внешнее воздействие.
Вычитаемое в правой части уравнения, содержащее N2, позволяет предсказать момент выхода системы из состояния равновесия в случаях, когда время запаздывания относительно велико по сравнению с временем релаксации (1/r) системы. В итоге при увеличении в системе времени запаздывания вместо асимптотического приближения к состоянию равновесия происходит колебание численности организмов относительно теоретической S-образной кривой. В случаях, когда пищевые ресурсы ограничены, популяция не достигает устойчивого равновесия, ибо численность одного поколения зависит от численности другого, что отражается на скорости репродукции и приводит к хищничеству и каннибализму. Колебания численности популяции, для которой характерны большие значения r, малое время воспроизводства τ и несложный регулирующий механизм, могут быть весьма значительными.
Описанные модели роста популяции и дифференциальные уравнения предполагают, что все организмы сходны между собой, имеют равную вероятность погибнуть и равную способность к размножению, так что скорость роста популяции в экспоненциальной фазе зависит только от ее численности и не ограничена условиями среды, которые остаются постоянными. Они точно описывают процессы роста и взаимодействия особей в большинстве искусственных и некоторых естественных популяциях. «Идеальность» всех экологических факторов в исходных условиях предопределила то, что рассматриваемые модели называют идеальными.
Для природных популяций принятые допущения чаще всего неверны. В естественных условиях J-и S-образные модели роста популяции преимущественно можно наблюдать в случаях, когда тех или иных животных вселяют или они сами распространяются в новые для них районы. Тем не менее теоретические модели роста позволяют лучше понять процессы, происходящие в естественных условиях. Большинство принципов, используемых для моделирования популяций животных, применимо также и для моделирования популяций растений.
Следует отметить, что при любой модели (как J-, так и S-образной) вначале характерна фаза экспоненциального роста численности популяции (рис. 4.5, 4.6, б). Поэтому при сочетании благоприятных (оптимальных) значений всех факторов среды возникает «популяционный взрыв», т. е. особо быстрый рост популяции того или иного вида.
Миграция или расселение, так же как и внезапное снижение скорости размножения, могут способствовать уменьшению численности популяции. Расселение может быть связано с определенной стадией жизненного цикла, например с образованием семян.
Применительно к условиям реальной природной среды принято использовать понятия биотический потенциал – совокупность всех экологических факторов, способствующих увеличению численности популяции, или видовая способность к размножению при отсутствии ограничений со стороны среды, а также сопротивление среды – сочетание факторов, ограничивающих рост (лимитирующих факторов).
Любые изменения популяции есть результат нарушения равновесия между ее биотическим потенциалом и сопротивлением окружающей среды.
4.4.3. Колебания численности популяции
По достижении заключительной фазы роста размеры популяции продолжают колебаться от поколения к поколению вокруг некоторой более или менее постоянной величины. При этом численность одних видов изменяется нерегулярно с большой амплитудой колебаний (насекомые-вредители, сорняки), колебания численности других (например, мелких млекопитающих) имеют относительно постоянный период, а в популяциях третьих видов численность колеблется от года к году незначительно (долгоживущие крупные позвоночные и древесные растения).
В природе в основном встречаются три вида кривых изменения численности популяции: относительно стабильный, скачкообразный и циклический (рис. 4.7).
Виды, у которых численность из года в год находится на уровне поддерживающей емкости среды, имеют достаточно стабильные популяции (кривая 1). Такое постоянство характерно для многих видов дикой природы и встречается, например, в нетронутых тропических влажных лесах, где среднегодовое количество осадков и температура изменяются день ото дня и из года в год крайне мало.
У других видов колебания численности популяций носят правильный циклический характер (кривая 2). Хорошо знакомы примеры сезонных колебаний численности. Тучи комаров; поля, заросшие цветами; леса, полные птиц, – все это характерно для теплого времени года в средней полосе и сходит практически на нет зимой.
Широко известен пример циклических колебаний численности леммингов (северных травоядных мышевидных грызунов) в Северной Америке и Скандинавии. Раз в четыре года плотность их популяций становится столь высокой, что они начинают мигрировать со своих перенаселенных местообитаний; при этом массово гибнут в фиордах и тонут в реках, что до настоящего времени не имеет достаточного объяснения. Еще с глубокой древности известны циклические нашествия странствующей африканской саранчи на Евразию.
