Страница:
Последнее обстоятельство заставляет вновь вернуться к цитированному нами учебнику, повествующему школьникам о карбоновом периоде «с его теплым влажным климатом и воздухом, богатым углекислым газом вследствие сильной вулканической деятельности». А как мы теперь помним – из балансовых моделей – захоронение больших количеств неокисленной органики (что и происходило в процессе углеобразования) по идее должно было вести как раз к уменьшению доли атмосферной углекислоты и, соответственно, к возрастанию содержания кислорода. Это предположение о составе тогдашней атмосферы может быть подкреплено с довольно неожиданной стороны.
На школьных олимпиадах по биологии, проводимых биофаком МГУ, есть «золотой фонд» из вопросов, ответа на которые (если по-честному) не знает никто. В их числе есть и такой: «Почему в карбоновом периоде насекомые достигали размеров, максимальных за всю историю класса (например, стрекоза Meganeura с размахом крыльев до 1 м)?» Стандартный ответ – «Потому что в это время еще не существовало конкурентов и хищников в лице летающих позвоночных – не только птиц и летучих мышей, но и крылатых ящеров,» – вряд ли можно счесть достаточным: гигантизм насекомых был характерен только для карбона, и существа эти вымерли, когда до появления первых птерозавров оставалось еще примерно сто миллионов (!) лет.
Одно из наиболее убедительных объяснений таково. Главным фактором, ограничивающим размер тела насекомого (самые крупные из них – тропические жуки-голиафы размером чуть больше куриного яйца), считают особенности их дыхательной системы. «Кровь» насекомых (гемолимфа) лишена дыхательных пигментов и не участвует в переносе кислорода; дыхание же осуществляется при помощи трахей – ветвящихся трубочек, непосредственно соединяющих клетки внутренних органов с воздушной средой. Воздух внутри трахейной трубки неподвижен – принудительной вентиляции, как в различных типах легочных мешков, там нет, и приток кислорода внутрь тела (так же, как отток углекислого газа) происходит за счет диффузии при разнице парциальных давлений этих газов на внутреннем и внешнем концах трубки. Такой механизм подачи кислорода жестко ограничивает длину трахейной трубки, максимальная протяженность которой (l) достаточно просто вычисляема «из физики»; поэтому максимальный размер тела самого насекомого не может превышать (в сечении) величины 2l , что и составляет ныне то самое «куриное яйцо» – размер жука-голиафа [47]. Поэтому существование в карбоне таких гигантов, как стрекоза-меганевра или основная ее добыча – диктионевриды размером с голубя (рисунок 36), заставляет предположить, что в те времена парциальное давление кислорода в земной атмосфере было заметно выше, чем ныне – как оно и должно быть при захоронении больших количеств неокисленного углерода. Кстати, видимо именно это увеличение соотношения O2/CO2 в атмосфере планеты вызвало «обратный парниковый эффект», приведший к Гондванскому оледенению (как уже однажды случалось в докембрии).

РИСУНОК 36. Палеозойские насекомые и гипотетические предки крылатых насекомых: (а) – стрекоза подотряда Meganeurina, (б) – диктионеврида, (в) – мисхоптерида, (г) – гипотетический предок, родственный современным щетинохвосткам.
Необходимо заметить, что появление в конце раннего карбона (намюр) крылатых насекомых вообще было одним из ключевых событий в истории жизни на суше. Собственно говоря, вся история наземных экосистем – это фактически история трех групп живых организмов: сосудистых растений, насекомых и позвоночных-тетрапод (о них речь пойдет чуть позже – в главе 10), а также их взаимодействия между собой. Оговорка насчет «крылатых» насекомых не случайна: вообще-то первые насекомые (архаичные формы из подкласса первичнобескрылых – Apterygota) появились в девоне, однако именно с возникновением способности к полету у крылатых насекомых (подкласс Pterygota) эта группа стала самой процветающей в животном царстве (сейчас известно более миллиона видов – много больше, чем всех прочих живых существ, вместе взятых).
Крыло насекомого – в отличие от крыльев всех летающих позвоночных – не является видоизмененной конечностью и представляет собой разросшуюся складку кутикулы грудных сегментов, укрепленную жилками – полыми каналами с утолщенными стенками, по которым осуществляется кровоснабжение крыла, проходят нервы и трахеи. Как предполагает А.П. Расницын (1976), крылатые насекомые произошли от предков, обитавших на деревьях и питавшихся содержимым спорангиев; при питании и нападении хищников (паукообразных) они вынуждены были регулярно прыгать с ветки на ветку или на землю (рисунок 36, г). Очевидное преимущество особей, способных к дальнему и точному прыжку и к контролю за положением тела в воздухе обусловило развитие крыльев. Зачатки крыльев в виде боковых пластинчатых выростов на грудных сегментах возникали именно у крупных насекомых, т.к. при малых размерах увеличение поверхности тела невыгодно (и длина прыжка, и особенно его точность уменьшается из-за влияния ветра). Предположение о том, что первичным типом питания для насекомых была палинофагия (питание пыльцой и спорами) и высасывание семезачатков, полностью подтвердилось, когда недавно был открыт способ прямо проанализировать содержимое пищеварительного тракта ископаемых насекомых: оказалось, что у древнейших насекомых кишечники буквально набиты пыльцой. В захоронениях до 70% семезачатков кордаитов оказываются поврежденны (видимо, диктионевридами).
Первые крылатые насекомые летали не слишком хорошо. Они, подобно современным подёнкам (Ephemeroptera), имели не только личиночные, но и имагинальные линьки (имаго – конечная стадия развития насекомого, способная к полету и размножению). Для имагинальных линек необходимо сохранять внутри крыла, между слоями кутикулы, живую ткань; такое крыло тяжелое и неуклюжее (недаром поденки лишь порхают над самой поверхностью реки, где развиваются их личинки), однако во времена, когда еще не было воздушных хищников, лучшего не требовалось. Подлинную революцию вызвало появление насекомых, отказавшихся от имагинальных линек и превративших крыло в тонкую легкую двуслойную кутикулу – стрекоз. Эти воздушные хищники оказались таким же «абсолютным оружием», как некогда зоопланктон миллиметрового размерного класса, спровоцировавший некогда вендский фитопланктонный кризис (см. главу 6-а).
