Страница:
Один из основоположников этого направления, энтомолог Л. Брундин (1966), изучал комаров-звонцов (Chironomidae) Южного полушария. Он показал, что они резко отличны от всех северополушарных (что отражает давнюю их изоляцию на территории Гондваны); среди южных хирономид наиболее архаичные, уклоняющиеся, формы обитают в Южной Африке, а наиболее близки между собой обитатели Австралии и Южной Америки. Эта последовательность родственных связей вполне соответствует порядку, в котором расходились фрагменты Пангеи: раньше всего произошел раскол на Лавразию и Гондвану (триас), внутри Гондваны же последовательно теряли связь друг с другом (через Антарктиду) Южная Африка (юра), Новая Зеландия (мел), и, наконец, Австралия и Южная Америка (эоцен и олигоцен) (рисунок 64). Позже такие последовательности были построены для множества южнополушарных («гондванских») групп, главным образом для насекомых и иных наземных и пресноводных беспозвоночных.
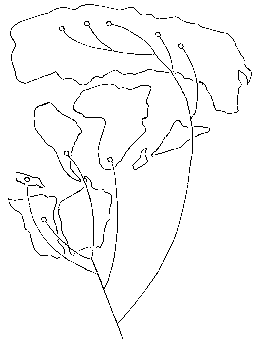
РИСУНОК 64. Схема Брундина, отражающая степень родства между комарами-звонцами, обитающими в Южной Америке (ЮА), Австралии (АВ), Новой Зеландии (НЗ), на юге Африки (АФ) и на блоке Северных континентов, в Лавразии (ЛВ). Порядок ветвлений кладограммы действительно совпадает с порядком, в котором теряли между собою связь фрагменты Пангеи.
Однако веские подтверждения получила в последнее время и уоллесова теория «оттесненных реликтов». По мере изученния мезозойских и палеогеновых насекомых стало ясно, что многие, если не большинство, из «крайне-южных» групп, которыми оперируют сторонники викариантной биогеографии (в том числе – те хирономиды, с которыми имел дело Брундин), в действительности были ранее широко распространены в Северном полушарии. Если искать современные аналоги той энтомофауне, что содержится в верхнемеловых янтарях Сибири и Канады, то на память приходит Новая Зеландия. Накапливается все больше аргументов в пользу того, что экзотическая биота, которая занимает ныне изолированные участки на крайнем юге Южных континентов, в мезозое имела почти всесветное распространение. Сторонники теории «оттесненных реликтов» не отрицают того, что некоторые из южнополушарных ареалов могли возникнуть в результате раскола Гондваны и расхождения ее фрагментов; они лишь подчеркивают, что распознать такие случаи среди дизъюнкций, достоверно возникших в результате вымирания «связующих звеньев» на Северных материках, в принципе невозможно.
Модели Уоллеса и Гукера, возникнув из в общем-то частной проблемы – происхождения биоты Южных материков, и акцентируя внимание в первом случае на ее архаичности и реликтовости, а во втором – на ее единстве (и обеспечивающих это единство механизмах), позже трансформировались в две альтернативные научные традиции (нечто вроде «линии Платона» и «линии Демокрита» в античной философии, приведших позднее к идеализму и материализму). К этим двум линиям в принципе может быть сведено все многообразие концепций и моделей, существующих в исторической биогеографии. Однако здесь (как и в иных случаях длительного сосуществования научных концепций) возникает впечатление, что «линия Уоллеса» и «линия Гукера» не взаимоисключающи, а взаимодополнительны: они просто рассматривают предмет исследования (ареал таксона) с разных сторон, лишь в паре дают о нем адекватное представление. В гукеровской модели таксон принимается как неизменная данность, а предметом анализа становится его расселение (преграды и их преодоление); в уоллесовской же модели исследуют таксонообразование и вымирание, полагая расселительный процесс относительно малосущественной деталью. Можно сказать, что уоллесовская модель пытается спроецировать время в пространство, а гукеровская – пространство во время.
В своей исходной версии уоллесовская модель предполагала, что основной центр формообразования находится на блоке Северных континентов, откуда всяческая архаика вытесняется на юг. Позже, однако, существование по всей планете единого, если так можно сказать, «градиента архаичности» от Югу к Севера было отвергнуто; в частности, было установлено, что представления о глубокой древности и консервативности тропической биоты не соответствуют действительности (см. главу 13). На самом деле реликты концентрируются во внетропических областях (главным образом – в субтропиках) обоих полушарий, хотя в Южном, по ряду причин, сильнее, чем в Северном.
Реликтовый ареал таксона, распространенного во внетропических областях обоих полушарий, называют биполярным или амфитропическим. Ареал этот может в дальнейшем подвергаться различным модификациям. Упомянем в этой связи предложенную К.Ю. Еськовым (1984) модель «циркумпацифического домино», основанную на распространении современных и вымерших беспозвоночных и растений в Тихоокеанском регионе. Она предстваляет собой нечто вроде «периодической системы» транспацифических дизьюнкций: все их варианты рассматриваются как последовательные стадии сокращения единого биполярного ареала (рисунок 65). Редукция северной его «дольки» (рисунок 65, 6) и породит тот самый «гондванский ареал», которым оперирует викариантная биогеография. Обратная ситуация (редукция южной «дольки» – рисунок 65, 2) наблюдается заметно реже, однако существует и целый ряд таксонов, вымерших в Южном полушарии и сохранившихся в Северном (таковы, например, хвостатые амфибии и насекомые из отрядов верблюдок и тараканосверчков). Некоторые авторы связывают эту асимметрию просто с худшей палеонтологической изученностью Южных материков; и действительно, за последние годы, после открытия крупных местонахождений мезозойских насекомых в Австралии и Бразилии, число случаев документированного вымирания в Гондване «лавразийских» ныне таксонов возросло вдвое!
