Страница:
Кажущихся аномалий лишены пирамиды энергий, рассматриваемые далее.
5.1.2.3. Пирамида энергий
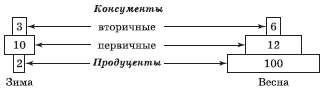
Самым фундаментальным способом отражения связей между организмами разных трофических уровней и функциональной организации биоценозов является п и р а м и д а энергий, в которой размер прямоугольников пропорционален энергетическому эквиваленту в единицу времени, т. е. количеству энергии (на единицу площади или объема), прошедшей через определенный трофический уровень за принятый период (рис. 5.7). К основанию пирамиды энергии можно обоснованно добавить снизу еще один прямоугольник, отражающий поступление энергии Солнца.
Пирамида энергий отражает динамику прохождения массы пищи через пищевую (трофическую) цепь, что принципиально отличает ее от пирамид численности и биомасс, отражающих статику системы (количество организмов в данный момент). На форму этой пирамиды не влияют изменения размеров и интенсивности метаболизма особей. Если учтены все источники энергии, то пирамида всегда будет иметь типичный вид (в виде пирамиды вершиной вверх), согласно второму закону термодинамики.
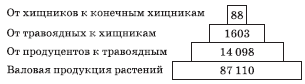 Рис. 5.7. Пирамида энергии: цифры – количество энергии, кДж-м -2r-1
Рис. 5.7. Пирамида энергии: цифры – количество энергии, кДж-м -2r-1
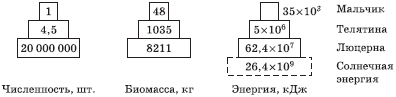 Рис. 5.8. Экологические пирамиды (по Ю. Одуму). Без соблюдения масштаба
Рис. 5.8. Экологические пирамиды (по Ю. Одуму). Без соблюдения масштаба
Пирамиды энергий позволяют не только сравнивать различные биоценозы, но и выявлять относительную значимость популяций в пределах одного сообщества. Они являются наиболее полезными из трех типов экологических пирамид, однако получить данные для их построения труднее всего.
Одним из наиболее удачных и наглядных примеров классических экологических пирамид служат пирамиды, изображенные на рис. 5.8. Они иллюстрируют условный биоценоз, предложенный американским экологом Ю. Одумом. «Биоценоз» состоит из мальчика, питающегося только телятиной, и телят, которые едят исключительно люцерну.
Пирамида энергий отражает динамику прохождения массы пищи через пищевую (трофическую) цепь, что принципиально отличает ее от пирамид численности и биомасс, отражающих статику системы (количество организмов в данный момент). На форму этой пирамиды не влияют изменения размеров и интенсивности метаболизма особей. Если учтены все источники энергии, то пирамида всегда будет иметь типичный вид (в виде пирамиды вершиной вверх), согласно второму закону термодинамики.
Пирамиды энергий позволяют не только сравнивать различные биоценозы, но и выявлять относительную значимость популяций в пределах одного сообщества. Они являются наиболее полезными из трех типов экологических пирамид, однако получить данные для их построения труднее всего.
Одним из наиболее удачных и наглядных примеров классических экологических пирамид служат пирамиды, изображенные на рис. 5.8. Они иллюстрируют условный биоценоз, предложенный американским экологом Ю. Одумом. «Биоценоз» состоит из мальчика, питающегося только телятиной, и телят, которые едят исключительно люцерну.
5.1.3. Закономерности трофического оборота в биоценозе
Живые организмы для своего существования должны постоянно пополнять и расходовать энергию. В пищевой (трофической) цепи, сети и экологических пирамидах каждый последующий уровень, условно говоря, поедает предыдущее звено, используя его для построения своего тела. Трофоэнергетические связи сообщества растений и животных в виде упрощенной схемы потоков на примере биоценоза Рыбинского водохранилища приведены на рис. 5.9.
Главный источник энергии для всего живого на Земле – Солнце. Из всего спектра солнечного излучения, достигающего земной поверхности, только около 40 % составляет фотосинтетически активная радиация (ФАР), имеющая длину волны 380–710 нм. Растения в процессе фотосинтеза усваивают лишь небольшую часть ФАР. Ниже приведены доли усваиваемой ФАР (в %) для различных экосистем.
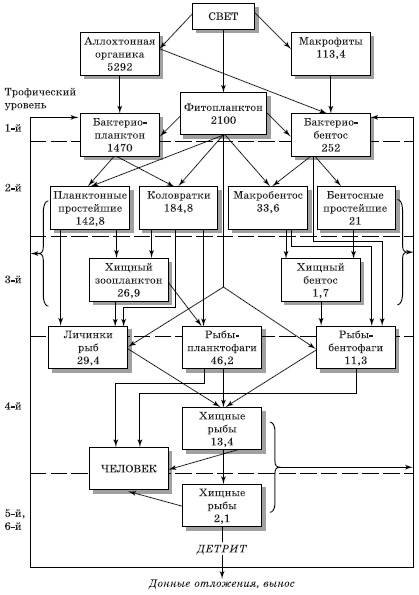 Рис. 5.9. Схема потоков энергии в трофической сети биоценоза (по Н. В. Бутурину, А. Г. Поддубному): цифры – годичная продукция популяций, кДж/м2
Рис. 5.9. Схема потоков энергии в трофической сети биоценоза (по Н. В. Бутурину, А. Г. Поддубному): цифры – годичная продукция популяций, кДж/м2
Океан……………………………………до 1,2
Тропические леса…………………………..до 3,4
Плантации сахарного тростника и кукурузы
(в оптимальных условиях) …………………….. 3—5
Опытные системы с кондиционированными условиями среды по всем показателям (за короткие
периоды времени)…………………………..8—10
В среднем растительность всей планеты…………0,8–1,0
Первичными поставщиками энергии для всех других организмов в цепях питания являются растения. При дальнейших переходах энергии и вещества с одного трофического уровня на другой существуют определенные закономерности.