По мнению В.В. Жерихина (1990), именно стрекозы полностью уничтожили характерную для карбона фауну крупноразмерных открытоживущих палинофагов типа диктионеврид (эта смена была одной из самых резких за всю историю класса), заставив крылатых насекомых специализироваться в двух направлениях. Одним вариантом было совершенствовать свой полет (попутно при этом уменьшаясь в размерах), а также тип метаморфоза – появление полного превращения позволяет спрятать червеобразную личинку и неподвижную куколку внутри субстрата. Эта линия эволюции привела в конечном счете к возникновению в начале мезозоя (триас) наиболее процветающих ныне отрядов – двукрылых и перепончатокрылых, а чуть позже – бабочек. Другой выход – перейти к скрытному образу жизни (например, внутри уже знакомой нам валежникообразной «подстилки» карбоновых лесов), превратив переднюю пару крыльев в крышеобразно складываемые жесткие надкрылья, защищающие от повреждений заднюю пару, которая и выполняет летательные функции; по этому пути пошли гриллоновые насекомые, сохраняющие неполный метаморфоз – тараканы, прямокрылые и их родственники, а из насекомых с полным превращением – жуки, самый большой из современных отрядов (более ста тысяч видов), появившийся в начале перми.
Хотя в карбоне уже существовали стрекозы и поденки, личинки их (в отличие от современных) были существами не водными, а наземными. Освоение насекомыми пресных вод началось лишь во второй половине следующего (пермского) периода, когда появились отряды, имеющие водную личинку – веснянки, ручейники, а также поденки современного типа (личинки же стрекоз перестали быть наземными лишь в мезозое). В этой связи В.В. Жерихин (1980) указывает на следующее обстоятельство. Как известно, смыв различных веществ (прежде всего – микроэлементов) должен сильно обеднять почвы водоразделов. В настоящее время подобное обеднение если не предотвращается, то заметно тормозится прямым выносом вещества из водоемов на сушу животными (например, азот и фосфор, смытые с суши в море, оказываются – по пищевым цепям – в организмах сперва рыб, затем рыбоядных морских птиц, а в конечном счете возвращаются на сушу в форме птичьего помета). В пресных водах эту функцию выполняют прежде всего насекомые, которые имеют водных личинок и имаго, способных к дальнему разлету из водоемов.
Что же касается карбона, когда насекомые с водными личинками, судя по всему, отсутствовали, то в те времена обеднение почв на водоразделах (если там вообще формировались почвы) должно было быть особенно сильным. Не исключено, что формирование в перми, и особенно в триасе, достаточно обильной фауны насекомых с водными личинками стало одним из тех факторов, которые изменили почвенную ситуацию на водоразделах и позволили растительности начать «наступление» на первичные пустыни. Важно подчеркнуть, что этот вынос микроэлементов насекомыми мог сыграть здесь роль первого толчка – дальше система будет развиваться с положительной обратной связью, ибо возникшая на водоразделах растительность сама является мощнейшим регулятором стока. Окончательно эта система могла оформиться в позднем триасе – когда появились двукрылые; именно такие их представители, как, например, комары-звонцы (Chironomidae) совмещают огромную биомассу со способностью к дальнему разлету (в том числе за счет пассивного переноса ветром).
В заключении остановимся на одном существенном отличии экосистем древних (палеозойских и мезозойских) озер от современных. Это полное отсутствие погруженных макрофитов, за исключением харовых водорослей (которые депонируют биогены почти на порядок хуже, чем покрытосеменные). Основными продуцентами там скорее всего были плавающие маты, причем в мезозое получили распространение водорослевые маты на каркасе из высших растений. В триасе это были плевромейи – крупные плавающие плауновидные (что-то вроде сильно увеличенного полушника), по виду напоминающие морской буй, где в роли верхней сигнальной лампочки выступает пучок листьев и спорангии; листья, скорее всего, плавали по поверхности воды, постепенно отгнивая с вершины [48]. На каркасе из этих плауновидных возникали плавающие острова из мелких водяных мхов и печеночников (типа любимой аквариумистами риччии), а снаружи все это обрастало зелеными водорослями и цианобактериями; на этих матах существовала богатая фауна беспозвоночных: в палеозое – сходные с многоножками растительноядные эвтикарциноиды, позже – ракообразные и насекомые.
Поскольку все фотосинтезирующие структуры были сосредоточены на поверхности озера (а дно было к тому же еще и затенено), гиполимнион почти всегда был асфиксным и безжизненным, а озера легко эвтрофицировались. Однако в непосредственной близости от плавающего мата концентрация кислорода в воде могла быть очень высока; возможно, именно этим объясняется присутствие в составе фауны эвтрофных (судя по составу отложений) мезозойских озер целого ряда групп насекомых, являющихся ныне реофилами (обитателями быстрых, богатых кислородом рек и ручьев). Кроме того, в составе этих фаун комбинируются как пресноводные формы, имеющие адаптации к водному образу жизни, так и наземные (которые могли «ходить пешком» по поверхности плавающего мата). Эти имевшие огромную биомассу и богатые животным белком «плавучие острова» могли составлять основу питания для многих позвоночных, включая и динозавров. Окончательно экосистемы такого типа исчезли лишь в кайнозое, когда на дне водоемов возникли заросли погруженных макрофитов из покрытосеменных (типа элодеи), резко увеличившие разнообразие и устойчивость пресноводных экосистем. Это и дало А.Г. Пономаренко основание разделить всю историю континентальных водоемов на два принципиальных этапа: первый – от докембрия до середины палеогена (когда основными продуцентами были маты, сперва прикрепленные, затем – плавающие), а второй – от середины палеогена доныне (когда эту роль стали играть погруженные макрофиты).