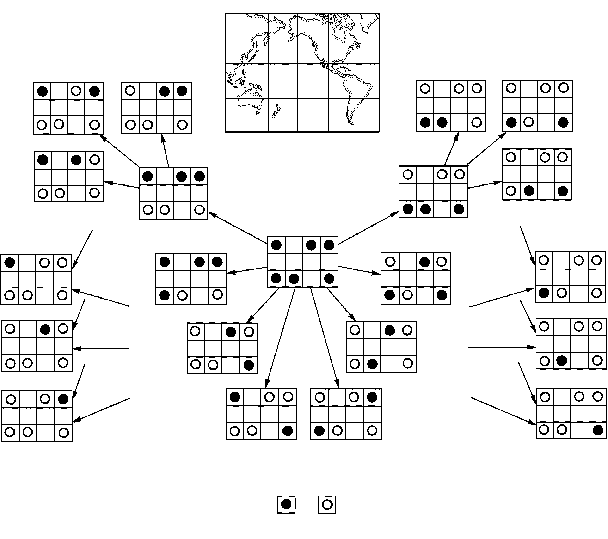
РИСУНОК 65. «Циркумпацифическое домино» – вероятный ход сокращения первоначально единого биполярного циркумпацифического ареала. I – группа присутствует в регионе, II – группа вымерла; а – внетропическая восточная Азия, б – запад Северной Америки, в – восток Северной Америки, г – внетропическая Австралия, д – Новая Зеландия, е – внетропическая Южная Америка. Примеры ареалов: 1 – пауки подотряда Hypochilomorphae, 2 – хвойные рода Tsuga, 3 – насекомые отряда Grylloblattida, 4 – гигантские саламандры Cryptobranchiidae, 5 – саламандры семейства Ambistomatidae, 6 – клопы подотряда Peloridiina, 7 – цветковые семейства Tetrachondraceae, 8 – цветковые семейства Eucryphiaceae, 9 – ручейники семейства Helicophiidae, 10 – хвойные семейства Taxodiaceae, 11 – насекомые-вислокрылки рода Protochauloides, 12 – пауки-птицеяды семейства Mecicobothriidae, 13 – лягушки семейства Leiopelmidae, 14 – цветковые семейства Lardisabalaceae, 15 – скорпионницы семейства Meropeidae.
Итак, ключевой проблемой для уоллесовской схемы оказывается механизм образования такого исходного ареала, из которого можно вывести все остальные: биполярного, биполярной пары «сестринских» таксонов (таковы, например, буки и нотофагусы), или же такого типа распространения, когда более продвинутые субтаксоны приурочены к тропикам, а более архаичные – к внетропическим областям. Необходимое для этого вымирание таксона на территории тропиков – с замещением его более продвинутыми формами или без такового – находит объяснение в двух альтернативных моделях: «экваториальной помпы» и «зональной стратификации».
«Экваториальная помпа» (термин принадлежит Ф. Дарлингтону) предполагает, что именно в тропических экосистемах с их стабильностью среды и высоким разнообразием идет постоянное и наиболее активное формообразование. При этом реликты «выдавливаются» (помпа!) и на север, и на юг, за счет конкуренции со стороны более молодых, продвинутых таксонов. Наиболее завершенный вид эта модель приобрела в работе С.В. Мейена (1987), основанной на палеоботаническом материале. Он показал, что в течение палеозоя почти все надродовые таксоны высших растений появлялись в экваториальном поясе в более древних стратиграфических слоях, чем за его пределами, и, следовательно, имеют экваториальное происхождение (так, доминирующие в Гондване и Ангариде глоссоптериевые и кордаиты первоначально возникли в тропиках Еврамерии – см. главу 9).
Именно тропический пояс является главной ареной макроэволюционных событий. Отсюда сформировавшиеся таксоны при потеплениях «растекаются» во внеэкваториальные широты (Мейен назвал это «фитоспредингом» – по аналогии со спредингом океанического дна), где при последующих похолоданиях они либо вымирают, либо эволюционируют в более узких (обычно внутриродовых) таксономических рамках; для внетропических областей на протяжении всей истории характерно длительное переживание архаических форм («внетропическое персистирование»). При этом процесс этот строго однонаправленный: вселения бореальных групп в тропики при похолоданиях не происходит никогда (рисунок 66). Причину повышенной интенсивности макроэволюции в тропиках Мейен видел в том, что здесь очень сильна биотическая конкуренция, но зато ослаблена неизбирательная абиотическая компонента отбора (холод, засухи, и т.д.). В результате естественный отбор менее жестко, чем в иных местах, выбраковывает крупные морфологические новации, давая шанс «перспективным монстрам» (термин Р. Гольдшмидта, 1940).

РИСУНОК 66. Филогения высших растений и географическая приуроченность основных филогенетических линий. 1 – нотальные (гондванские) внетропические области, 2 – экваториальные области, 3 – бореальные внетропические области, 4 – географическая ситуация неопределенна.
Альтернативный взгляд на природу амфитропических ареалов высказывал В.В. Жерихин (1978). Он основан на том, что в мезозое и раннем палеогене климатическая зональность в ее современном виде отсутствовала, и климат практически по всей планете был близок к нынешнему субтропическому. В конце палеогена произошла климатическая перестройка, в результате которой тепло стало распределяться по поверхности планеты менее равномерно, и на полюсах стало заметно холоднее, чем прежде, а на экваторе – жарче. В итоге единую «теплоумеренно-субтропическую» биоту Земли охватил процесс «зональной стратификации»; он заключался в том, что в экваториальных и приполярных широтах (где сменился климат) начались экосистемные перестройки, приведшие к достаточно массовому вымиранию мезозойских и раннепалеогеновых групп. Эти группы имели наилучшие шансы выжить в двух амфитропических «кольцах» с субтропическим и теплоумеренным климатом, господствовавшим ранее по всей планете – где они и поныне существуют в статусе реликтов (см. также главу 13).