Ряд видов, таких, как енот, в основном имеют достаточно стабильные популяции, однако время от времени их численность резко возрастает (подскакивает) до наивысшего значения, а затем резко падает до некоторого низкого, но относительно стабильного уровня. Эти виды относят к популяциям со скачкообразным ростом численности (кривая 3).
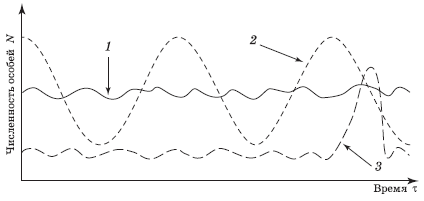 Рис. 4.7. Основные кривые изменения численности популяций различных видов: 1 – стабильный; 2 – цикличный; 3 – скачкообразный
Рис. 4.7. Основные кривые изменения численности популяций различных видов: 1 – стабильный; 2 – цикличный; 3 – скачкообразный
Внезапное увеличение численности происходит при временном повышении емкости среды для данной популяции и может быть связано с улучшением климатических условий (факторов) и питания или резким уменьшением численности хищников (включая охотников). После превышения новой, более высокой емкости среды в популяции возрастает смертность и ее размеры резко сокращаются.
На протяжении истории в разных странах не раз наблюдались случаи краха популяций человека, например, в Ирландии в 1845 г., когда в результате заражения грибком погиб весь урожай картофеля. Поскольку рацион питания ирландцев сильно зависел от картофеля, к 1900 г. половина восьмимиллионного населения Ирландии умерла от голода или эмигрировала в другие страны.
Тем не менее численность человечества на Земле в целом и во многих регионах в частности продолжает расти. Люди путем технологических, социальных и культурных перемен неоднократно увеличивали для себя поддерживающую емкость планеты (рис. 4.8). По сути, они смогли изменить свою экологическую нишу за счет увеличения производства продуктов питания, борьбы с болезнями и использования больших количеств энергетических и материальных ресурсов, чтобы сделать обычно непригодные для жизни районы Земли обитаемыми.
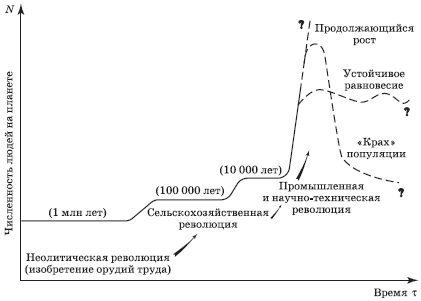 Рис. 4.8. Увеличение поддерживающей емкости среды для популяции человека (по Т. Миллеру), масштаб по осям условный
Рис. 4.8. Увеличение поддерживающей емкости среды для популяции человека (по Т. Миллеру), масштаб по осям условный
В правой части рис. 4.8 приведены возможные сценарии дальнейшего изменения фактической численности людей на планете в случае превышения поддерживающей емкости биосферы.
В природе в основном встречаются три вида кривых изменения численности популяции: относительно стабильный, скачкообразный и циклический (рис. 4.7).
Виды, у которых численность из года в год находится на уровне поддерживающей емкости среды, имеют достаточно стабильные популяции (кривая 1). Такое постоянство характерно для многих видов дикой природы и встречается, например, в нетронутых тропических влажных лесах, где среднегодовое количество осадков и температура изменяются день ото дня и из года в год крайне мало.
У других видов колебания численности популяций носят правильный циклический характер (кривая 2). Хорошо знакомы примеры сезонных колебаний численности. Тучи комаров; поля, заросшие цветами; леса, полные птиц, – все это характерно для теплого времени года в средней полосе и сходит практически на нет зимой.
Широко известен пример циклических колебаний численности леммингов (северных травоядных мышевидных грызунов) в Северной Америке и Скандинавии. Раз в четыре года плотность их популяций становится столь высокой, что они начинают мигрировать со своих перенаселенных местообитаний; при этом массово гибнут в фиордах и тонут в реках, что до настоящего времени не имеет достаточного объяснения. Еще с глубокой древности известны циклические нашествия странствующей африканской саранчи на Евразию.
Ряд видов, таких, как енот, в основном имеют достаточно стабильные популяции, однако время от времени их численность резко возрастает (подскакивает) до наивысшего значения, а затем резко падает до некоторого низкого, но относительно стабильного уровня. Эти виды относят к популяциям со скачкообразным ростом численности (кривая 3).