Главный источник энергии для всего живого на Земле – Солнце. Из всего спектра солнечного излучения, достигающего земной поверхности, только около 40 % составляет фотосинтетически активная радиация (ФАР), имеющая длину волны 380–710 нм. Растения в процессе фотосинтеза усваивают лишь небольшую часть ФАР. Ниже приведены доли усваиваемой ФАР (в %) для различных экосистем.
Океан……………………………………до 1,2
Тропические леса…………………………..до 3,4
Плантации сахарного тростника и кукурузы
(в оптимальных условиях) …………………….. 3—5
Опытные системы с кондиционированными условиями среды по всем показателям (за короткие
периоды времени)…………………………..8—10
В среднем растительность всей планеты…………0,8–1,0
Первичными поставщиками энергии для всех других организмов в цепях питания являются растения. При дальнейших переходах энергии и вещества с одного трофического уровня на другой существуют определенные закономерности.
5.1.3.1. Правило десяти процентов
Р. Линдеман (1942) сформулировал закон пирамиды энергий, или правило 10 %:
Обратный поток, связанный с потреблением веществ и продуцируемым верхним уровнем экологической пирамиды энергии более низкими ее уровнями, например, от животных к растениям, намного слабее – не более 0,5 % (и даже 0,25 %) от общего ее потока, поэтому говорить о круговороте энергии в биоценозе не приходится.
с одного трофического уровня экологической пирамиды переходит на другой, более высокий ее уровень (по «лестнице» продуцент – консумент – редуцент), в среднем около 10 % энергии, поступившей на предыдущий уровень экологической пирамиды.На самом деле потеря бывает либо несколько меньшей, либо несколько большей, но порядок чисел сохраняется.
Обратный поток, связанный с потреблением веществ и продуцируемым верхним уровнем экологической пирамиды энергии более низкими ее уровнями, например, от животных к растениям, намного слабее – не более 0,5 % (и даже 0,25 %) от общего ее потока, поэтому говорить о круговороте энергии в биоценозе не приходится.
5.1.3.2. Правило биологического усиления
Вместе с полезными веществами с одного трофического уровня на другой поступают и «вредные» вещества. Однако если полезное вещество при его излишке легко выводится из организма, то вредное не только плохо выводится, но и накапливается в пищевой цепи. Таков закон природы, называемый правилом накопления токсических веществ (биотического усиления) в пищевой цепи и справедливый для всех биоценозов.
Иначе говоря, если энергия при переходе на более высокий уровень экологической пирамиды десятикратно теряется, то накопление ряда веществ, в том числе токсичных и радиоактивных, примерно в такой же пропорции увеличивается, что впервые было обнаружено в 50-х годах на одном из заводов комиссией по атомной энергии в штате Вашингтон. Явление биотического накопления нагляднее всего демонстрируют устойчивые радионуклиды и пестициды. В водных биоценозах накопление многих токсичных веществ, в том числе хлорорганических пестицидов, коррелируется с массой жиров (липидов), т. е. явно имеет энергетическую подоснову.
В середине 60-х годов появилось, казалось бы, неожиданное сообщение о том, что пестицид дихлордифенилтрихлорэтан (ДДТ) обнаружен в печени пингвинов в Антарктиде – месте, чрезвычайно удаленном от районов его возможного применения. От отравления ДДТ сильно страдают конечные хищники, особенно птицы, так на востоке США полностью исчез сапсан. Птицы оказались наиболее уязвимы в связи с вызываемыми ДДТ гормональными изменениями, влияющими на обмен кальция. Это приводит к утончению скорлупы яиц, и они чаще разбиваются.
Биотическое накопление происходит очень стремительно, например, в случае с пестицидом ДДТ, попавшим в воду болот при многолетнем их опылении с целью сокращения численности нежелательных человеку насекомых на Лонг-Айленде. Для данного случая содержание ДДТ в ррт[27] (по Ю. Одуму) приведено ниже для следующих объектов:
вода…………………………………0,00005
планктон ……………………………….. 0,04
планктоноядные организмы………………….0,23
щука (хищная рыба)………………………..1,33
рыба-игла (хищная рыба)…………………….2,07
цапля (питается мелкими животными)………… 3,57
крачка (питается мелкими животными)………… 3,91
серебристая чайка (падальщик)………………..6,00
крохаль (птица, питается мелкой рыбой)……….. 22,8
баклан (питается крупной рыбой) ……………… 26,4
Специалисты по борьбе с насекомыми «благоразумно» не применяли такие концентрации, которые могли бы быть непосредственно летальны для рыб и других животных. Тем не менее со временем было установлено, что в тканях рыбоядных животных концентрация ДДТ почти в 500 тыс. раз выше, чем в воде. В среднем, как и в приведенном примере, концентрация вредного вещества в каждом последующем звене экологической пирамиды примерно в 10 раз выше, чем в предыдущем.