Сравнение моделей «экваториальной помпы» и «зональной стратификации» позволяет прийти к достаточно неожиданному выводу о том, что они в действительности не альтернативны, а взаимодополнительны. Более того, К.Ю. Еськовым (1994) была предложена и обобщающая их концепция. Она состоит в том, что единый механизм мейеновского «фитоспрединга» может работать в различных режимах, диктуемых внешними (макроклиматическими) условиями, и «экваториальная помпа» есть просто один из этих рабочих режимов. Фитоспрединг понимается здесь как локализация макроэволюционных процессов (ведущих к возникновению новых таксонов) в тех районах Земли, где климатические условия наиболее благоприятны, и, соответственно, абиотическая компонента естественного отбора наиболее ослаблена; отсюда затем происходит однонаправленное «растекание» таксонов в районы с более жесткой абиотикой, где эти группы могут выживать и после своего вымирания в исходных районах. Фитоспрединг, таким образом, должен порождать целый ряд «градиентов архаичности»: гумидно-аридный, равнинно-высокогорный, океаническо-континентальный и т.д. Однако в эпохи резко выраженной широтной зональности климата (т.е. в криоэры) на всех них накладывается мощнейший температурный экваториально-полярный градиент, который низводит их до «градиентов второго порядка» и затушевывает их существование.
Именно в эти периоды фитоспрединг и работает в режиме «экваториальной помпы». В эпохи же с ослабленной широтной зональностью (термоэры) центры формообразования не локализованы в экваториальной зоне, а более диффузно распределены по всей планете. Число нарушений установленной Мейеном закономерности в стратиграфическом распределении членов таксона (в низких широтах они появляются в более древних слоях, чем в высоких) должно заметно возрастать в азональные эпохи – и анализ распределения позднепалеозойских и мезозойских групп насекомых подтвердил, что в целом это действительно так. Процесс же перехода биоты Земли от «полиградиентного» состояния (азональные эпохи) к «моноградиентному» (зональные эпохи) происходит в соответствии с моделью «зональной стратификации». Итак, фитоспрединг – это универсальный механизм флоро– и фауногенеза; он имеет три режима работы, определяемых глобальным термическим градиентом, и режимы эти формируют замкнутый трехчленный цикл: «экваториальная помпа» (поздний палеозой, криоэра) – «диффузные центры формообразования» (мезозой-ранний палеоген, термоэра) – «зональная стратификация» (поздний палеоген) – снова «экваториальная помпа» (неоген, криоэра), и т.д.
Рекомендуемая литература
В.А.Обручев «Плутония»; «Земля Санникова»
Ж. Рони Старший «Борьба за огонь»; «Пещерный лев»
М. Крайтон «Парк юрского периода»
С. Уэда «Новый взгляд на Землю»
А.С. Монин «Популярная история Земли»
Дж. Имбри, К.П. Имбри «Тайны ледниковых эпох»
Юл. Медведев «Судеб посланник» (в книге «Бросая вызов»)
И.А. Ефремов «Дорога ветров»
Дж. Смит «Старина четвероног»
А.К. Рождественский «На поиски динозавров в Гоби»
И. Аугуста, З. Буриан «По путям развития жизни»; «Летающие ящеры и древние птицы»; «Ящеры древних морей»; «Гиганты суши»; «Книга о мамонтах»; «Жизнь древнего человека»
К.Л. Фентон, М.А. Фентон «Каменная книга. Летопись доисторической жизни»
Ю.А. Орлов «О мире древних животных»
Д. Эттенборо «Жизнь на Земле»
И. Яковлева, В. Яковлев «По следам минувшего»
М.Ф. Ивахненко, В.А. Корабельников «Живое прошлое Земли»
А. Гангнус «Через горы времени»
Г.А. Заварзин «Бактерии и состав атмосферы Земли»
И.Н. Крылов «На заре жизни»
А.Ю. Розанов «Что произошло 600 миллионов лет назад»
С.В. Мейен «Из истории растительных династий»; «Следы трав индейских»
Н. Эйдельман «Ищу предка»
Р. Фоули «Еще один великолепный вид»
В.А. Красилов «Нерешенные вопросы теории эволюции»
Шишков «История климата»
Р. Кэрролл «Палеонтология и эволюция позвоночных» (3 тома)
В.В. Друщиц «Палеонтология беспозвоночных»
С.В. Мейен «Основы палеоботаники»; «Историческое развитие класса насекомых», Труды ПИН АН СССР, т.175
В.В. Жерихин «Развитие и смена меловых и кайнозойских фаунистических комплексов» (Общая часть)
Ф. Дарлингтон «Зоогеография» Дж. Симпсон «Великолепная изоляция»
С.М. Разумовский «Закономерности динамики фитоценозов»
О.Г. Кусакин, А.Л. Дроздов «Филема органического мира»
В.А. Красилов «Происхождение и ранняя эволюция цветковых растений»; «Эволюция и биостратиграфия»
Л.П. Татаринов «Очерки по теории эволюции»
Т. де Шарден «Феномен человека»
Э. Шредингер «Что такое жизнь с точки зрения физика»
И. Пригожин, И. Стенгерс «Порядок из хаоса»
Словарь терминов
РИСУНОК 64. Схема Брундина, отражающая степень родства между комарами-звонцами, обитающими в Южной Америке (ЮА), Австралии (АВ), Новой Зеландии (НЗ), на юге Африки (АФ) и на блоке Северных континентов, в Лавразии (ЛВ). Порядок ветвлений кладограммы действительно совпадает с порядком, в котором теряли между собою связь фрагменты Пангеи.
Однако веские подтверждения получила в последнее время и уоллесова теория «оттесненных реликтов». По мере изученния мезозойских и палеогеновых насекомых стало ясно, что многие, если не большинство, из «крайне-южных» групп, которыми оперируют сторонники викариантной биогеографии (в том числе – те хирономиды, с которыми имел дело Брундин), в действительности были ранее широко распространены в Северном полушарии. Если искать современные аналоги той энтомофауне, что содержится в верхнемеловых янтарях Сибири и Канады, то на память приходит Новая Зеландия. Накапливается все больше аргументов в пользу того, что экзотическая биота, которая занимает ныне изолированные участки на крайнем юге Южных континентов, в мезозое имела почти всесветное распространение. Сторонники теории «оттесненных реликтов» не отрицают того, что некоторые из южнополушарных ареалов могли возникнуть в результате раскола Гондваны и расхождения ее фрагментов; они лишь подчеркивают, что распознать такие случаи среди дизъюнкций, достоверно возникших в результате вымирания «связующих звеньев» на Северных материках, в принципе невозможно.