Внезапное увеличение численности происходит при временном повышении емкости среды для данной популяции и может быть связано с улучшением климатических условий (факторов) и питания или резким уменьшением численности хищников (включая охотников). После превышения новой, более высокой емкости среды в популяции возрастает смертность и ее размеры резко сокращаются.
На протяжении истории в разных странах не раз наблюдались случаи краха популяций человека, например, в Ирландии в 1845 г., когда в результате заражения грибком погиб весь урожай картофеля. Поскольку рацион питания ирландцев сильно зависел от картофеля, к 1900 г. половина восьмимиллионного населения Ирландии умерла от голода или эмигрировала в другие страны.
Тем не менее численность человечества на Земле в целом и во многих регионах в частности продолжает расти. Люди путем технологических, социальных и культурных перемен неоднократно увеличивали для себя поддерживающую емкость планеты (рис. 4.8). По сути, они смогли изменить свою экологическую нишу за счет увеличения производства продуктов питания, борьбы с болезнями и использования больших количеств энергетических и материальных ресурсов, чтобы сделать обычно непригодные для жизни районы Земли обитаемыми.
В правой части рис. 4.8 приведены возможные сценарии дальнейшего изменения фактической численности людей на планете в случае превышения поддерживающей емкости биосферы.
4.5. Популяции синантропных видов
В предыдущих разделах рассмотрены природные популяции, находящиеся в естественных местообитаниях. Однако вследствие хозяйственной деятельности человека образуются природно-антропогенные популяции, тесно связанные, например, с сельским хозяйством. Многие насекомые, мышевидные грызуны и прочие виды находят здесь экологическую нишу, адаптируя свою структуру и динамику численности к той или иной системе хозяйствования.
С эволюцией человеческого общества и образованием стабильных поселений также возникли с и н а н т р о п н ы е в и д ы, популяции которых обитают в жилищах и в местах скопления отходов жизнедеятельности людей. Они могут быть «вредителями», «паразитами», переносчиками возбудителей болезней, выступая в качестве опасных экологических (биотических) факторов для человека.
В пещеру древнего человека перешли на жительство многие насекомые, осваивая скопления различных отходов. Однако человечество постепенно развивало культуру быта и улучшало санитарно-гигиеническую обстановку в своих поселениях, одновременно создавая разнообразные методы уничтожения нежелательных «спутников». В то же время насекомых отпугивала сама обстановка города, с течением времени все больше отличавшаяся от привычной природной среды. В результате большинство насекомых отступило, но тараканы, комнатные мухи, некоторые виды муравьев и другие преодолели все биологические преграды и стали «хозяйничать» в городской и поселковой среде.
Мухи активны в квартирах днем, а ночью их сменяют тараканы – представители крупного одноименного отряда насекомых. Синантропные тараканы – теплолюбивые насекомые, происходящие из тропических лесов Южной Азии. В северных районах они живут только в отапливаемых помещениях. Понижение температуры ниже +5 °C для тараканов смертельно, поэтому их не бывает в дачных домиках в средней полосе России, а в природных условиях тараканы встречаются в Крыму, на юге Дальнего Востока и в южных районах Средней Азии. Но даже в теплом помещении они не будут жить при отсутствии источника воды.
Тараканы распространяются по городу или поселку чаще всего при перевозках продуктов или комнатных вещей, а в теплое время года могут переходить из дома в дом самостоятельно.
Вместе с мухами и тараканами в домах поселяются муравьи. В деревянных домах в сельской местности живут муравьи-древоточцы, обитающие и в окрестных лесах. В городах стал обычным мелкий рыжеватый муравей, в естественных условиях встречающийся только в тропиках.
Все комнатные насекомые, посещающие различные отбросы, переносят на теле возбудителей опасных заболеваний и создают антисанитарные условия для людей.
С эволюцией человеческого общества и образованием стабильных поселений также возникли с и н а н т р о п н ы е в и д ы, популяции которых обитают в жилищах и в местах скопления отходов жизнедеятельности людей. Они могут быть «вредителями», «паразитами», переносчиками возбудителей болезней, выступая в качестве опасных экологических (биотических) факторов для человека.