Принцип биотического усиления (накопления) должен быть принят во внимание при любых решениях, связанных с поступлением соответствующих загрязнений в природную среду. Следует учитывать, что скорость изменения концентрации может увеличиваться или уменьшаться под действием некоторых факторов. Так, человек получит меньше ДДТ, чем птица, питающаяся рыбой. Это частично объясняется удалением пестицидов при обработке и варке рыбы. Кроме того, рыба находится в более опасном положении, ибо получает ДДТ не только через пищу, но и непосредственно из воды.
Иначе говоря, если энергия при переходе на более высокий уровень экологической пирамиды десятикратно теряется, то накопление ряда веществ, в том числе токсичных и радиоактивных, примерно в такой же пропорции увеличивается, что впервые было обнаружено в 50-х годах на одном из заводов комиссией по атомной энергии в штате Вашингтон. Явление биотического накопления нагляднее всего демонстрируют устойчивые радионуклиды и пестициды. В водных биоценозах накопление многих токсичных веществ, в том числе хлорорганических пестицидов, коррелируется с массой жиров (липидов), т. е. явно имеет энергетическую подоснову.
В середине 60-х годов появилось, казалось бы, неожиданное сообщение о том, что пестицид дихлордифенилтрихлорэтан (ДДТ) обнаружен в печени пингвинов в Антарктиде – месте, чрезвычайно удаленном от районов его возможного применения. От отравления ДДТ сильно страдают конечные хищники, особенно птицы, так на востоке США полностью исчез сапсан. Птицы оказались наиболее уязвимы в связи с вызываемыми ДДТ гормональными изменениями, влияющими на обмен кальция. Это приводит к утончению скорлупы яиц, и они чаще разбиваются.
Биотическое накопление происходит очень стремительно, например, в случае с пестицидом ДДТ, попавшим в воду болот при многолетнем их опылении с целью сокращения численности нежелательных человеку насекомых на Лонг-Айленде. Для данного случая содержание ДДТ в ррт[27] (по Ю. Одуму) приведено ниже для следующих объектов:
вода…………………………………0,00005
планктон ……………………………….. 0,04
планктоноядные организмы………………….0,23
щука (хищная рыба)………………………..1,33
рыба-игла (хищная рыба)…………………….2,07
цапля (питается мелкими животными)………… 3,57
крачка (питается мелкими животными)………… 3,91
серебристая чайка (падальщик)………………..6,00
крохаль (птица, питается мелкой рыбой)……….. 22,8
баклан (питается крупной рыбой) ……………… 26,4
Специалисты по борьбе с насекомыми «благоразумно» не применяли такие концентрации, которые могли бы быть непосредственно летальны для рыб и других животных. Тем не менее со временем было установлено, что в тканях рыбоядных животных концентрация ДДТ почти в 500 тыс. раз выше, чем в воде. В среднем, как и в приведенном примере, концентрация вредного вещества в каждом последующем звене экологической пирамиды примерно в 10 раз выше, чем в предыдущем.
Принцип биотического усиления (накопления) должен быть принят во внимание при любых решениях, связанных с поступлением соответствующих загрязнений в природную среду. Следует учитывать, что скорость изменения концентрации может увеличиваться или уменьшаться под действием некоторых факторов. Так, человек получит меньше ДДТ, чем птица, питающаяся рыбой. Это частично объясняется удалением пестицидов при обработке и варке рыбы. Кроме того, рыба находится в более опасном положении, ибо получает ДДТ не только через пищу, но и непосредственно из воды.
5.2. Видовая структура биоценозов
Видовая структура – это количество видов, образующих биоценоз, и соотношение их численностей. Точные сведения о числе видов, входящих в тот или иной биоценоз, получить чрезвычайно трудно из-за микроорганизмов, практически не поддающихся учету.
Видовой состав и насыщенность биоценоза зависят от условий среды. На Земле существуют как резко обедненные сообщества полярных пустынь, так и богатейшие сообщества тропических лесов, коралловых рифов и т. п. Самыми богатыми по видовому разнообразию являются биоценозы влажных тропических лесов, в которых одних растений фитоценоза насчитываются сотни видов.
Виды, преобладающие по численности, массе и развитию, называют доминантными (от лат. dominantis – господствующий). Однако среди них выделяют эдификаторы (от лат. edifikator – строитель) – виды, которые своей жизнедеятельностью в наибольшей степени формируют среду обитания, предопределяя существование других организмов. Именно они порождают спектр разнообразия в биоценозе. Так, в еловом лесу доминирует ель, в смешанном – ель, береза и осина, в степи – ковыль и типчак. При этом ель в еловом лесу наряду с доминантностью обладает сильными эдификаторными свойствами, выражающимися в способности затенять почву, создавать кислую среду своими корнями и образовывать специфические подзолистые почвы. Вследствие этого под пологом ели могут жить только тенелюбивые растения. Одновременно с этим в нижнем ярусе елового леса доминантой может быть, например, черника, но эдификатором она не является.