10. Эволюция наземных позвоночных (1): поздний палеозой – ранний мезозой. Анамнии и амниоты. Две линии амниот – тероморфная и завроморфная.
На школьных олимпиадах по биологии, проводимых биофаком МГУ, есть «золотой фонд» из вопросов, ответа на которые (если по-честному) не знает никто. В их числе есть и такой: «Почему в карбоновом периоде насекомые достигали размеров, максимальных за всю историю класса (например, стрекоза Meganeura с размахом крыльев до 1 м)?» Стандартный ответ – «Потому что в это время еще не существовало конкурентов и хищников в лице летающих позвоночных – не только птиц и летучих мышей, но и крылатых ящеров,» – вряд ли можно счесть достаточным: гигантизм насекомых был характерен только для карбона, и существа эти вымерли, когда до появления первых птерозавров оставалось еще примерно сто миллионов (!) лет.
Одно из наиболее убедительных объяснений таково. Главным фактором, ограничивающим размер тела насекомого (самые крупные из них – тропические жуки-голиафы размером чуть больше куриного яйца), считают особенности их дыхательной системы. «Кровь» насекомых (гемолимфа) лишена дыхательных пигментов и не участвует в переносе кислорода; дыхание же осуществляется при помощи трахей – ветвящихся трубочек, непосредственно соединяющих клетки внутренних органов с воздушной средой. Воздух внутри трахейной трубки неподвижен – принудительной вентиляции, как в различных типах легочных мешков, там нет, и приток кислорода внутрь тела (так же, как отток углекислого газа) происходит за счет диффузии при разнице парциальных давлений этих газов на внутреннем и внешнем концах трубки. Такой механизм подачи кислорода жестко ограничивает длину трахейной трубки, максимальная протяженность которой (l) достаточно просто вычисляема «из физики»; поэтому максимальный размер тела самого насекомого не может превышать (в сечении) величины 2l , что и составляет ныне то самое «куриное яйцо» – размер жука-голиафа [47]. Поэтому существование в карбоне таких гигантов, как стрекоза-меганевра или основная ее добыча – диктионевриды размером с голубя (рисунок 36), заставляет предположить, что в те времена парциальное давление кислорода в земной атмосфере было заметно выше, чем ныне – как оно и должно быть при захоронении больших количеств неокисленного углерода. Кстати, видимо именно это увеличение соотношения O2/CO2 в атмосфере планеты вызвало «обратный парниковый эффект», приведший к Гондванскому оледенению (как уже однажды случалось в докембрии).
РИСУНОК 36. Палеозойские насекомые и гипотетические предки крылатых насекомых: (а) – стрекоза подотряда Meganeurina, (б) – диктионеврида, (в) – мисхоптерида, (г) – гипотетический предок, родственный современным щетинохвосткам.
Необходимо заметить, что появление в конце раннего карбона (намюр) крылатых насекомых вообще было одним из ключевых событий в истории жизни на суше. Собственно говоря, вся история наземных экосистем – это фактически история трех групп живых организмов: сосудистых растений, насекомых и позвоночных-тетрапод (о них речь пойдет чуть позже – в главе 10), а также их взаимодействия между собой. Оговорка насчет «крылатых» насекомых не случайна: вообще-то первые насекомые (архаичные формы из подкласса первичнобескрылых – Apterygota) появились в девоне, однако именно с возникновением способности к полету у крылатых насекомых (подкласс Pterygota) эта группа стала самой процветающей в животном царстве (сейчас известно более миллиона видов – много больше, чем всех прочих живых существ, вместе взятых).
Крыло насекомого – в отличие от крыльев всех летающих позвоночных – не является видоизмененной конечностью и представляет собой разросшуюся складку кутикулы грудных сегментов, укрепленную жилками – полыми каналами с утолщенными стенками, по которым осуществляется кровоснабжение крыла, проходят нервы и трахеи. Как предполагает А.П. Расницын (1976), крылатые насекомые произошли от предков, обитавших на деревьях и питавшихся содержимым спорангиев; при питании и нападении хищников (паукообразных) они вынуждены были регулярно прыгать с ветки на ветку или на землю (рисунок 36, г). Очевидное преимущество особей, способных к дальнему и точному прыжку и к контролю за положением тела в воздухе обусловило развитие крыльев. Зачатки крыльев в виде боковых пластинчатых выростов на грудных сегментах возникали именно у крупных насекомых, т.к. при малых размерах увеличение поверхности тела невыгодно (и длина прыжка, и особенно его точность уменьшается из-за влияния ветра). Предположение о том, что первичным типом питания для насекомых была палинофагия (питание пыльцой и спорами) и высасывание семезачатков, полностью подтвердилось, когда недавно был открыт способ прямо проанализировать содержимое пищеварительного тракта ископаемых насекомых: оказалось, что у древнейших насекомых кишечники буквально набиты пыльцой. В захоронениях до 70% семезачатков кордаитов оказываются поврежденны (видимо, диктионевридами).
Первые крылатые насекомые летали не слишком хорошо. Они, подобно современным подёнкам (Ephemeroptera), имели не только личиночные, но и имагинальные линьки (имаго – конечная стадия развития насекомого, способная к полету и размножению). Для имагинальных линек необходимо сохранять внутри крыла, между слоями кутикулы, живую ткань; такое крыло тяжелое и неуклюжее (недаром поденки лишь порхают над самой поверхностью реки, где развиваются их личинки), однако во времена, когда еще не было воздушных хищников, лучшего не требовалось. Подлинную революцию вызвало появление насекомых, отказавшихся от имагинальных линек и превративших крыло в тонкую легкую двуслойную кутикулу – стрекоз. Эти воздушные хищники оказались таким же «абсолютным оружием», как некогда зоопланктон миллиметрового размерного класса, спровоцировавший некогда вендский фитопланктонный кризис (см. главу 6-а).