В пещеру древнего человека перешли на жительство многие насекомые, осваивая скопления различных отходов. Однако человечество постепенно развивало культуру быта и улучшало санитарно-гигиеническую обстановку в своих поселениях, одновременно создавая разнообразные методы уничтожения нежелательных «спутников». В то же время насекомых отпугивала сама обстановка города, с течением времени все больше отличавшаяся от привычной природной среды. В результате большинство насекомых отступило, но тараканы, комнатные мухи, некоторые виды муравьев и другие преодолели все биологические преграды и стали «хозяйничать» в городской и поселковой среде.
Мухи активны в квартирах днем, а ночью их сменяют тараканы – представители крупного одноименного отряда насекомых. Синантропные тараканы – теплолюбивые насекомые, происходящие из тропических лесов Южной Азии. В северных районах они живут только в отапливаемых помещениях. Понижение температуры ниже +5 °C для тараканов смертельно, поэтому их не бывает в дачных домиках в средней полосе России, а в природных условиях тараканы встречаются в Крыму, на юге Дальнего Востока и в южных районах Средней Азии. Но даже в теплом помещении они не будут жить при отсутствии источника воды.
Тараканы распространяются по городу или поселку чаще всего при перевозках продуктов или комнатных вещей, а в теплое время года могут переходить из дома в дом самостоятельно.
Вместе с мухами и тараканами в домах поселяются муравьи. В деревянных домах в сельской местности живут муравьи-древоточцы, обитающие и в окрестных лесах. В городах стал обычным мелкий рыжеватый муравей, в естественных условиях встречающийся только в тропиках.
Все комнатные насекомые, посещающие различные отбросы, переносят на теле возбудителей опасных заболеваний и создают антисанитарные условия для людей.
Контрольные вопросы и задания
4.1. Дайте определение популяции и ее свойств.
4.2. Почему элементарной частицей эволюции является популяция?
4.3. Сформулируйте правило Ю. Одума и теорию К. Фридерихса.
4.4. Каково место популяции на Земле?
4.5. Что отражают статистические показатели популяции?
4.6. Почему толерантность популяции к факторам среды значительно шире, чем у особи, и каково экологическое значение этого явления?
4.7. Каковы экологические причины, вызывающие рост численности популяций по экспоненте и логистической кривой?
4.8. В чем суть экологической стратегии выживания?
4.9. Какие экологические факторы вызывают саморегуляцию плотности популяции?
4.10. Что такое синантропные виды? Почему они являются «опасными» видами для человека?
4.2. Почему элементарной частицей эволюции является популяция?
4.3. Сформулируйте правило Ю. Одума и теорию К. Фридерихса.
4.4. Каково место популяции на Земле?
4.5. Что отражают статистические показатели популяции?
4.6. Почему толерантность популяции к факторам среды значительно шире, чем у особи, и каково экологическое значение этого явления?
4.7. Каковы экологические причины, вызывающие рост численности популяций по экспоненте и логистической кривой?
4.8. В чем суть экологической стратегии выживания?
4.9. Какие экологические факторы вызывают саморегуляцию плотности популяции?
4.10. Что такое синантропные виды? Почему они являются «опасными» видами для человека?
ГЛАВА 5
ЭКОЛОГИЯ СООБЩЕСТВ (СИНЭКОЛОГИЯ)
Популяции разных видов в природных условиях объединяются в системы более высокого ранга – сообщества и биоценоз.[25]
Термин «биоценоз» был предложен немецким зоологом К. Мебиусом и обозначает организованную группу популяций растений, животных и микроорганизмов, приспособленных к совместному обитанию в пределах определенного объема пространства.
Любой биоценоз занимает определенный участок абиотической среды. Биотоп – пространство с более или менее однородными условиями, заселенное тем или иным сообществом организмов.
Размеры биоценотических группировок организмов чрезвычайно разнообразны – от сообществ на стволе дерева или на болотной моховой кочке до биоценоза ковыльной степи. Биоценоз (сообщество) – не просто сумма образующих его видов, но и совокупность взаимодействий между ними. Экология сообществ (синэкология)[26] – это также научный подход в экологии, в соответствии с которым прежде всего исследуют комплекс отношений и господствующие взаимосвязи в биоценозе. Синэкология занимается преимущественно биотическими экологическими факторами среды.