Предворяя обсуждение видовой структуры биоценоза, следует обратить внимание на принцип Л. Г. Раменского (1924) – Г. А. Глизона (1926) или принцип континуума:[28]
Видовой состав и насыщенность биоценоза зависят от условий среды. На Земле существуют как резко обедненные сообщества полярных пустынь, так и богатейшие сообщества тропических лесов, коралловых рифов и т. п. Самыми богатыми по видовому разнообразию являются биоценозы влажных тропических лесов, в которых одних растений фитоценоза насчитываются сотни видов.
Виды, преобладающие по численности, массе и развитию, называют доминантными (от лат. dominantis – господствующий). Однако среди них выделяют эдификаторы (от лат. edifikator – строитель) – виды, которые своей жизнедеятельностью в наибольшей степени формируют среду обитания, предопределяя существование других организмов. Именно они порождают спектр разнообразия в биоценозе. Так, в еловом лесу доминирует ель, в смешанном – ель, береза и осина, в степи – ковыль и типчак. При этом ель в еловом лесу наряду с доминантностью обладает сильными эдификаторными свойствами, выражающимися в способности затенять почву, создавать кислую среду своими корнями и образовывать специфические подзолистые почвы. Вследствие этого под пологом ели могут жить только тенелюбивые растения. Одновременно с этим в нижнем ярусе елового леса доминантой может быть, например, черника, но эдификатором она не является.
Предворяя обсуждение видовой структуры биоценоза, следует обратить внимание на принцип Л. Г. Раменского (1924) – Г. А. Глизона (1926) или принцип континуума:[28]
широкое перекрытие экологических амплитуд и рассредоточенность центров распределения популяций вдоль градиента среды приводят к плавному переходу одного сообщества в другое, поэтому, как правило, не образуют строго фиксированные сообщества.Принципу континуума Н. Ф. Реймерс противопоставляет принцип биоценотической прерывности:
виды формируют экологически определенные системные совокупности – сообщества и биоценозы, отличающиеся от соседних, хотя и сравнительно постепенно в них переходящие.
5.2.1. Взаимоотношения между организмами
При изучении взаимоотношений между организмами прежде всего выделяют:
• внутривидовые взаимодействия между особями одного вида;
• межвидовую конкуренцию на данном трофическом уровне за необходимые ресурсы, например, пищу, свет и пространство;
• межвидовые взаимодействия популяций, находящихся на разных трофических уровнях, например, отношения типа «хищник – жертва» и «хозяин – паразит»;
• иные типы, более сложные, например симбиотические.
• внутривидовые взаимодействия между особями одного вида;
• межвидовую конкуренцию на данном трофическом уровне за необходимые ресурсы, например, пищу, свет и пространство;
• межвидовые взаимодействия популяций, находящихся на разных трофических уровнях, например, отношения типа «хищник – жертва» и «хозяин – паразит»;
• иные типы, более сложные, например симбиотические.
5.2.1.1. Конкуренция
Конкуренция имеет место в тех случаях, когда взаимодействие между двумя или несколькими особями или популяциями неблагоприятно влияет на рост, выживаемость, приспособленность каждой особи и (или) величину каждой популяции. В основном это происходит при недостатке какого-либо необходимого им всем ресурса. Конкуренция может быть между особями одного вида (внутривидовая) или разных видов (межвидовая), причем и та и другая важны для сообщества. Считают, что конкуренция, особенно межвидовая, – главный механизм возникновения биоразнообразия.
Каждой популяции выгодно использовать любую возможность для защиты от конкуренции с другими видами. Естественный отбор помогает особям, занимающим недоступные для других участки в пространстве экологических ниш, и тем самым ведет к уменьшению перекрывания в потреблении ресурсов и увеличению разнообразия ниш. Таким образом, конкуренция влияет на величину реализованной ниши, которая в свою очередь является фактором, влияющим на видовое богатство биоценоза.
Внутривидовая конкуренция. Имеющиеся ресурсы потребляются особями вида неодинаково (рис. 5.10, а). Те особи, которые используют данный ресурс в краевых, но зато менее оспариваемых местах его градиента, обладают более высокой индивидуальной приспособленностью, чем особи, потребляющие ресурс в зоне его оптимума, где конкуренция особенно велика.
В период роста популяции вторые особи используют оптимальные ресурсы. С увеличением ее плотности преимущества первых уменьшаются из-за внутривидовой конкуренции. Одновременно складываются благоприятные условия для «отклоняющихся» особей, использующих менее оспариваемый ресурс не в оптимальной зоне. Таким образом, увеличивается разнообразие ресурсов и местообитаний, освоенных данной популяцией в целом. Следовательно, внутривидовая конкуренция способствует расширению ниши и приближению реализованной ниши к фундаментальной (см. разд. 5.4). Однако понижение доступности самих ресурсов вызывает прямо противоположную реакцию.