Модели Уоллеса и Гукера, возникнув из в общем-то частной проблемы – происхождения биоты Южных материков, и акцентируя внимание в первом случае на ее архаичности и реликтовости, а во втором – на ее единстве (и обеспечивающих это единство механизмах), позже трансформировались в две альтернативные научные традиции (нечто вроде «линии Платона» и «линии Демокрита» в античной философии, приведших позднее к идеализму и материализму). К этим двум линиям в принципе может быть сведено все многообразие концепций и моделей, существующих в исторической биогеографии. Однако здесь (как и в иных случаях длительного сосуществования научных концепций) возникает впечатление, что «линия Уоллеса» и «линия Гукера» не взаимоисключающи, а взаимодополнительны: они просто рассматривают предмет исследования (ареал таксона) с разных сторон, лишь в паре дают о нем адекватное представление. В гукеровской модели таксон принимается как неизменная данность, а предметом анализа становится его расселение (преграды и их преодоление); в уоллесовской же модели исследуют таксонообразование и вымирание, полагая расселительный процесс относительно малосущественной деталью. Можно сказать, что уоллесовская модель пытается спроецировать время в пространство, а гукеровская – пространство во время.
В своей исходной версии уоллесовская модель предполагала, что основной центр формообразования находится на блоке Северных континентов, откуда всяческая архаика вытесняется на юг. Позже, однако, существование по всей планете единого, если так можно сказать, «градиента архаичности» от Югу к Севера было отвергнуто; в частности, было установлено, что представления о глубокой древности и консервативности тропической биоты не соответствуют действительности (см. главу 13). На самом деле реликты концентрируются во внетропических областях (главным образом – в субтропиках) обоих полушарий, хотя в Южном, по ряду причин, сильнее, чем в Северном.
Реликтовый ареал таксона, распространенного во внетропических областях обоих полушарий, называют биполярным или амфитропическим. Ареал этот может в дальнейшем подвергаться различным модификациям. Упомянем в этой связи предложенную К.Ю. Еськовым (1984) модель «циркумпацифического домино», основанную на распространении современных и вымерших беспозвоночных и растений в Тихоокеанском регионе. Она предстваляет собой нечто вроде «периодической системы» транспацифических дизьюнкций: все их варианты рассматриваются как последовательные стадии сокращения единого биполярного ареала (рисунок 65). Редукция северной его «дольки» (рисунок 65, 6) и породит тот самый «гондванский ареал», которым оперирует викариантная биогеография. Обратная ситуация (редукция южной «дольки» – рисунок 65, 2) наблюдается заметно реже, однако существует и целый ряд таксонов, вымерших в Южном полушарии и сохранившихся в Северном (таковы, например, хвостатые амфибии и насекомые из отрядов верблюдок и тараканосверчков). Некоторые авторы связывают эту асимметрию просто с худшей палеонтологической изученностью Южных материков; и действительно, за последние годы, после открытия крупных местонахождений мезозойских насекомых в Австралии и Бразилии, число случаев документированного вымирания в Гондване «лавразийских» ныне таксонов возросло вдвое!
РИСУНОК 65. «Циркумпацифическое домино» – вероятный ход сокращения первоначально единого биполярного циркумпацифического ареала. I – группа присутствует в регионе, II – группа вымерла; а – внетропическая восточная Азия, б – запад Северной Америки, в – восток Северной Америки, г – внетропическая Австралия, д – Новая Зеландия, е – внетропическая Южная Америка. Примеры ареалов: 1 – пауки подотряда Hypochilomorphae, 2 – хвойные рода Tsuga, 3 – насекомые отряда Grylloblattida, 4 – гигантские саламандры Cryptobranchiidae, 5 – саламандры семейства Ambistomatidae, 6 – клопы подотряда Peloridiina, 7 – цветковые семейства Tetrachondraceae, 8 – цветковые семейства Eucryphiaceae, 9 – ручейники семейства Helicophiidae, 10 – хвойные семейства Taxodiaceae, 11 – насекомые-вислокрылки рода Protochauloides, 12 – пауки-птицеяды семейства Mecicobothriidae, 13 – лягушки семейства Leiopelmidae, 14 – цветковые семейства Lardisabalaceae, 15 – скорпионницы семейства Meropeidae.
Итак, ключевой проблемой для уоллесовской схемы оказывается механизм образования такого исходного ареала, из которого можно вывести все остальные: биполярного, биполярной пары «сестринских» таксонов (таковы, например, буки и нотофагусы), или же такого типа распространения, когда более продвинутые субтаксоны приурочены к тропикам, а более архаичные – к внетропическим областям. Необходимое для этого вымирание таксона на территории тропиков – с замещением его более продвинутыми формами или без такового – находит объяснение в двух альтернативных моделях: «экваториальной помпы» и «зональной стратификации».
«Экваториальная помпа» (термин принадлежит Ф. Дарлингтону) предполагает, что именно в тропических экосистемах с их стабильностью среды и высоким разнообразием идет постоянное и наиболее активное формообразование. При этом реликты «выдавливаются» (помпа!) и на север, и на юг, за счет конкуренции со стороны более молодых, продвинутых таксонов. Наиболее завершенный вид эта модель приобрела в работе С.В. Мейена (1987), основанной на палеоботаническом материале. Он показал, что в течение палеозоя почти все надродовые таксоны высших растений появлялись в экваториальном поясе в более древних стратиграфических слоях, чем за его пределами, и, следовательно, имеют экваториальное происхождение (так, доминирующие в Гондване и Ангариде глоссоптериевые и кордаиты первоначально возникли в тропиках Еврамерии – см. главу 9).