В пределах биоценоза различают фитоценоз – устойчивое сообщество растительных организмов, зооценоз – совокупность взаимосвязанных видов животных и микробиоценоз – сообщество микроорганизмов:
ФИТОЦЕНОЗ + ЗООЦЕНОЗ + МИКРОБИОЦЕНОЗ = БИОЦЕНОЗ.
При этом в чистом виде ни фитоценоз, ни зооценоз, ни микробиоценоз в природе не встречаются, как и биоценоз в отрыве от биотопа.
Биоценоз формируют межвидовые связи, обеспечивающие структуру биоценоза – численность особей, распределение их в пространстве, видовой состав и тому подобное, а также структуру пищевой сети, продуктивность и биомассу. Для оценки роли отдельного вида в видовой структуре биоценоза используют обилие вида – показатель, равный числу особей на единицу площади или объема занимаемого пространства.
Термин «биоценоз» был предложен немецким зоологом К. Мебиусом и обозначает организованную группу популяций растений, животных и микроорганизмов, приспособленных к совместному обитанию в пределах определенного объема пространства.
Любой биоценоз занимает определенный участок абиотической среды. Биотоп – пространство с более или менее однородными условиями, заселенное тем или иным сообществом организмов.
Размеры биоценотических группировок организмов чрезвычайно разнообразны – от сообществ на стволе дерева или на болотной моховой кочке до биоценоза ковыльной степи. Биоценоз (сообщество) – не просто сумма образующих его видов, но и совокупность взаимодействий между ними. Экология сообществ (синэкология)[26] – это также научный подход в экологии, в соответствии с которым прежде всего исследуют комплекс отношений и господствующие взаимосвязи в биоценозе. Синэкология занимается преимущественно биотическими экологическими факторами среды.
В пределах биоценоза различают фитоценоз – устойчивое сообщество растительных организмов, зооценоз – совокупность взаимосвязанных видов животных и микробиоценоз – сообщество микроорганизмов:
ФИТОЦЕНОЗ + ЗООЦЕНОЗ + МИКРОБИОЦЕНОЗ = БИОЦЕНОЗ.
При этом в чистом виде ни фитоценоз, ни зооценоз, ни микробиоценоз в природе не встречаются, как и биоценоз в отрыве от биотопа.
Биоценоз формируют межвидовые связи, обеспечивающие структуру биоценоза – численность особей, распределение их в пространстве, видовой состав и тому подобное, а также структуру пищевой сети, продуктивность и биомассу. Для оценки роли отдельного вида в видовой структуре биоценоза используют обилие вида – показатель, равный числу особей на единицу площади или объема занимаемого пространства.
5.1. Трофическая структура биоценозов
Важнейший вид взаимоотношений между организмами в биоценозе, фактически формирующими его структуру, – это пищевые связи хищника и жертвы: одни – поедающие, другие – поедаемые. При этом все организмы, живые и мертвые, являются пищей для других организмов: заяц ест траву, лиса и волк охотятся на зайцев, хищные птицы (ястребы, орлы и т. п.) способны утащить и съесть как лисенка, так и волчонка. Погибшие растения, зайцы, лисы, волки, птицы становятся пищей для детритофагов (редуцентов или иначе деструкторов).
5.1.1. Пищевые цепи и сети
Пищевая цепь – это последовательность организмов, в которой каждый из них съедает или разлагает другой. Она представляет собой путь движущегося через живые организмы однонаправленного потока поглощенной при фотосинтезе малой части высокоэффективной солнечной энергии, поступившей на Землю. В конечном итоге эта цепь возвращается в окружающую природную среду в виде низкоэффективной тепловой энергии. По ней также движутся питательные вещества от продуцентов к консументам и далее к редуцентам, а затем обратно к продуцентам.
Каждое звено пищевой цепи называют трофическим уровнем. Первый трофический уровень занимают автотрофы, иначе именуемые первичными продуцентами. Организмы второго трофического уровня называют первичными консументами, третьего – вторичными консументами и т. д. Обычно бывают четыре или пять трофических уровней и редко более шести (рис. 5.1).
Существуют два главных типа пищевых цепей – пастбищные (или «выедания») и детритные (или «разложения»).