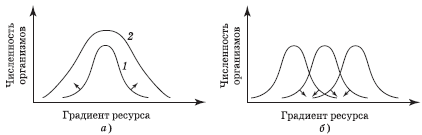 Рис. 5.10. Изменения ширины ниши при внутривидовой (а) и межвидовой (б) конкуренциях (по П. Джиллеру): 1 – низкая плотность популяции; 2 – высокая плотность популяции. Стрелки – направление изменений
Рис. 5.10. Изменения ширины ниши при внутривидовой (а) и межвидовой (б) конкуренциях (по П. Джиллеру): 1 – низкая плотность популяции; 2 – высокая плотность популяции. Стрелки – направление изменений
Межвидовая конкуренция. Особи некоторого вида, потребляющие краевые ресурсы, не могут использовать их так же эффективно, как представители иных видов, для которых эти ресурсы оптимальны. Поэтому область перекрытия между нишами уменьшается так, что по мере специализации ниши становятся уже. В результате размеры популяции одного или нескольких конкурирующих видов также сокращаются (рис. 5.10, б). Конкуренция неблагоприятно воздействует на все виды, использующие один и тот же ограниченный ресурс, причем в одно и то же время и в одном и том же месте, что потенциально вызывает конкурентное исключение некоторых видов по принципу Г. Ф. Гаузе (рис. 5.11).
При совместном выращивании двух видов инфузорий в единой питательной среде вид 1 оказывается более конкурентоспособным в захвате пищи, чем вид 2. Через 5–6 сут численность вида 2 начинает уменьшаться, и спустя примерно 20 сут этот вид почти полностью исчезает, т. е. происходит его конкурентное исключение. Вид 1 достигает стационарной фазы роста позднее, чем при выращивании в отдельной культуре. Хотя этот вид и более конкурентоспособен, но на него также отрицательно влияет конкуренция.
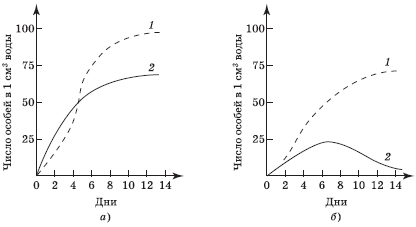 Рис. 5.11. Рост численности двух видов инфузорий в одной культуре (в опытах Г. Гаузе) (по Ф.Дрэ): а – при выращивании видов отдельно; б – при совместном выращивании в общей среде
Рис. 5.11. Рост численности двух видов инфузорий в одной культуре (в опытах Г. Гаузе) (по Ф.Дрэ): а – при выращивании видов отдельно; б – при совместном выращивании в общей среде
В природных условиях менее конкурентоспособный вид редко исчезает полностью – просто его численность сильно уменьшается, однако иногда она может снова возрасти, прежде чем установится равновесное состояние. Принцип конкурентного исключения Г. Ф. Гаузе был в дальнейшем неоднократно подтвержден на животных. Таким образом, при возрастании видового разнообразия в результате межвидовой конкуренции имеет место большее разделение ниш и реализованные ниши взаимодействующих видов пропорционально сокращаются. При большой сходности видов происходит их конкурентное исключение.
Каждой популяции выгодно использовать любую возможность для защиты от конкуренции с другими видами. Естественный отбор помогает особям, занимающим недоступные для других участки в пространстве экологических ниш, и тем самым ведет к уменьшению перекрывания в потреблении ресурсов и увеличению разнообразия ниш. Таким образом, конкуренция влияет на величину реализованной ниши, которая в свою очередь является фактором, влияющим на видовое богатство биоценоза.
Внутривидовая конкуренция. Имеющиеся ресурсы потребляются особями вида неодинаково (рис. 5.10, а). Те особи, которые используют данный ресурс в краевых, но зато менее оспариваемых местах его градиента, обладают более высокой индивидуальной приспособленностью, чем особи, потребляющие ресурс в зоне его оптимума, где конкуренция особенно велика.
В период роста популяции вторые особи используют оптимальные ресурсы. С увеличением ее плотности преимущества первых уменьшаются из-за внутривидовой конкуренции. Одновременно складываются благоприятные условия для «отклоняющихся» особей, использующих менее оспариваемый ресурс не в оптимальной зоне. Таким образом, увеличивается разнообразие ресурсов и местообитаний, освоенных данной популяцией в целом. Следовательно, внутривидовая конкуренция способствует расширению ниши и приближению реализованной ниши к фундаментальной (см. разд. 5.4). Однако понижение доступности самих ресурсов вызывает прямо противоположную реакцию.
Межвидовая конкуренция. Особи некоторого вида, потребляющие краевые ресурсы, не могут использовать их так же эффективно, как представители иных видов, для которых эти ресурсы оптимальны. Поэтому область перекрытия между нишами уменьшается так, что по мере специализации ниши становятся уже. В результате размеры популяции одного или нескольких конкурирующих видов также сокращаются (рис. 5.10, б). Конкуренция неблагоприятно воздействует на все виды, использующие один и тот же ограниченный ресурс, причем в одно и то же время и в одном и том же месте, что потенциально вызывает конкурентное исключение некоторых видов по принципу Г. Ф. Гаузе (рис. 5.11).
При совместном выращивании двух видов инфузорий в единой питательной среде вид 1 оказывается более конкурентоспособным в захвате пищи, чем вид 2. Через 5–6 сут численность вида 2 начинает уменьшаться, и спустя примерно 20 сут этот вид почти полностью исчезает, т. е. происходит его конкурентное исключение. Вид 1 достигает стационарной фазы роста позднее, чем при выращивании в отдельной культуре. Хотя этот вид и более конкурентоспособен, но на него также отрицательно влияет конкуренция.