Именно тропический пояс является главной ареной макроэволюционных событий. Отсюда сформировавшиеся таксоны при потеплениях «растекаются» во внеэкваториальные широты (Мейен назвал это «фитоспредингом» – по аналогии со спредингом океанического дна), где при последующих похолоданиях они либо вымирают, либо эволюционируют в более узких (обычно внутриродовых) таксономических рамках; для внетропических областей на протяжении всей истории характерно длительное переживание архаических форм («внетропическое персистирование»). При этом процесс этот строго однонаправленный: вселения бореальных групп в тропики при похолоданиях не происходит никогда (рисунок 66). Причину повышенной интенсивности макроэволюции в тропиках Мейен видел в том, что здесь очень сильна биотическая конкуренция, но зато ослаблена неизбирательная абиотическая компонента отбора (холод, засухи, и т.д.). В результате естественный отбор менее жестко, чем в иных местах, выбраковывает крупные морфологические новации, давая шанс «перспективным монстрам» (термин Р. Гольдшмидта, 1940).
РИСУНОК 66. Филогения высших растений и географическая приуроченность основных филогенетических линий. 1 – нотальные (гондванские) внетропические области, 2 – экваториальные области, 3 – бореальные внетропические области, 4 – географическая ситуация неопределенна.
Альтернативный взгляд на природу амфитропических ареалов высказывал В.В. Жерихин (1978). Он основан на том, что в мезозое и раннем палеогене климатическая зональность в ее современном виде отсутствовала, и климат практически по всей планете был близок к нынешнему субтропическому. В конце палеогена произошла климатическая перестройка, в результате которой тепло стало распределяться по поверхности планеты менее равномерно, и на полюсах стало заметно холоднее, чем прежде, а на экваторе – жарче. В итоге единую «теплоумеренно-субтропическую» биоту Земли охватил процесс «зональной стратификации»; он заключался в том, что в экваториальных и приполярных широтах (где сменился климат) начались экосистемные перестройки, приведшие к достаточно массовому вымиранию мезозойских и раннепалеогеновых групп. Эти группы имели наилучшие шансы выжить в двух амфитропических «кольцах» с субтропическим и теплоумеренным климатом, господствовавшим ранее по всей планете – где они и поныне существуют в статусе реликтов (см. также главу 13).
Сравнение моделей «экваториальной помпы» и «зональной стратификации» позволяет прийти к достаточно неожиданному выводу о том, что они в действительности не альтернативны, а взаимодополнительны. Более того, К.Ю. Еськовым (1994) была предложена и обобщающая их концепция. Она состоит в том, что единый механизм мейеновского «фитоспрединга» может работать в различных режимах, диктуемых внешними (макроклиматическими) условиями, и «экваториальная помпа» есть просто один из этих рабочих режимов. Фитоспрединг понимается здесь как локализация макроэволюционных процессов (ведущих к возникновению новых таксонов) в тех районах Земли, где климатические условия наиболее благоприятны, и, соответственно, абиотическая компонента естественного отбора наиболее ослаблена; отсюда затем происходит однонаправленное «растекание» таксонов в районы с более жесткой абиотикой, где эти группы могут выживать и после своего вымирания в исходных районах. Фитоспрединг, таким образом, должен порождать целый ряд «градиентов архаичности»: гумидно-аридный, равнинно-высокогорный, океаническо-континентальный и т.д. Однако в эпохи резко выраженной широтной зональности климата (т.е. в криоэры) на всех них накладывается мощнейший температурный экваториально-полярный градиент, который низводит их до «градиентов второго порядка» и затушевывает их существование.
Именно в эти периоды фитоспрединг и работает в режиме «экваториальной помпы». В эпохи же с ослабленной широтной зональностью (термоэры) центры формообразования не локализованы в экваториальной зоне, а более диффузно распределены по всей планете. Число нарушений установленной Мейеном закономерности в стратиграфическом распределении членов таксона (в низких широтах они появляются в более древних слоях, чем в высоких) должно заметно возрастать в азональные эпохи – и анализ распределения позднепалеозойских и мезозойских групп насекомых подтвердил, что в целом это действительно так. Процесс же перехода биоты Земли от «полиградиентного» состояния (азональные эпохи) к «моноградиентному» (зональные эпохи) происходит в соответствии с моделью «зональной стратификации». Итак, фитоспрединг – это универсальный механизм флоро– и фауногенеза; он имеет три режима работы, определяемых глобальным термическим градиентом, и режимы эти формируют замкнутый трехчленный цикл: «экваториальная помпа» (поздний палеозой, криоэра) – «диффузные центры формообразования» (мезозой-ранний палеоген, термоэра) – «зональная стратификация» (поздний палеоген) – снова «экваториальная помпа» (неоген, криоэра), и т.д.