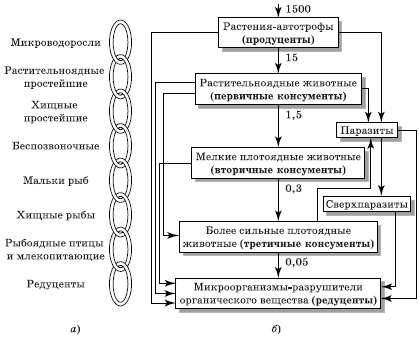 Рис. 5.1. Пищевые цепи биоценоза по Н. Ф. Реймерсу: обобщенная (а) и реальная (б). Стрелками показано направление перемещения энергии, а цифрами – относительное количество энергии, приходящей на трофический уровень
Рис. 5.1. Пищевые цепи биоценоза по Н. Ф. Реймерсу: обобщенная (а) и реальная (б). Стрелками показано направление перемещения энергии, а цифрами – относительное количество энергии, приходящей на трофический уровень
В пастбищных пищевых цепях первый трофический уровень занимают зеленые растения, второй – пастбищные животные (термин «пастбищные» охватывает все организмы, питающиеся растениями), а третий – хищники. Так, пастбищными пищевыми цепями являются:
 Детритная пищевая цепь начинается с детрита по схеме:
Детритная пищевая цепь начинается с детрита по схеме:
ДЕТРИТ → ДЕТРИТОФАГ → ХИЩНИК
Характерными детритными пищевыми цепями являются:
 Концепция пищевых цепей позволяет в дальнейшем проследить круговорот химических элементов в природе, хотя простые пищевые цепи, подобные изображенным ранее, где каждый организм представлен как питающийся организмами только какого-то одного типа, в природе встречаются редко. Реальные пищевые связи намного сложнее, ибо животное может питаться организмами разных типов, входящих в одну и ту же пищевую цепь или в различные цепи, что особенно характерно для хищников (консументов) высших трофических уровней. Связь между пастбищной и детритной пищевыми цепями иллюстрирует предложенная Ю. Одумом модель потока энергии (рис. 5.2).
Концепция пищевых цепей позволяет в дальнейшем проследить круговорот химических элементов в природе, хотя простые пищевые цепи, подобные изображенным ранее, где каждый организм представлен как питающийся организмами только какого-то одного типа, в природе встречаются редко. Реальные пищевые связи намного сложнее, ибо животное может питаться организмами разных типов, входящих в одну и ту же пищевую цепь или в различные цепи, что особенно характерно для хищников (консументов) высших трофических уровней. Связь между пастбищной и детритной пищевыми цепями иллюстрирует предложенная Ю. Одумом модель потока энергии (рис. 5.2).
Всеядные животные (в частности, человек) питаются и консументами, и продуцентами. Таким образом, в природе пищевые цепи переплетаются, образуют пищевые (трофические) сети.
Каждое звено пищевой цепи называют трофическим уровнем. Первый трофический уровень занимают автотрофы, иначе именуемые первичными продуцентами. Организмы второго трофического уровня называют первичными консументами, третьего – вторичными консументами и т. д. Обычно бывают четыре или пять трофических уровней и редко более шести (рис. 5.1).
Существуют два главных типа пищевых цепей – пастбищные (или «выедания») и детритные (или «разложения»).
В пастбищных пищевых цепях первый трофический уровень занимают зеленые растения, второй – пастбищные животные (термин «пастбищные» охватывает все организмы, питающиеся растениями), а третий – хищники. Так, пастбищными пищевыми цепями являются:
ДЕТРИТ → ДЕТРИТОФАГ → ХИЩНИК
Характерными детритными пищевыми цепями являются:
Всеядные животные (в частности, человек) питаются и консументами, и продуцентами. Таким образом, в природе пищевые цепи переплетаются, образуют пищевые (трофические) сети.
5.1.2. Экологические пирамиды
Для наглядности представления взаимоотношений между организмами различных видов в биоценозе принято использовать экологические пирамиды, различая пирамиды численности, биомасс и энергии.
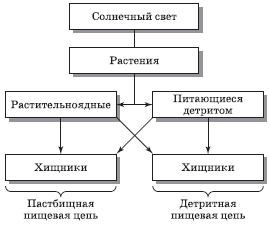 Рис. 5.2. Схема пастбищной и детритной пищевых цепей (по Ю. Одуму)
Рис. 5.2. Схема пастбищной и детритной пищевых цепей (по Ю. Одуму)
5.1.2.1. Пирамида численности
Для построения пирамиды численности подсчитывают число организмов на некоторой территории, группируя их по трофическим уровням:
• продуценты – зеленые растения;
• первичные консументы – травоядные животные;
• вторичные консументы – плотоядные животные;
• третичные консументы – плотоядные животные;
• n-е консументы («конечные хищники») – плотоядные животные;
• редуценты – деструкторы.
Консументы второго, третьего и более высоких порядков могут быть хищниками (охотиться, схватывая и убивая жертву), могут питаться падалью или быть паразитами. В последнем случае они по величине меньше своих хозяев, в результате чего пищевые цепи паразитов необычны по ряду параметров. В типичных пищевых цепях хищников плотоядные животные становятся крупнее на каждом трофическом уровне.
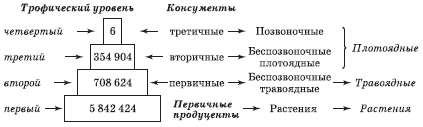 Рис. 5.3. Экологическая пирамида численности для луга, поросшего злаками: цифры – число особей
Рис. 5.3. Экологическая пирамида численности для луга, поросшего злаками: цифры – число особей
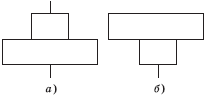 Рис. 5.4. Нарушенная (а) и перевернутая (б) пирамиды численности
Рис. 5.4. Нарушенная (а) и перевернутая (б) пирамиды численности
Каждый уровень изображается условно в виде прямоугольника, длина или площадь которого соответствуют численному значению количества особей. Расположив эти прямоугольники в соподчиненной последовательности, получают экологическую пирамиду численности (рис. 5.3), основной принцип построения которой впервые сформулировал американский эколог Ч. Элтон.
Данные для пирамид численности получают достаточно легко путем прямого сбора образцов, однако существуют и некоторые трудности:
• продуценты сильно различаются по размерам, хотя один экземпляр злака или водоросли имеет одинаковый статус с одним деревом. Это порой нарушает правильную пирамидальную форму, иногда давая даже перевернутые пирамиды (рис. 5.4);
• диапазон численности различных видов настолько широк, что при графическом изображении затрудняет соблюдение масштаба, однако в таких случаях можно использовать логарифмическую шкалу.
• продуценты – зеленые растения;
• первичные консументы – травоядные животные;
• вторичные консументы – плотоядные животные;
• третичные консументы – плотоядные животные;
• n-е консументы («конечные хищники») – плотоядные животные;
• редуценты – деструкторы.
Консументы второго, третьего и более высоких порядков могут быть хищниками (охотиться, схватывая и убивая жертву), могут питаться падалью или быть паразитами. В последнем случае они по величине меньше своих хозяев, в результате чего пищевые цепи паразитов необычны по ряду параметров. В типичных пищевых цепях хищников плотоядные животные становятся крупнее на каждом трофическом уровне.
Каждый уровень изображается условно в виде прямоугольника, длина или площадь которого соответствуют численному значению количества особей. Расположив эти прямоугольники в соподчиненной последовательности, получают экологическую пирамиду численности (рис. 5.3), основной принцип построения которой впервые сформулировал американский эколог Ч. Элтон.
Данные для пирамид численности получают достаточно легко путем прямого сбора образцов, однако существуют и некоторые трудности:
• продуценты сильно различаются по размерам, хотя один экземпляр злака или водоросли имеет одинаковый статус с одним деревом. Это порой нарушает правильную пирамидальную форму, иногда давая даже перевернутые пирамиды (рис. 5.4);
• диапазон численности различных видов настолько широк, что при графическом изображении затрудняет соблюдение масштаба, однако в таких случаях можно использовать логарифмическую шкалу.
5.1.2.2. Пирамида биомасс
Экологическую пирамиду биомасс строят аналогично пирамиде численности. Ее основное значение состоит в том, чтобы показывать количество живого вещества (биомассу – суммарную массу организмов) на каждом трофическом уровне. Это позволяет избежать неудобств, характерных для пирамид численности. В этом случае размер прямоугольников пропорционален массе живого вещества соответствующего уровня, отнесенной к единице площади или объема (рис. 5.5, а, б). Термин «пирамида биомасс» возник в связи с тем, что в абсолютном большинстве случаев масса первичных консументов, живущих за счет продуцентов, значительно меньше массы этих продуцентов, а масса вторичных консументов значительно меньше массы первичных консументов. Биомассу деструкторов принято показывать отдельно.