В природных условиях менее конкурентоспособный вид редко исчезает полностью – просто его численность сильно уменьшается, однако иногда она может снова возрасти, прежде чем установится равновесное состояние. Принцип конкурентного исключения Г. Ф. Гаузе был в дальнейшем неоднократно подтвержден на животных. Таким образом, при возрастании видового разнообразия в результате межвидовой конкуренции имеет место большее разделение ниш и реализованные ниши взаимодействующих видов пропорционально сокращаются. При большой сходности видов происходит их конкурентное исключение.
5.2.1.2. Хищничество
Во многих существующих природных сообществах отмечается сильное перекрывание ниш потребления ресурсов, тем не менее не приводящее к конкурентному исключению видов, описанному ранее. Причиной этого может быть либо неограниченность ресурса (например, в наземных биоценозах никто не испытывает недостатка в кислороде), либо наличие какого-нибудь внешнего фактора, удерживающего численность потенциально конкурирующих популяций сосуществующих видов ниже уровня, допускаемого емкостью среды.
Важным механизмом создания структуры сообщества, альтернативным механизму разделения ресурсов путем конкуренции, является хищничество. Так, при значительной смертности в результате хищничества в популяции самого конкурентоспособного или многочисленного вида конкурентное исключение иных видов будет остановлено на неопределенно долгое время. При этом возможно более сильное перекрывание ниш и, следовательно, локальное увеличение видового разнообразия.
Хищничество – трудный и требующий большой затраты времени процесс. При активной охоте хищники нередко подвергаются опасностям не меньше, чем их жертвы. Многие хищники сами погибают в процессе межвидовой борьбы за добычу, а также от голода. Известны случаи гибели львиц в ходе столкновения со слонами или кабанами. Только самые быстрые и сильные хищники способны затрачивать необходимое время в поисках добычи, преследовать жертву на большом расстоянии. Менее энергичные обречены на голодную смерть.
Хищничество воздействует на динамику и пространственное распределение популяции жертвы, что в свою очередь влияет на структуру и функции сообщества (биоценоза) вплоть до катастрофического их изменения. При этом в наземных системах полное уничтожение растений происходит редко и в основном бывает не избирательным (например, налет саранчи).
Большая часть фактов, подтверждающих теорию о роли хищничества, связана с взаимодействиями на трофическом уровне. Воздействия выпаса на урожай надземных частей растений не предсказуемы, но он может изменить конкурентное равновесие между поедаемым растением и другими видами. Выпас ведет также к уменьшению числа семян.
Поедание семян и плодов некоторыми первичными консументами ведет к изменениям или регулированию видового состава растительных сообществ. Эксперименты, в которых проводилось искусственное удаление отдельных видов, показали, что поедание семян муравьями или грызунами увеличивает видовое разнообразие в биоценозе.
Хищничество не всегда вызывает повышение разнообразия на нижних трофических уровнях. Хотя хищники могут понизить плотность популяции жертвы, это не обязательно уменьшает потребление ресурса – условие, необходимое для повышения видового разнообразия. В некоторых случаях ослабление внутривидовой конкуренции может активизировать вид и его размножение, что в свою очередь увеличит использование ресурса. Хищничество на одном трофическом уровне может привести к «каскадному» эффекту на других уровнях и вызвать снижение разнообразия в биоценозе в целом.
Важным механизмом создания структуры сообщества, альтернативным механизму разделения ресурсов путем конкуренции, является хищничество. Так, при значительной смертности в результате хищничества в популяции самого конкурентоспособного или многочисленного вида конкурентное исключение иных видов будет остановлено на неопределенно долгое время. При этом возможно более сильное перекрывание ниш и, следовательно, локальное увеличение видового разнообразия.
Хищничество – трудный и требующий большой затраты времени процесс. При активной охоте хищники нередко подвергаются опасностям не меньше, чем их жертвы. Многие хищники сами погибают в процессе межвидовой борьбы за добычу, а также от голода. Известны случаи гибели львиц в ходе столкновения со слонами или кабанами. Только самые быстрые и сильные хищники способны затрачивать необходимое время в поисках добычи, преследовать жертву на большом расстоянии. Менее энергичные обречены на голодную смерть.
Хищничество воздействует на динамику и пространственное распределение популяции жертвы, что в свою очередь влияет на структуру и функции сообщества (биоценоза) вплоть до катастрофического их изменения. При этом в наземных системах полное уничтожение растений происходит редко и в основном бывает не избирательным (например, налет саранчи).
Большая часть фактов, подтверждающих теорию о роли хищничества, связана с взаимодействиями на трофическом уровне. Воздействия выпаса на урожай надземных частей растений не предсказуемы, но он может изменить конкурентное равновесие между поедаемым растением и другими видами. Выпас ведет также к уменьшению числа семян.
Поедание семян и плодов некоторыми первичными консументами ведет к изменениям или регулированию видового состава растительных сообществ. Эксперименты, в которых проводилось искусственное удаление отдельных видов, показали, что поедание семян муравьями или грызунами увеличивает видовое разнообразие в биоценозе.