По мнению В.В. Жерихина (1990), именно стрекозы полностью уничтожили характерную для карбона фауну крупноразмерных открытоживущих палинофагов типа диктионеврид (эта смена была одной из самых резких за всю историю класса), заставив крылатых насекомых специализироваться в двух направлениях. Одним вариантом было совершенствовать свой полет (попутно при этом уменьшаясь в размерах), а также тип метаморфоза – появление полного превращения позволяет спрятать червеобразную личинку и неподвижную куколку внутри субстрата. Эта линия эволюции привела в конечном счете к возникновению в начале мезозоя (триас) наиболее процветающих ныне отрядов – двукрылых и перепончатокрылых, а чуть позже – бабочек. Другой выход – перейти к скрытному образу жизни (например, внутри уже знакомой нам валежникообразной «подстилки» карбоновых лесов), превратив переднюю пару крыльев в крышеобразно складываемые жесткие надкрылья, защищающие от повреждений заднюю пару, которая и выполняет летательные функции; по этому пути пошли гриллоновые насекомые, сохраняющие неполный метаморфоз – тараканы, прямокрылые и их родственники, а из насекомых с полным превращением – жуки, самый большой из современных отрядов (более ста тысяч видов), появившийся в начале перми.
Хотя в карбоне уже существовали стрекозы и поденки, личинки их (в отличие от современных) были существами не водными, а наземными. Освоение насекомыми пресных вод началось лишь во второй половине следующего (пермского) периода, когда появились отряды, имеющие водную личинку – веснянки, ручейники, а также поденки современного типа (личинки же стрекоз перестали быть наземными лишь в мезозое). В этой связи В.В. Жерихин (1980) указывает на следующее обстоятельство. Как известно, смыв различных веществ (прежде всего – микроэлементов) должен сильно обеднять почвы водоразделов. В настоящее время подобное обеднение если не предотвращается, то заметно тормозится прямым выносом вещества из водоемов на сушу животными (например, азот и фосфор, смытые с суши в море, оказываются – по пищевым цепям – в организмах сперва рыб, затем рыбоядных морских птиц, а в конечном счете возвращаются на сушу в форме птичьего помета). В пресных водах эту функцию выполняют прежде всего насекомые, которые имеют водных личинок и имаго, способных к дальнему разлету из водоемов.
Что же касается карбона, когда насекомые с водными личинками, судя по всему, отсутствовали, то в те времена обеднение почв на водоразделах (если там вообще формировались почвы) должно было быть особенно сильным. Не исключено, что формирование в перми, и особенно в триасе, достаточно обильной фауны насекомых с водными личинками стало одним из тех факторов, которые изменили почвенную ситуацию на водоразделах и позволили растительности начать «наступление» на первичные пустыни. Важно подчеркнуть, что этот вынос микроэлементов насекомыми мог сыграть здесь роль первого толчка – дальше система будет развиваться с положительной обратной связью, ибо возникшая на водоразделах растительность сама является мощнейшим регулятором стока. Окончательно эта система могла оформиться в позднем триасе – когда появились двукрылые; именно такие их представители, как, например, комары-звонцы (Chironomidae) совмещают огромную биомассу со способностью к дальнему разлету (в том числе за счет пассивного переноса ветром).
В заключении остановимся на одном существенном отличии экосистем древних (палеозойских и мезозойских) озер от современных. Это полное отсутствие погруженных макрофитов, за исключением харовых водорослей (которые депонируют биогены почти на порядок хуже, чем покрытосеменные). Основными продуцентами там скорее всего были плавающие маты, причем в мезозое получили распространение водорослевые маты на каркасе из высших растений. В триасе это были плевромейи – крупные плавающие плауновидные (что-то вроде сильно увеличенного полушника), по виду напоминающие морской буй, где в роли верхней сигнальной лампочки выступает пучок листьев и спорангии; листья, скорее всего, плавали по поверхности воды, постепенно отгнивая с вершины [48]. На каркасе из этих плауновидных возникали плавающие острова из мелких водяных мхов и печеночников (типа любимой аквариумистами риччии), а снаружи все это обрастало зелеными водорослями и цианобактериями; на этих матах существовала богатая фауна беспозвоночных: в палеозое – сходные с многоножками растительноядные эвтикарциноиды, позже – ракообразные и насекомые.
Поскольку все фотосинтезирующие структуры были сосредоточены на поверхности озера (а дно было к тому же еще и затенено), гиполимнион почти всегда был асфиксным и безжизненным, а озера легко эвтрофицировались. Однако в непосредственной близости от плавающего мата концентрация кислорода в воде могла быть очень высока; возможно, именно этим объясняется присутствие в составе фауны эвтрофных (судя по составу отложений) мезозойских озер целого ряда групп насекомых, являющихся ныне реофилами (обитателями быстрых, богатых кислородом рек и ручьев). Кроме того, в составе этих фаун комбинируются как пресноводные формы, имеющие адаптации к водному образу жизни, так и наземные (которые могли «ходить пешком» по поверхности плавающего мата). Эти имевшие огромную биомассу и богатые животным белком «плавучие острова» могли составлять основу питания для многих позвоночных, включая и динозавров. Окончательно экосистемы такого типа исчезли лишь в кайнозое, когда на дне водоемов возникли заросли погруженных макрофитов из покрытосеменных (типа элодеи), резко увеличившие разнообразие и устойчивость пресноводных экосистем. Это и дало А.Г. Пономаренко основание разделить всю историю континентальных водоемов на два принципиальных этапа: первый – от докембрия до середины палеогена (когда основными продуцентами были маты, сперва прикрепленные, затем – плавающие), а второй – от середины палеогена доныне (когда эту роль стали играть погруженные макрофиты).
10. Эволюция наземных позвоночных (1): поздний палеозой – ранний мезозой. Анамнии и амниоты. Две линии амниот – тероморфная и завроморфная.