При отборе образцов определяют биомассу на корню или урожай на корню (т. е. в данный момент времени), которая не содержит никакой информации о скорости образования или потребления биомассы.
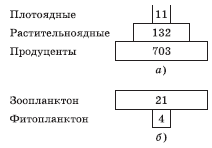 Рис. 5.5. Пирамиды биомасс биоценозов кораллового рифа (а) и пролива Ла-Манш (б): цифры – биомасса в граммах сухого вещества, приходящегося на 1 м2
Рис. 5.5. Пирамиды биомасс биоценозов кораллового рифа (а) и пролива Ла-Манш (б): цифры – биомасса в граммах сухого вещества, приходящегося на 1 м2
Скорость создания органического вещества не определяет его суммарные запасы, т. е. общую биомассу всех организмов каждого трофического уровня. Поэтому при дальнейшем анализе могут возникнуть ошибки, если не учитывать следующее:
• во-первых, при равенстве скорости потребления биомассы (потеря из-за поедания) и скорости ее образования урожай на корню не свидетельствует о продуктивности, т. е. о количестве энергии и вещества, переходящих с одного трофического уровня на другой, более высокий, за некоторый период времени (например, за год). Так, на плодородном, интенсивно используемом пастбище урожай трав на корню может быть ниже, а продуктивность выше, чем на менее плодородном, но мало используемом для выпаса;
• во-вторых, продуцентам небольших размеров, например водорослям, свойственна высокая скорость роста и размножения, уравновешиваемая интенсивным потреблением их в пищу другими организмами и естественной гибелью. Поэтому продуктивность их может быть не меньше чем у крупных продуцентов (например, деревьев), хотя на корню биомасса может быть мала. Иными словами, фитопланктон с такой же продуктивностью, как у дерева, будет иметь намного меньшую биомассу, хотя мог бы поддерживать жизнь животных такой же массы.
Одним из следствий описанного являются «перевернутые пирамиды» (рис. 5.5, б). Зоопланктон биоценозов озер и морей чаще всего обладает большей биомассой, чем его пища – фитопланктон, однако скорость размножения зеленых водорослей настолько велика, что в течение суток они восстанавливают всю съеденную зоопланктоном биомассу. Тем не менее в определенные периоды года (во время весеннего цветения) наблюдают обычное соотношение их биомасс (рис. 5.6).
При отборе образцов определяют биомассу на корню или урожай на корню (т. е. в данный момент времени), которая не содержит никакой информации о скорости образования или потребления биомассы.
Скорость создания органического вещества не определяет его суммарные запасы, т. е. общую биомассу всех организмов каждого трофического уровня. Поэтому при дальнейшем анализе могут возникнуть ошибки, если не учитывать следующее:
• во-первых, при равенстве скорости потребления биомассы (потеря из-за поедания) и скорости ее образования урожай на корню не свидетельствует о продуктивности, т. е. о количестве энергии и вещества, переходящих с одного трофического уровня на другой, более высокий, за некоторый период времени (например, за год). Так, на плодородном, интенсивно используемом пастбище урожай трав на корню может быть ниже, а продуктивность выше, чем на менее плодородном, но мало используемом для выпаса;
• во-вторых, продуцентам небольших размеров, например водорослям, свойственна высокая скорость роста и размножения, уравновешиваемая интенсивным потреблением их в пищу другими организмами и естественной гибелью. Поэтому продуктивность их может быть не меньше чем у крупных продуцентов (например, деревьев), хотя на корню биомасса может быть мала. Иными словами, фитопланктон с такой же продуктивностью, как у дерева, будет иметь намного меньшую биомассу, хотя мог бы поддерживать жизнь животных такой же массы.
Одним из следствий описанного являются «перевернутые пирамиды» (рис. 5.5, б). Зоопланктон биоценозов озер и морей чаще всего обладает большей биомассой, чем его пища – фитопланктон, однако скорость размножения зеленых водорослей настолько велика, что в течение суток они восстанавливают всю съеденную зоопланктоном биомассу. Тем не менее в определенные периоды года (во время весеннего цветения) наблюдают обычное соотношение их биомасс (рис. 5.6).