Хищничество не всегда вызывает повышение разнообразия на нижних трофических уровнях. Хотя хищники могут понизить плотность популяции жертвы, это не обязательно уменьшает потребление ресурса – условие, необходимое для повышения видового разнообразия. В некоторых случаях ослабление внутривидовой конкуренции может активизировать вид и его размножение, что в свою очередь увеличит использование ресурса. Хищничество на одном трофическом уровне может привести к «каскадному» эффекту на других уровнях и вызвать снижение разнообразия в биоценозе в целом.
5.2.1.3. Сопряженные колебания численности хищника и жертвы
Как правило, хищник не может полностью истребить жертву. В большинстве случаев наблюдают сопряженные (согласованные между собой) колебания численности обеих популяций. В одном из наиболее известных и повторяемых в литературе примеров описывается цикл колебаний численности зайца-беляка и рыси (рис. 5.12). При этом основным является вопрос, кто чью численность контролирует, хищник ли жертву, или наоборот.
Достоверно установлено, что популяции зайцев достигают пика численности через каждые 9 лет; вслед за этим численности популяций рыси также достигают пика. Однако затем численность популяций зайца резко сокращается. Первоначально эту закономерность объясняли тем, что рыси в определенный момент съедают слишком много пищи (зайцев), превышая поддерживающую емкость среды, что ведет к сокращению численности самой рыси, и весь цикл повторяется заново.
Позже в регионах, где рысь истреблена, была обнаружена точно такая же цикличность изменения численности зайцев. Таким образом, было установлено, что численность зайцев (пищевой ресурс) контролирует численность рыси, а не наоборот.
На основании изложенного можно сделать вывод, что главным механизмом, создающим структуру сообществ и биоценозов, служит конкуренция, а хищничество лишь регулирует видовое богатство в отдельных случаях. При этом, как следует из рис. 5.12, изменение численности хищника отстает от колебаний популяции жертвы, что относится в первую очередь к специализированным хищникам, которые не могут переключиться на другие виды пищи при снижении численности основного кормового вида (или переключаются в малой степени и с опазданием). И, напротив, обилие альтернативных кормов для хищника даже стабилизирует численность жертв. Вероятно, именно поэтому резкие вспышки численности не характерны для сложных биоценозов, например тропических лесов.
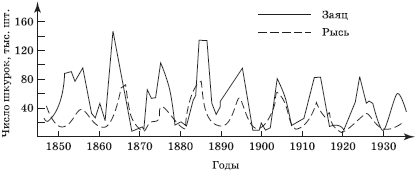 Рис. 5.12. Равновесие популяций в системе «хищник—жертва» на примере «рысь—заяц» по результатам наблюдений за количеством шкурок, добытых компанией Гудзонова залива (по Мак-Луличу)
Рис. 5.12. Равновесие популяций в системе «хищник—жертва» на примере «рысь—заяц» по результатам наблюдений за количеством шкурок, добытых компанией Гудзонова залива (по Мак-Луличу)
Хотя крупные хищники вроде рыси чаще привлекают внимание исследователей, значительно более важными и многочисленными в этом отношении являются разнообразные паразиты. Все виды растений, животных, грибов и даже микробов могут быть поражены паразитами, роль которых сходна с ролью крупных хищников. С ростом численности популяции организма-хозяина паразиты и их переносчики (например, кровососущие комары) легко находят себе новые жертвы, что увеличивает смертность хозяев. При малой плотности последних распространение паразитов затруднено, число пораженных особей падает и численность вида-хозяина восстанавливается.
Паразиты не обязательно убивают хозяина, но обычно ослабляют его, делая более чувствительным к влиянию неблагоприятных условий и (или) хищничества. В большинстве случаев животные, пойманные хищниками, сильно поражены паразитами, а добытые охотниками – здоровы. В пищевой сети каждый организм обычно зависит от нескольких паразитов и хищников, т. е. речь обычно идет о равновесии между организмом и его естественными врагами.
Равновесие в системах хищник—жертва и паразит—хозяин не возникает автоматически, а устанавливается на протяжении длительного времени (много тысяч и даже миллионов лет). За такое время виды, взаимодействующие подобным образом, адаптируются друг к другу и к среде обитания так, что естественные враги не уничтожают полностью популяцию жертвы.
Важность эволюционного приспособления становится яснее при его отсутствии. При встрече организмов, развивавшихся в изоляции друг от друга, например на разных материках, их популяции не способны сосуществовать в равновесии. Так, интродуцированный (завезенный, переселенный) вид не всегда сталкивается на новом месте с естественными врагами, способными контролировать его численность. В результате численность привезенной популяции стремительно вырастает, нанося исключительный ущерб местному биоценозу, включая вымирание отдельных видов. Классическими примерами таких случаев являются интродукция кроликов[29] в Австралию и занос грибка, вызывающего рак коры каштана, в Северную Америку.
Достоверно установлено, что популяции зайцев достигают пика численности через каждые 9 лет; вслед за этим численности популяций рыси также достигают пика. Однако затем численность популяций зайца резко сокращается. Первоначально эту закономерность объясняли тем, что рыси в определенный момент съедают слишком много пищи (зайцев), превышая поддерживающую емкость среды, что ведет к сокращению численности самой рыси, и весь цикл повторяется заново.
Позже в регионах, где рысь истреблена, была обнаружена точно такая же цикличность изменения численности зайцев. Таким образом, было установлено, что численность зайцев (пищевой ресурс) контролирует численность рыси, а не наоборот.