Рекомендуемая литература
1. Художественная
А. Конан-Дойл «Затерянный мир»В.А.Обручев «Плутония»; «Земля Санникова»
Ж. Рони Старший «Борьба за огонь»; «Пещерный лев»
М. Крайтон «Парк юрского периода»
2. Научно-популярная
А. Азимов .....................С. Уэда «Новый взгляд на Землю»
А.С. Монин «Популярная история Земли»
Дж. Имбри, К.П. Имбри «Тайны ледниковых эпох»
Юл. Медведев «Судеб посланник» (в книге «Бросая вызов»)
И.А. Ефремов «Дорога ветров»
Дж. Смит «Старина четвероног»
А.К. Рождественский «На поиски динозавров в Гоби»
И. Аугуста, З. Буриан «По путям развития жизни»; «Летающие ящеры и древние птицы»; «Ящеры древних морей»; «Гиганты суши»; «Книга о мамонтах»; «Жизнь древнего человека»
К.Л. Фентон, М.А. Фентон «Каменная книга. Летопись доисторической жизни»
Ю.А. Орлов «О мире древних животных»
Д. Эттенборо «Жизнь на Земле»
И. Яковлева, В. Яковлев «По следам минувшего»
М.Ф. Ивахненко, В.А. Корабельников «Живое прошлое Земли»
А. Гангнус «Через горы времени»
Г.А. Заварзин «Бактерии и состав атмосферы Земли»
И.Н. Крылов «На заре жизни»
А.Ю. Розанов «Что произошло 600 миллионов лет назад»
С.В. Мейен «Из истории растительных династий»; «Следы трав индейских»
Н. Эйдельман «Ищу предка»
Р. Фоули «Еще один великолепный вид»
В.А. Красилов «Нерешенные вопросы теории эволюции»
3. Научная и учебная
А.С. Монин, ......Шишков «История климата»
Р. Кэрролл «Палеонтология и эволюция позвоночных» (3 тома)
В.В. Друщиц «Палеонтология беспозвоночных»
С.В. Мейен «Основы палеоботаники»; «Историческое развитие класса насекомых», Труды ПИН АН СССР, т.175
В.В. Жерихин «Развитие и смена меловых и кайнозойских фаунистических комплексов» (Общая часть)
Ф. Дарлингтон «Зоогеография» Дж. Симпсон «Великолепная изоляция»
С.М. Разумовский «Закономерности динамики фитоценозов»
О.Г. Кусакин, А.Л. Дроздов «Филема органического мира»
В.А. Красилов «Происхождение и ранняя эволюция цветковых растений»; «Эволюция и биостратиграфия»
Л.П. Татаринов «Очерки по теории эволюции»
Т. де Шарден «Феномен человека»
Э. Шредингер «Что такое жизнь с точки зрения физика»
И. Пригожин, И. Стенгерс «Порядок из хаоса»
Словарь терминов
автотрофы – организмы, способные самостоятельно синтезировать органические вещества из неорганических, используя энергию солнца (фототрофы – см.) или окислительно-восстановительных химических реакций (хемотрофы – см.).
адаптация – приспособление строения и функций организмов к условиям их существования.
адекватный – вполне соответствующий.
амфибиотический – связанный с жизнью как в воде, так и на суше.
аналогия – сходство в каком-либо отношении между предметами, явлениями или понятиями.
анаэробы – организмы, живущие в анаэробных условиях: при отсутствии свободного кислорода. Аэробами являются часть микроорганизмов.
анемофилия – приспособленность цветков растений к опылению при помощи ветра.
антициклон – крупномасштабная область высокого атмосферного давления с циркуляцией воздуха по часовой стрелке в северном полушарии и и против часовой стрелки – в южном, со слабыми ветрами и малооблачной погодой.
антофилия – приспособленность различных животных к связям с цветками.
аспект – точка зрения, с которой рассматривается предмет, явление или понятие.
аридный климат – климат, характеризующийся преобладанием испарения над атмосферными осадками, что создает дефицит влаги.
архаичность – черты, свидетельствующие о древности организма или биоты.
аэробы – организмы, живущие в аэробных условиях: при наличии свободного кислорода. Аэробами являются животные, растения и часть микроорганизмов.
бентос – совокупность живых организмов, обитающих на дне водоемов (на грунте и в грунте).
биогены (биогенные элементы) – химические элементы, непременно входящие в состав живых организмов (углерод, кислород, азот, сера и т.д.).
биосфера – область распространения жизни на Земле; состав, структура и энергетика биосферы определяется главным образом прошлой и/или современной деятельностью живых организмов.
биота – исторически сложившаяся совокупность живых организмов, объединенных общей областью распространения и временем существования.
брожение – процесс ферментативного неполного окисления углеводов, происходящий без участия свободного кислорода; конечным продуктом брожения могут быть спирт или органические кислоты (молочная, уксусная). Составляет основу обмена веществ многих бактерий.
буферные растворы – водные растворы слабой кислоты и ее соли или слабого основания и его соли; обладают способностью сохранять определенный уровень кислотности (концентрации водородных ионов) при добавлении небольших количеств сильной кислоты или щелочи. Пример буферного раствора – существующий в морской воде карбонатно-бикарбонатный буфер (угольная кислота с углекислый кальцием).
восстановительная атмосфера – атмосфера, в составе которой отсутствуют газы-окислители: молекулярный кислород, галогены и т.п.
геохимический цикл – путь миграции атомов некоего химического элемента в оболочках Земли (атмосфере, гидросфере, литосфере и биосфере) в ходе природных (в том числе – биологических) процессов.
гетеротрофы – организмы, неспособные самостоятельно синтезировать органические вещества из неорганических и использующие органику, созданную другими живыми существами. Животные, грибы и часть бактерий.
гипертонический раствор – раствор, осмотическое давление (см.) которого выше осмотического давления внутриклеточного содержимого.
гипотеза – научное предположение, выдвигаемое для объяснения какого-либо явления и требующее проверки на опыте и/или теоретического обоснования для того, чтобы стать достоверной научной теорией (см.).
гипотонический раствор – раствор, осмотическое давление (см.) которого ниже осмотического давления внутриклеточного содержимого.
гликолиз – анаэробное расщепление углеводов в животном организме с образованием молочной кислоты.
гомункулус – по представлениям средневековых алхимиков – существо, подобное человеку, которое можно получить искусственно, «в колбе».
градиент – мера возрастания или убывания в пространстве какой-либо физической величины на единицу расстояния.
гумус – органический компонент почвы, образующийся в результате биохимических превращений растительных и животных остатков.
гумидный климат – климат, характеризующийся преобладанием атмосферных осадков в виде воды над испарением, что ведет к избыточному увлажнению.
давление света – давление, производимое светом на тела, отражающие или поглощающие свет. Сила давления света на малые частицы в космических процессах того же порядка, что и сила тяжести.
дегазация магмы – удаление из расплавленной магмы растворенных в ней газов, происходящее как в канале вулкана, так и на поверхности Земли.
детерминизм – философская концепция, признающая объективную закономерность и причинную обусловленность всех явлений природы и общества.
детритная цепь – трофическая цепь (см.), которая начинается от мертвого органического вещества, идет к микроорганизмам, которые им питаются, а затем к детритофагам (см.) и их хищникам.
детритофаг – потребитель мертвой органики – трупов, остатков растений, экскрементов.
дисперсный – дисперсная система: вещество в виде малых частиц (дисперсная фаза) вместе с той средой (дисперсионная среда), в которой они распределены.
дифференциация – разделение (структурное или функциональное) целого на части.
диффузия – проникновение молекул одного вещества (жидкости, газа, твердого тела) в другое при непосредственном их соприкосновении, обусловленное тепловым движением молекул.
дыхание (в биохимическом смысле) – процесс полного окисления углеводов до углекислого газа и воды, происходящий в присутствии кислорода. Основа обмена веществ животных и растений; имеется и у некоторых бактерий.
естественный отбор – стохастический (вероятностный) процесс преимущественного переживания и размножения более приспособленных организмов и элиминации (выбраковки) менее приспособленных.
западный перенос (в атмосфере) – отклонение воздушных течений от первоначального направления: в Северном полушарии вправо, в Южном – влево; возникает под действием отклоняющей силы вращения Земли (криолисова сила).
зоопланктоном – совокупность планктонных (см.) животных.
изотопы – атомы одного и того же химического элемента, которые содержат одинаковое число протонов, но разное число нейтронов. Обладают одинаковыми химическими, но разными физическими свойствами. Существуют устойчивые (стабильные) и неустойчивые (радиоактивные) изотопы.
интерпретация – истолкование, разъяснение смысла чего-либо.
информация – количественная мера устранения неопределенности (энтропии), мера организации системы.
информационный шум – ..................................................................
каверна – пустота, полое пространство, образовавшееся в массиве горных пород.
катализатор – вещество, изменяющее скорость химической реакции.
квант света (фотон) – частица – элементарный носитель свойств электромагнитного поля.
квантовая механика – раздел физики, изучающий законы движения и взаимодействия частиц очень малой массы (например, электронов, протонов и т.д.), лежащие в основе современного понимания строения материи на уровне атомов.
Кельвина шкала – абсолютная шкала температур, не зависящая от свойств термометрического вещества (начало отсчета – абсолютный нуль температуры).
коадаптация – взаимное приспособление друг к другу в процессе совместной эволюции организмов, входящих в одну экосистему.
конвергенция – приобретение в ходе эволюции сходного строения и функций неродственными организмами вследствие их приспособления к сходным условиям обитания.
кора выветривания – комплекс горных пород, возникающих путем преобразования в континентальных условиях магматических и осадочных пород под действием выветривания. Многие коры выветривания возникли в результате биогенных процессов.
конгломерат – сцементированная обломочная горная порода, состоящая преимущественно из гальки разного состава; образуется в результате размыва и переотложения более древних пород.
конечный водоем стока – море или бессточное озеро.
консументы – второй и более высокие трофические уровни (см.) экосистемы: растительноядные животные, хищники, вторичные хищники.
конус выноса – аккумулятивная форма рельефа в виде полуконуса; возникает на месте крутого перелома продольного профиля реки с крутого на более пологий, в результате чего поток теряет силу и несомые им твердые частицы начинают отлагаться.
красноцветы – собирательное название осадочных пород различного происхождения, имеющих красноватую окраску за счет содержащихся в них окислов железа.
Кребса цикл (цикл трикарбоновых кислот) – одна из стадий процесса дыхания (см.). Сложный замкнутый круг превращений, по ходу которых молекула трехатомной пировиноградной кислоты превращается в три молекулы углекислого газа с выделением большого количества энергии.
кремнезем – двуокись кремния.
латентный – скрытый, внешне не проявляющийся.
литораль – прибрежная часть морского дна, затопляемая в прилив и обсыхающая в отлив.
локомоция – совокупность координированных (согласованных) движений, при помощи которых животное перемещается в пространстве.
Лотки-Вольтерры модель – система двух диференциальных уравнений, используемых для моделирования динамики численности в изолированной паре взаимодействующих между собой популяций (хищник-жертва, паразит-хозяин и т.п.).
адаптация – приспособление строения и функций организмов к условиям их существования.
адекватный – вполне соответствующий.
амфибиотический – связанный с жизнью как в воде, так и на суше.
аналогия – сходство в каком-либо отношении между предметами, явлениями или понятиями.
анаэробы – организмы, живущие в анаэробных условиях: при отсутствии свободного кислорода. Аэробами являются часть микроорганизмов.
анемофилия – приспособленность цветков растений к опылению при помощи ветра.
антициклон – крупномасштабная область высокого атмосферного давления с циркуляцией воздуха по часовой стрелке в северном полушарии и и против часовой стрелки – в южном, со слабыми ветрами и малооблачной погодой.
антофилия – приспособленность различных животных к связям с цветками.
аспект – точка зрения, с которой рассматривается предмет, явление или понятие.
аридный климат – климат, характеризующийся преобладанием испарения над атмосферными осадками, что создает дефицит влаги.
архаичность – черты, свидетельствующие о древности организма или биоты.
аэробы – организмы, живущие в аэробных условиях: при наличии свободного кислорода. Аэробами являются животные, растения и часть микроорганизмов.
бентос – совокупность живых организмов, обитающих на дне водоемов (на грунте и в грунте).
биогены (биогенные элементы) – химические элементы, непременно входящие в состав живых организмов (углерод, кислород, азот, сера и т.д.).
биосфера – область распространения жизни на Земле; состав, структура и энергетика биосферы определяется главным образом прошлой и/или современной деятельностью живых организмов.
биота – исторически сложившаяся совокупность живых организмов, объединенных общей областью распространения и временем существования.
брожение – процесс ферментативного неполного окисления углеводов, происходящий без участия свободного кислорода; конечным продуктом брожения могут быть спирт или органические кислоты (молочная, уксусная). Составляет основу обмена веществ многих бактерий.
буферные растворы – водные растворы слабой кислоты и ее соли или слабого основания и его соли; обладают способностью сохранять определенный уровень кислотности (концентрации водородных ионов) при добавлении небольших количеств сильной кислоты или щелочи. Пример буферного раствора – существующий в морской воде карбонатно-бикарбонатный буфер (угольная кислота с углекислый кальцием).
восстановительная атмосфера – атмосфера, в составе которой отсутствуют газы-окислители: молекулярный кислород, галогены и т.п.
геохимический цикл – путь миграции атомов некоего химического элемента в оболочках Земли (атмосфере, гидросфере, литосфере и биосфере) в ходе природных (в том числе – биологических) процессов.
гетеротрофы – организмы, неспособные самостоятельно синтезировать органические вещества из неорганических и использующие органику, созданную другими живыми существами. Животные, грибы и часть бактерий.
гипертонический раствор – раствор, осмотическое давление (см.) которого выше осмотического давления внутриклеточного содержимого.
гипотеза – научное предположение, выдвигаемое для объяснения какого-либо явления и требующее проверки на опыте и/или теоретического обоснования для того, чтобы стать достоверной научной теорией (см.).
гипотонический раствор – раствор, осмотическое давление (см.) которого ниже осмотического давления внутриклеточного содержимого.
гликолиз – анаэробное расщепление углеводов в животном организме с образованием молочной кислоты.
гомункулус – по представлениям средневековых алхимиков – существо, подобное человеку, которое можно получить искусственно, «в колбе».
градиент – мера возрастания или убывания в пространстве какой-либо физической величины на единицу расстояния.
гумус – органический компонент почвы, образующийся в результате биохимических превращений растительных и животных остатков.
гумидный климат – климат, характеризующийся преобладанием атмосферных осадков в виде воды над испарением, что ведет к избыточному увлажнению.
давление света – давление, производимое светом на тела, отражающие или поглощающие свет. Сила давления света на малые частицы в космических процессах того же порядка, что и сила тяжести.
дегазация магмы – удаление из расплавленной магмы растворенных в ней газов, происходящее как в канале вулкана, так и на поверхности Земли.
детерминизм – философская концепция, признающая объективную закономерность и причинную обусловленность всех явлений природы и общества.
детритная цепь – трофическая цепь (см.), которая начинается от мертвого органического вещества, идет к микроорганизмам, которые им питаются, а затем к детритофагам (см.) и их хищникам.
детритофаг – потребитель мертвой органики – трупов, остатков растений, экскрементов.
дисперсный – дисперсная система: вещество в виде малых частиц (дисперсная фаза) вместе с той средой (дисперсионная среда), в которой они распределены.
дифференциация – разделение (структурное или функциональное) целого на части.
диффузия – проникновение молекул одного вещества (жидкости, газа, твердого тела) в другое при непосредственном их соприкосновении, обусловленное тепловым движением молекул.
дыхание (в биохимическом смысле) – процесс полного окисления углеводов до углекислого газа и воды, происходящий в присутствии кислорода. Основа обмена веществ животных и растений; имеется и у некоторых бактерий.
естественный отбор – стохастический (вероятностный) процесс преимущественного переживания и размножения более приспособленных организмов и элиминации (выбраковки) менее приспособленных.
западный перенос (в атмосфере) – отклонение воздушных течений от первоначального направления: в Северном полушарии вправо, в Южном – влево; возникает под действием отклоняющей силы вращения Земли (криолисова сила).
зоопланктоном – совокупность планктонных (см.) животных.
изотопы – атомы одного и того же химического элемента, которые содержат одинаковое число протонов, но разное число нейтронов. Обладают одинаковыми химическими, но разными физическими свойствами. Существуют устойчивые (стабильные) и неустойчивые (радиоактивные) изотопы.
интерпретация – истолкование, разъяснение смысла чего-либо.
информация – количественная мера устранения неопределенности (энтропии), мера организации системы.
информационный шум – ..................................................................
каверна – пустота, полое пространство, образовавшееся в массиве горных пород.
катализатор – вещество, изменяющее скорость химической реакции.
квант света (фотон) – частица – элементарный носитель свойств электромагнитного поля.
квантовая механика – раздел физики, изучающий законы движения и взаимодействия частиц очень малой массы (например, электронов, протонов и т.д.), лежащие в основе современного понимания строения материи на уровне атомов.
Кельвина шкала – абсолютная шкала температур, не зависящая от свойств термометрического вещества (начало отсчета – абсолютный нуль температуры).
коадаптация – взаимное приспособление друг к другу в процессе совместной эволюции организмов, входящих в одну экосистему.
конвергенция – приобретение в ходе эволюции сходного строения и функций неродственными организмами вследствие их приспособления к сходным условиям обитания.
кора выветривания – комплекс горных пород, возникающих путем преобразования в континентальных условиях магматических и осадочных пород под действием выветривания. Многие коры выветривания возникли в результате биогенных процессов.
конгломерат – сцементированная обломочная горная порода, состоящая преимущественно из гальки разного состава; образуется в результате размыва и переотложения более древних пород.
конечный водоем стока – море или бессточное озеро.
консументы – второй и более высокие трофические уровни (см.) экосистемы: растительноядные животные, хищники, вторичные хищники.
конус выноса – аккумулятивная форма рельефа в виде полуконуса; возникает на месте крутого перелома продольного профиля реки с крутого на более пологий, в результате чего поток теряет силу и несомые им твердые частицы начинают отлагаться.
красноцветы – собирательное название осадочных пород различного происхождения, имеющих красноватую окраску за счет содержащихся в них окислов железа.
Кребса цикл (цикл трикарбоновых кислот) – одна из стадий процесса дыхания (см.). Сложный замкнутый круг превращений, по ходу которых молекула трехатомной пировиноградной кислоты превращается в три молекулы углекислого газа с выделением большого количества энергии.
кремнезем – двуокись кремния.
латентный – скрытый, внешне не проявляющийся.
литораль – прибрежная часть морского дна, затопляемая в прилив и обсыхающая в отлив.
локомоция – совокупность координированных (согласованных) движений, при помощи которых животное перемещается в пространстве.
Лотки-Вольтерры модель – система двух диференциальных уравнений, используемых для моделирования динамики численности в изолированной паре взаимодействующих между собой популяций (хищник-жертва, паразит-хозяин и т.п.).