Мы с вами расстались с самыми первыми тетраподами (девонской ихтиостегой и родственными ей формами) когда те... Чуть было не сказал: «Делали первые шаги по суше», но нет – мы ведь теперь знаем, что существа те были чисто водными... Скажем лучше так, более осторожно: «Получили принципиальную возможность периодически покидать водную среду». Да и что они, собственно говоря, могли бы найти на той суше – кроме перегрузок из-за многократного увеличения собственного веса, теплового удара да смертоносного обезвоживания организма?.. Пищу? Какую пищу – в девонском-то периоде? В общем, в девоне целенаправленный «выход на сушу» был бы даже не авантюрой, а чем-то по разряду мазохизма. Есть пятипалая конечность, позволяющая при необходимости переползти из совсем уже пересохшей лужи в не совсем пересохшую – и ладно...
Ситуация принципиально изменилась с начала карбона, когда в уже знакомых нам «лесах-водоемах» возникла богатая фауна из амфибиотических и наземных членистоногих – паукообразные, многоножки, а потом и насекомые: за этот ресурс уже стоило побороться. Среди карбоновых амфибий появляются небольшие (менее 1 м), похожие на современных саламандр антракозавры, имеющие несомненные приспособления к наземной жизни и явно ориентированные на питание беспозвоночными (а не рыбой – как их более крупные родственники). Именно антракозавров считают предками рептилий [49]; первые рептилии – карбоновые капториниды – напоминали по внешнему виду крупных ящериц, причем, судя по строению их челюстного аппарата, они специализировались именно на питании насекомыми. Впрочем, не будем забегать вперед.
Чем объяснить итоговый эволюционный неуспех амфибий? Ведь этот класс процветал только в карбоновом периоде (причем – только на территории тогдашних тропиков, в Еврамерии: ни в Ангариде, ни в Гондване по сию пору не найдено ни единой косточки карбонового возраста [50]), когда у него не было конкурентов; во все же остальные времена они занимали сугубо подчиненное положение в экосистемах – как наземных, так и пресноводных. Судя по всему, амфибий сгубила даже не облигатная связь с водой (они не выработали яйца и размножаются икрой), а неисправимое несовершенство их дыхательной системы, предопределившее целую цепь пренеприятнейших анатомо-физиологических следствий. Давайте по порядку.
Из курса зоологии вы должны помнить, что лягушка не имеет ни ребер, ни диафрагмы. Поэтому она – в отличие от нас – неспособна делать вдох, увеличивая объем замкнутой грудной полости, и вынуждена нагнетать воздух в легкие крайне несовершенным способом: набирать его в ротовую полость, замыкать ее, а потом «проглатывать» воздух, поднимая дно ротовой полости и сокращая ее объем. Итак, легочное дыхание у амфибий развито слабо.
Слабость легочного дыхания не позволяет амфибиям освободить от дыхательных функций кожу. Об этом обычно упоминают в той связи, что земноводные должны всегда сохранять покровы влажными (именно в этой влаге и растворяется воздушный кислород, «всасываемый» затем кожей), а потому они навсегда прикованы к воде. Это верно – однако это еще полбеды. Хуже другое: не освободив от дыхательных функций кожу, невозможно изолировать друг от друга большой и малый круги кровообращения и отделить венозную кровь от артериальной. Появление легких у тетрапод привело к возникновению двух кругов кровообращения; при этом правая сторона сердца становится «венозной», а левая – «артериальной». У амфибий сердце трехкамерное – два предсердия и желудочек; у рептилий – формально! – тоже трехкамерное, но в желудочке возникает неполная продольная перегородка, делящая его на «венозную» и «артериальную» половины. В ходе дальнейшей эволюции перегородка становится полной, а сердце – четырехкамерным (хотя у крокодилов с птицами и у млекопитающих это происходит по-разному). У амфибий же кислород приносят в сердце вены, идущие не только от легких (малый круг) но и от кожи (большой круг), и потому создавать какую-либо перегородку между правой и левой половинами желудочка бессмысленно, а кровь в сердце всегда будет смешанной – артериально-венозной. Пока же кровь не разделена на артериальную и венозную, уровень энергетического обмена организма будет принципиально невысоким. К тому же при постоянно влажных покровах любое повышение температуры тела будет тут же сводиться на нет за счет поверхностного испарения – термическое охлаждение. Все это полностью закрывает амфибиям путь к достижению любых вариантов эндотермии («теплокровности») [51]– с понятными эволюционными последствиями.
Из-за единственно доступного для амфибий способа наполнения легких («проглатывания» воздуха – гулярное дыхание) необходимо сохранять «подчелюстной насос», поднимающий и опускающий дно ротовой полости. Размещение последнего требует совершенно специфической («лягушачьей») формы черепа – очень широкой и приплюснутой. При этом челюстная мускулатура оказывается размещенной крайне невыгодным образом: ее можно подсоединить лишь к самому челюстному суставу, с задней его стороны; получающийся в результате рычаг будет крайне неэффективен – все равно, как если бы мы затворяли дверь, упираясь не в ручку, а около петель. Для того, чтобы развивать при помощи таких челюстей сколь-нибудь приличные усилия, приходится наращивать объем челюстной мускулатуры; в итоге эта мускулатура занимает практически все пространство черепа, попросту не оставляя места для мозговой коробки. То есть амфибии – существа принципиально «безмозглые», и это закрывает им путь к сложным формам поведения.
Отсюда понятно, как следует бороться (в эволюционном смысле) со всем этим комплексом неприятностей: надо радикально перестроить дыхательную систему. Именно анализируя переход от амфибий к рептилиям, Н.Н. Иорданский (1977) выдвинул свой принцип ключевого ароморфоза – изменение в одной системе органов, ведущее к целому каскаду последовательных прогрессивных перестроек всего организма.
Достаточно наладить нормальное легочное дыхание (путем изменения объема грудной полости), как приведенные выше цепочки начинают «раскручиваться» в обратную сторону. Убрав «подчелюстной насос», мы можем сделать череп высоким и узким, подвести жевательную мускулатуру к челюсти не сзади, а сверху (как у нас с вами), уменьшить – за счет улучшения рычага – ее объем и отдать освободившееся место под «мозги»; кроме того, такое расположение жевательных мышц позволит в дальнейшем не только удерживать схваченную добычу, но и пережевывать пищу (рисунок 37). Освободив кожу от дыхательных функций, мы получаем возможность разделить круги кровообращения и резко интенсифицировать обмен веществ. Эти инженерные решения вполне однозначны, равно как и необходимость одеть икринку особой оболочкой – амнионом, как бы создающей для зародыша маленький искусственный водоем и делающим его развитие независимым от водной среды; отсюда фундаментальное разделение позвоночных на «прикованных к воде» анамний (рыб и амфибий), и «истинно сухопутных» амниот – рептилий, птиц и млекопитающих). А вот дальше начинается интереснейший эволюционный выбор, когда вариантов – два, в обоих есть свои плюсы и свои минусы.
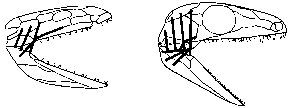
РИСУНОК 37. Расположение жевательной мускулатуры лабиринтодонта Ichthyostega (а) и примитивной капториноморфной рептилии Paleothyrus (б).
Главная проблема, которую нам предстоит решать теперь, когда осуществлен настоящий выход на сушу – это экономия воды. Возникает вопрос: как нам быть с кожей, доставшейся в наследство от амфибий – мягкой и влажной, богатой железами? Можно ее полностью заизолировать, создав на поверхности водонепроницаемый роговой слой – а можно лишь модифицировать, сохранив ее основные характеристики; оба решения вполне реализуемы, и каждое из них влечет за собою целую цепь физиологических следствий.
Создав сухую кожу с роговым покрытием, мы сводим к минимуму потери влаги: организм становится практически независимым от внешних источников воды (как одетые в «пустынные скафандры» герои фантастического романа «Дюна»). Однако за удобство надо платить.
Во-первых, необходимо перестроить выделительную систему. Почки амфибий функционально не отличаются от рыбьих и предназначены для выведения из организма избытка воды (которую тот постоянно «насасывает» за счет осмоса из гипотонической внешней среды). Проблема удаления конечного продукта белкового обмена – весьма токсичной мочевины CO(NH2)2 – решается в этом случае элементарно: ее просто растворяют в водяном потоке, который так и так постоянно «течет сквозь организм». Иное дело – когда мы начинаем экономить воду, потребляя ее по минимуму; легко создать канализацию в населенном пункте, стоящем на реке, а что делать, если в твоем распоряжении лишь артезианский колодец? Тогда «почки выведения» необходимо заменить на «почки сбережения», призванные выводить во внешнюю среду как можно меньше воды. При этом приходится менять конечный продукт белкового обмена с мочевины на менее токсичную мочевую кислоту – а этот дополнительный «технологический цикл» весьма энергоемок [52].
Во-вторых (и это даже более важно), сухая, лишенная желез кожа создает большие трудности с терморегуляцией – а при жизни на суше, где обычны резкие температурные перепады, эта проблема одна из важнейших. При голых, ороговевших покровах равно затруднены и теплоизоляция (в холод), и теплосброс (в жару) – потому для этой эволюционной линии путь к достижению гомойотермии (поддержанию постоянной, независимой от внешней среды температуры тела) если и не закрыт полностью, то очень сильно затруднен.
Если же мы решим не гнаться за экономией воды и оставим кожу железистой, не ороговевшей, то перед нами откроется множество новых возможностей. Эти кожные железы можно преобразовать в самые разнообразные структуры. Можно превратить их в волоски, выполняющие осязательные функции, а сделав эти волоски достаточно густыми, создать теплоизолирующий покров – шерсть. Можно превратить их дополнительный орган выделения – потовые железы, которые являются еще и терморегулятором (пот, испаряясь, охлаждает поверхность тела); можно, наконец, изменить состав их выделений, превратив эти железы в млечные, и выкармливать с их помощью детенышей. Как легко видеть, в этой эволюционной линии, с ее терморегуляторными возможностями, появление гомойотермии просто-таки напрашивается – да так оно и было на самом деле. Правда, по степени зависимости от источников воды эти существа будут не слишком превосходить амфибий – но ведь за всё в этом мире приходится платить... А с другой стороны – раз уж внутренняя среда нашего организма все равно будет существовать как бы в постоянном водяном токе, нет нужды радикально перестраивать почки на «водосберегающую технологию» и возиться со сменой конечного продукта азотного обмена!
Что же это за «две эволюционные линии»? Традиционный взгляд на эволюцию рептилий – он и поныне отражен во многих учебниках – предполагает, что от каких-то антракозавров (здесь часто поминают сеймурию, имевшую уже рептилийное строение челюсти и грудной клетки, пригодной для реберного дыхания, но еще не расставшуюся с водой – личинки ее так и оставались «головастиками» с наружными жабрами) происходит некий гипотетический «общий предок пресмыкающихся», который обладает уже всем набором рептилийных признаков (водонепроницаемой кожей, почками сбережения и т.д.). От него-то и берут начало основные эволюционные ветви этого класса – анапсиды, синапсиды диапсиды и эвриапсиды; этим группам зачастую придают статус подклассов. Они выделены на основе того, каким способом в каждом из них трансформировался исходный стегальный (крышевидный) череп лабиритодонтов (рисунок 38). У анапсид (сюда относятся черепахи и ряд вымерших групп, например, парейазавры) сохраняется исходный тип черепа – массивный, лишенный облегчающих конструкцию отверстий. У синапсид – «зверообразных ящеров» , чьи зубы были дифференцированы по форме и функциям (пеликозавров и более продвинутых терапсид, которых традиционно считают предками млекопитающих) возникает одно височное окно, от краев которого отходят челюстные мышцы, и возникает единственная скуловая дуга. У диапсид же височных окон возникает два, и соответственно, две скуловых дуги – верхняя и нижняя; к этой группе принадлежат большинство рептилий, как современных (ящерицы со змеями, гаттерии и крокодилы), так и вымерших (динозавры, птерозавры). К эвриапсидам относят несколько вымерших морских групп (ихтиозавров, плезиозавров и родственных им нотозавров и плакодонтов); их черепа имеют единственное височное окно, но образуется оно иным образом, чем у синапсид
Ситуация принципиально изменилась с начала карбона, когда в уже знакомых нам «лесах-водоемах» возникла богатая фауна из амфибиотических и наземных членистоногих – паукообразные, многоножки, а потом и насекомые: за этот ресурс уже стоило побороться. Среди карбоновых амфибий появляются небольшие (менее 1 м), похожие на современных саламандр антракозавры, имеющие несомненные приспособления к наземной жизни и явно ориентированные на питание беспозвоночными (а не рыбой – как их более крупные родственники). Именно антракозавров считают предками рептилий [49]; первые рептилии – карбоновые капториниды – напоминали по внешнему виду крупных ящериц, причем, судя по строению их челюстного аппарата, они специализировались именно на питании насекомыми. Впрочем, не будем забегать вперед.
Чем объяснить итоговый эволюционный неуспех амфибий? Ведь этот класс процветал только в карбоновом периоде (причем – только на территории тогдашних тропиков, в Еврамерии: ни в Ангариде, ни в Гондване по сию пору не найдено ни единой косточки карбонового возраста [50]), когда у него не было конкурентов; во все же остальные времена они занимали сугубо подчиненное положение в экосистемах – как наземных, так и пресноводных. Судя по всему, амфибий сгубила даже не облигатная связь с водой (они не выработали яйца и размножаются икрой), а неисправимое несовершенство их дыхательной системы, предопределившее целую цепь пренеприятнейших анатомо-физиологических следствий. Давайте по порядку.
Из курса зоологии вы должны помнить, что лягушка не имеет ни ребер, ни диафрагмы. Поэтому она – в отличие от нас – неспособна делать вдох, увеличивая объем замкнутой грудной полости, и вынуждена нагнетать воздух в легкие крайне несовершенным способом: набирать его в ротовую полость, замыкать ее, а потом «проглатывать» воздух, поднимая дно ротовой полости и сокращая ее объем. Итак, легочное дыхание у амфибий развито слабо.
Слабость легочного дыхания не позволяет амфибиям освободить от дыхательных функций кожу. Об этом обычно упоминают в той связи, что земноводные должны всегда сохранять покровы влажными (именно в этой влаге и растворяется воздушный кислород, «всасываемый» затем кожей), а потому они навсегда прикованы к воде. Это верно – однако это еще полбеды. Хуже другое: не освободив от дыхательных функций кожу, невозможно изолировать друг от друга большой и малый круги кровообращения и отделить венозную кровь от артериальной. Появление легких у тетрапод привело к возникновению двух кругов кровообращения; при этом правая сторона сердца становится «венозной», а левая – «артериальной». У амфибий сердце трехкамерное – два предсердия и желудочек; у рептилий – формально! – тоже трехкамерное, но в желудочке возникает неполная продольная перегородка, делящая его на «венозную» и «артериальную» половины. В ходе дальнейшей эволюции перегородка становится полной, а сердце – четырехкамерным (хотя у крокодилов с птицами и у млекопитающих это происходит по-разному). У амфибий же кислород приносят в сердце вены, идущие не только от легких (малый круг) но и от кожи (большой круг), и потому создавать какую-либо перегородку между правой и левой половинами желудочка бессмысленно, а кровь в сердце всегда будет смешанной – артериально-венозной. Пока же кровь не разделена на артериальную и венозную, уровень энергетического обмена организма будет принципиально невысоким. К тому же при постоянно влажных покровах любое повышение температуры тела будет тут же сводиться на нет за счет поверхностного испарения – термическое охлаждение. Все это полностью закрывает амфибиям путь к достижению любых вариантов эндотермии («теплокровности») [51]– с понятными эволюционными последствиями.
Из-за единственно доступного для амфибий способа наполнения легких («проглатывания» воздуха – гулярное дыхание) необходимо сохранять «подчелюстной насос», поднимающий и опускающий дно ротовой полости. Размещение последнего требует совершенно специфической («лягушачьей») формы черепа – очень широкой и приплюснутой. При этом челюстная мускулатура оказывается размещенной крайне невыгодным образом: ее можно подсоединить лишь к самому челюстному суставу, с задней его стороны; получающийся в результате рычаг будет крайне неэффективен – все равно, как если бы мы затворяли дверь, упираясь не в ручку, а около петель. Для того, чтобы развивать при помощи таких челюстей сколь-нибудь приличные усилия, приходится наращивать объем челюстной мускулатуры; в итоге эта мускулатура занимает практически все пространство черепа, попросту не оставляя места для мозговой коробки. То есть амфибии – существа принципиально «безмозглые», и это закрывает им путь к сложным формам поведения.
Отсюда понятно, как следует бороться (в эволюционном смысле) со всем этим комплексом неприятностей: надо радикально перестроить дыхательную систему. Именно анализируя переход от амфибий к рептилиям, Н.Н. Иорданский (1977) выдвинул свой принцип ключевого ароморфоза – изменение в одной системе органов, ведущее к целому каскаду последовательных прогрессивных перестроек всего организма.
Достаточно наладить нормальное легочное дыхание (путем изменения объема грудной полости), как приведенные выше цепочки начинают «раскручиваться» в обратную сторону. Убрав «подчелюстной насос», мы можем сделать череп высоким и узким, подвести жевательную мускулатуру к челюсти не сзади, а сверху (как у нас с вами), уменьшить – за счет улучшения рычага – ее объем и отдать освободившееся место под «мозги»; кроме того, такое расположение жевательных мышц позволит в дальнейшем не только удерживать схваченную добычу, но и пережевывать пищу (рисунок 37). Освободив кожу от дыхательных функций, мы получаем возможность разделить круги кровообращения и резко интенсифицировать обмен веществ. Эти инженерные решения вполне однозначны, равно как и необходимость одеть икринку особой оболочкой – амнионом, как бы создающей для зародыша маленький искусственный водоем и делающим его развитие независимым от водной среды; отсюда фундаментальное разделение позвоночных на «прикованных к воде» анамний (рыб и амфибий), и «истинно сухопутных» амниот – рептилий, птиц и млекопитающих). А вот дальше начинается интереснейший эволюционный выбор, когда вариантов – два, в обоих есть свои плюсы и свои минусы.
РИСУНОК 37. Расположение жевательной мускулатуры лабиринтодонта Ichthyostega (а) и примитивной капториноморфной рептилии Paleothyrus (б).
Главная проблема, которую нам предстоит решать теперь, когда осуществлен настоящий выход на сушу – это экономия воды. Возникает вопрос: как нам быть с кожей, доставшейся в наследство от амфибий – мягкой и влажной, богатой железами? Можно ее полностью заизолировать, создав на поверхности водонепроницаемый роговой слой – а можно лишь модифицировать, сохранив ее основные характеристики; оба решения вполне реализуемы, и каждое из них влечет за собою целую цепь физиологических следствий.
Создав сухую кожу с роговым покрытием, мы сводим к минимуму потери влаги: организм становится практически независимым от внешних источников воды (как одетые в «пустынные скафандры» герои фантастического романа «Дюна»). Однако за удобство надо платить.
Во-первых, необходимо перестроить выделительную систему. Почки амфибий функционально не отличаются от рыбьих и предназначены для выведения из организма избытка воды (которую тот постоянно «насасывает» за счет осмоса из гипотонической внешней среды). Проблема удаления конечного продукта белкового обмена – весьма токсичной мочевины CO(NH2)2 – решается в этом случае элементарно: ее просто растворяют в водяном потоке, который так и так постоянно «течет сквозь организм». Иное дело – когда мы начинаем экономить воду, потребляя ее по минимуму; легко создать канализацию в населенном пункте, стоящем на реке, а что делать, если в твоем распоряжении лишь артезианский колодец? Тогда «почки выведения» необходимо заменить на «почки сбережения», призванные выводить во внешнюю среду как можно меньше воды. При этом приходится менять конечный продукт белкового обмена с мочевины на менее токсичную мочевую кислоту – а этот дополнительный «технологический цикл» весьма энергоемок [52].
Во-вторых (и это даже более важно), сухая, лишенная желез кожа создает большие трудности с терморегуляцией – а при жизни на суше, где обычны резкие температурные перепады, эта проблема одна из важнейших. При голых, ороговевших покровах равно затруднены и теплоизоляция (в холод), и теплосброс (в жару) – потому для этой эволюционной линии путь к достижению гомойотермии (поддержанию постоянной, независимой от внешней среды температуры тела) если и не закрыт полностью, то очень сильно затруднен.
Если же мы решим не гнаться за экономией воды и оставим кожу железистой, не ороговевшей, то перед нами откроется множество новых возможностей. Эти кожные железы можно преобразовать в самые разнообразные структуры. Можно превратить их в волоски, выполняющие осязательные функции, а сделав эти волоски достаточно густыми, создать теплоизолирующий покров – шерсть. Можно превратить их дополнительный орган выделения – потовые железы, которые являются еще и терморегулятором (пот, испаряясь, охлаждает поверхность тела); можно, наконец, изменить состав их выделений, превратив эти железы в млечные, и выкармливать с их помощью детенышей. Как легко видеть, в этой эволюционной линии, с ее терморегуляторными возможностями, появление гомойотермии просто-таки напрашивается – да так оно и было на самом деле. Правда, по степени зависимости от источников воды эти существа будут не слишком превосходить амфибий – но ведь за всё в этом мире приходится платить... А с другой стороны – раз уж внутренняя среда нашего организма все равно будет существовать как бы в постоянном водяном токе, нет нужды радикально перестраивать почки на «водосберегающую технологию» и возиться со сменой конечного продукта азотного обмена!
Что же это за «две эволюционные линии»? Традиционный взгляд на эволюцию рептилий – он и поныне отражен во многих учебниках – предполагает, что от каких-то антракозавров (здесь часто поминают сеймурию, имевшую уже рептилийное строение челюсти и грудной клетки, пригодной для реберного дыхания, но еще не расставшуюся с водой – личинки ее так и оставались «головастиками» с наружными жабрами) происходит некий гипотетический «общий предок пресмыкающихся», который обладает уже всем набором рептилийных признаков (водонепроницаемой кожей, почками сбережения и т.д.). От него-то и берут начало основные эволюционные ветви этого класса – анапсиды, синапсиды диапсиды и эвриапсиды; этим группам зачастую придают статус подклассов. Они выделены на основе того, каким способом в каждом из них трансформировался исходный стегальный (крышевидный) череп лабиритодонтов (рисунок 38). У анапсид (сюда относятся черепахи и ряд вымерших групп, например, парейазавры) сохраняется исходный тип черепа – массивный, лишенный облегчающих конструкцию отверстий. У синапсид – «зверообразных ящеров» , чьи зубы были дифференцированы по форме и функциям (пеликозавров и более продвинутых терапсид, которых традиционно считают предками млекопитающих) возникает одно височное окно, от краев которого отходят челюстные мышцы, и возникает единственная скуловая дуга. У диапсид же височных окон возникает два, и соответственно, две скуловых дуги – верхняя и нижняя; к этой группе принадлежат большинство рептилий, как современных (ящерицы со змеями, гаттерии и крокодилы), так и вымерших (динозавры, птерозавры). К эвриапсидам относят несколько вымерших морских групп (ихтиозавров, плезиозавров и родственных им нотозавров и плакодонтов); их черепа имеют единственное височное окно, но образуется оно иным образом, чем у синапсид