На основании изложенного можно сделать вывод, что главным механизмом, создающим структуру сообществ и биоценозов, служит конкуренция, а хищничество лишь регулирует видовое богатство в отдельных случаях. При этом, как следует из рис. 5.12, изменение численности хищника отстает от колебаний популяции жертвы, что относится в первую очередь к специализированным хищникам, которые не могут переключиться на другие виды пищи при снижении численности основного кормового вида (или переключаются в малой степени и с опазданием). И, напротив, обилие альтернативных кормов для хищника даже стабилизирует численность жертв. Вероятно, именно поэтому резкие вспышки численности не характерны для сложных биоценозов, например тропических лесов.
Хотя крупные хищники вроде рыси чаще привлекают внимание исследователей, значительно более важными и многочисленными в этом отношении являются разнообразные паразиты. Все виды растений, животных, грибов и даже микробов могут быть поражены паразитами, роль которых сходна с ролью крупных хищников. С ростом численности популяции организма-хозяина паразиты и их переносчики (например, кровососущие комары) легко находят себе новые жертвы, что увеличивает смертность хозяев. При малой плотности последних распространение паразитов затруднено, число пораженных особей падает и численность вида-хозяина восстанавливается.
Паразиты не обязательно убивают хозяина, но обычно ослабляют его, делая более чувствительным к влиянию неблагоприятных условий и (или) хищничества. В большинстве случаев животные, пойманные хищниками, сильно поражены паразитами, а добытые охотниками – здоровы. В пищевой сети каждый организм обычно зависит от нескольких паразитов и хищников, т. е. речь обычно идет о равновесии между организмом и его естественными врагами.
Равновесие в системах хищник—жертва и паразит—хозяин не возникает автоматически, а устанавливается на протяжении длительного времени (много тысяч и даже миллионов лет). За такое время виды, взаимодействующие подобным образом, адаптируются друг к другу и к среде обитания так, что естественные враги не уничтожают полностью популяцию жертвы.
Важность эволюционного приспособления становится яснее при его отсутствии. При встрече организмов, развивавшихся в изоляции друг от друга, например на разных материках, их популяции не способны сосуществовать в равновесии. Так, интродуцированный (завезенный, переселенный) вид не всегда сталкивается на новом месте с естественными врагами, способными контролировать его численность. В результате численность привезенной популяции стремительно вырастает, нанося исключительный ущерб местному биоценозу, включая вымирание отдельных видов. Классическими примерами таких случаев являются интродукция кроликов[29] в Австралию и занос грибка, вызывающего рак коры каштана, в Северную Америку.
5.2.2. Влияние абиотических факторов среды
Поскольку ни конкуренция, ни хищничество в полной мере не объясняют все известные в живой природе случаи формирования видовой структуры биоценозов, учеными были предприняты попытки найти какой-либо другой механизм, обобщающий все варианты. Важным условием является степень суровости (или, напротив, благоприятности) физической среды обитания, т. е. совокупности абиотических факторов.
Установлено, что в очень суровых условиях среды численность популяций падает ниже тех уровней, при которых они конкурируют. На основании этого вывода и учитывая, что при наиболее благоприятных абиотических факторах плотность популяции уменьшается под действием хищников, Дж. Коннелл предложил схему, приведенную на рис. 5.13. В соответствии с ней в мягких условиях тропиков главное – устоять против растительноядных организмов, а с увеличением широты основным становится противодействие конкуренции.
Принцип действия законов минимума Ю. Либиха в масштабах сообществ и биоценозов установил А. Тинеман (1926) как закон действия факторов:
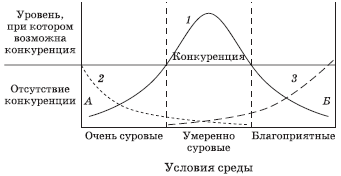 Рис. 5.13. Схема взаимодействия механизмов организации биоценоза (по Дж. Коннеллу): 1 – численность популяции; 2 – смертность, вызванная неблагоприятными абиотическими факторами среды обитания; 3 – смертность, обусловленная хищничеством; А – популяции, численность которых ограничивается неблагоприятными физическими факторами среды; Б – популяции, численность которых ограничивается интенсивным хищничеством
Рис. 5.13. Схема взаимодействия механизмов организации биоценоза (по Дж. Коннеллу): 1 – численность популяции; 2 – смертность, вызванная неблагоприятными абиотическими факторами среды обитания; 3 – смертность, обусловленная хищничеством; А – популяции, численность которых ограничивается неблагоприятными физическими факторами среды; Б – популяции, численность которых ограничивается интенсивным хищничеством
Установлено, что в очень суровых условиях среды численность популяций падает ниже тех уровней, при которых они конкурируют. На основании этого вывода и учитывая, что при наиболее благоприятных абиотических факторах плотность популяции уменьшается под действием хищников, Дж. Коннелл предложил схему, приведенную на рис. 5.13. В соответствии с ней в мягких условиях тропиков главное – устоять против растительноядных организмов, а с увеличением широты основным становится противодействие конкуренции.
Принцип действия законов минимума Ю. Либиха в масштабах сообществ и биоценозов установил А. Тинеман (1926) как закон действия факторов: