Страница:
Складывающиеся на совершенно новой основе раннекайнофитные сообщества первоначально были несбалансированными и неустойчивыми. Слабость биоценотической регуляции и обилие незанятых экологических ниш вели к очень быстрой, взрывной, эволюции различных групп организмов (из насекомых здесь можно отметить высших бабочек и мух, паразитических перепончатокрылых, а также общественных насекомых – муравьев и термитов), по мере же насыщения биоценозов темпы эволюции снижались. Сложные трофические цепи в новых кайнофитных сообществах сформировались очень быстро (листья, поврежденные минирующими бабочками, сами эти бабочки и их высокоспециализированные паразиты из числа наездников Braconidae появляются в летописи почти одновременно, в сеномане) и в дальнейшем менялись мало. «По экологическому облику и систематическому составу , – заключает Жерихин (1980), – позднемеловые фауны значительно сильнее отличаются от раннемеловых, чем от кайнозойских, и могут рассматриваться как ранний, первоначально сильно обедненный, вариант этих последних. В сущности, по набору жизненных форм они имеют не так уж много отличий от современных».
Если вернуться к истокам среднемелового кризиса – самому возникновению покрытосеменных, – то здесь обнаруживается еще один интересный аспект, на который обратил внимание А.Г. Пономаренко (1997). Дело в том, что, по данным последних лет, серьезные эволюционные изменения среди насекомых начинаются несколько раньше широкого распространения покрытосеменных – с конца юры. Это, казалось бы, противоречит гипотезе о причинной связи между этими явлениями – но только на первый взгляд. Именно в это время начинается очередная «...зация» – ангиоспермизация голосеменных, и широко распространяются проангиоспермы (термин В.А. Красилова, 1989) – голосеменные с отдельными чертами цветковых (таковы, например, гнетовое Eoantha с крупными, явно энтомофильными «цветами», или беннетит Manlaia, имитирующий осоку – см. рисунок 52). «С точки зрения экосистемы» точное систематическое положение всех этих проангиоспермов не столь уж важно – все они могут в некоторой степени играть экологическую роль настоящих покрытосеменных.
Неудивительно, что параллельно с энтомофильными голосеменными появляются антофильные насекомые; имитаторам цветковых в растительном мире сопутствуют столь же многочисленные имитаторы бабочек среди насекомых (таковы, например, Calligrammatidae из сетчатокрылых, или Aneuretopsychidae из скорпионниц – см. рисунок 52). Важнее же всего то, что здесь складывается система с положительной обратной связью: наличие у некого беннетита комплекса опыляющих его насекомых стимулирует появление энтомофилии у его соседей-голосеменных, что, в свою очередь, расширяет экологическую нишу этих насекомых-опылителей, и т.д. Каждый шаг на этом пути увеличивает вероятность следующего и делает появление покрытосеменных (т.е. растений с полным синдромом ангиоспермных признаков) совершенно неизбежным; какая же конкретно группа голосеменных даст начало «настоящим цветковым» (равно как – какие именно из мекоптероидных насекомых породят «настоящих бабочек»), вообще-то говоря, не столь уж важно. Именно эти взаимно индуцируемые процессы Пономаренко и назвал «ангиоспермизацией мира».
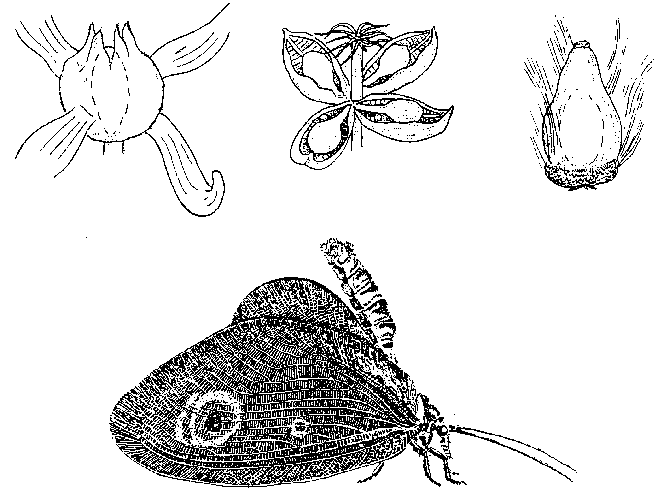
РИСУНОК 52. Голосеменные-имитаторы цветковых и насекомые-имитаторы бабочек. (а) – гнетовое Dinophyton, (б) – гнетовое Eoantha, (в) – беннетит Baisia, (г) – сетчатокрылое Kalligrammatidae.
«Ангиоспермизация мира» захватывает не только насекомых, но и все прочие компоненты континентальных экосистем. С начала мела на суше стремительно распространяются птицы (это особенно хорошо видно по укрупнению птерозавров – те полностью уходят в нишу крупных парителей) и териевые млекопитающие; в пресных водоемах настоящие костистые рыбы вытесняют ганоидов. После апт-альбского кризиса мир обретает вполне знакомый нам облик; правда, в нем наличествуют динозавры (и не просто «наличествуют» – они вполне процветает на протяжении большей части позднего мела), однако судьба этой группы была рассмотрена нами выше (глава 11), и нет оснований напрямую связывать ее вымирание с драматическими изменениями в морской биоте на границе мела и кайнозоя. Таким образом, мезозойско-кайнозойская смена на суше произошла много раньше, чем в море.
Ну, а каковы же всё-таки причины позднемелового «Великого вымирания»? Если не падение астероида и прочие «импакты» – то что же? Давайте прежде всего попытаемся найти какие-нибудь общие черты для этого события и для другого морского вымирания – пермско-триасового. Оно было самым крупным в истории Земли – тогда в морских группах вымерло 50% семейств, 70% родов и более 90% видов (для сравнения: при мел-кайнозойском вымирании исчезло менее четверти семейств). Помимо прямого исчезновения множества палеозойских групп (трилобиты, палеозойские кораллы – ругозы и табуляты) произошла радикальная смена структуры морских экосистем (роль основных рифостроителей перешла от мшанок к кораллам, донных фильтраторов – от брахиопод к двустворчатым моллюскам, и т.п.). Причины этой грандиозной фаунистической смены остаются загадкой; стандартные объяснения вроде «глобального катастрофического вулканизма» или морской регрессии, резко сократившей площадь шельфов (основной зоны обитания морских организмов) производят впечатление явно придуманных «за неимением лучшего». Мне лично наиболее остроумной и логичной (хотя и не бесспорной) представляется гипотеза Х. Таппан – специалиста по фитопланктону.
В 1986 году Таппан опубликовала работу под названием «Фитопланктон: за солью на планетарном столе (Phytoplancton: below the salt at the global table)»; название это требует пояснения. В средневековом английском замке представители всех социальных слоев ели за общим длинным столом. Дворяне, рыцари и почетные гости сидели во главе, и большая солонка, стоящая примерно посередине стола, как бы отделяла их места от мест слуг, солдат и «гостей второй свежести», занимающих другой его конец; места эти называли, соответственно, «перед солью» и «за солью» («above and below the salt»). Кушанья, как легко догадаться, подавали сперва на тот конец, что «перед солью», а «за соль» доходило – уж что останется...
Так вот, по мысли Таппан, наземная биота планеты всегда занимает место «во главе стола», а океанская – оказывается «за солью». Необходимые для жизнедеятельности фитопланктона биогены (прежде всего азот и фосфор) поступают с суши, за счет эрозии; наземная растительность, между тем, стремится подавить эрозию и оставить как можно больше биогенов для собственного потребления (помните – замыкание геохимических циклов?). Таким образом, все заметные эволюционные успехи наземной растительности замедляют поступление биогенов в океан, что со временем вызывает вымирание соответствующих групп фитопланктона; фитопланктон же – основа первого трофического уровня любых морских экосистем, так что дальше разваливается вся цепь. Следует помнить, однако, что инерция такой системы огромна, и между «прогрессивным изменением» в наземной растительности и вызванным им «фитопланктонным шоком» в океане проходит ощутимый срок.
Подойдя с этих позиций к пермо-триасовому и мел-палеогеновому морским вымираниям, мы увидим, что в поздней перми и в позднем мелу действительно произошли два серьезных изменения в структуре наземной растительности, прямо влияющие на регуляцию потока биогенов с суши. В перми высшая растительность впервые начинает заселять водоразделы, тормозя их прежде беспрепятственный размыв; этому способствует возникновение засухоустойчивых голосеменных (типа вольциевых) с пикноксилической древесиной и мелкими, сильно склеротизованными листьями (а также, как мы помним, появление насекомых с водными личинками – см. главу 9). В позднем мелу появляется трава, образующая дернину – наиболее эффективный из известных подавителей эрозии.
Что же касается импактов, то они, по всей видимсти, не представляют угрозы для сбалансированных, «здоровых» экосистем, но вот если те уже находятся в состоянии кризиса (вроде планктона, столкнувшегося с серьезными глобальными изменениями в порядке поступления биогенов), то кратковременная «астероидная зима» вполне способна сыграть роль «последней соломинки, ломающей спину верблюда». Тут возможна аналогия с выпадением соли из пересыщенного раствора: никому ведь не придет в голову считать причиной этого процесса упавший в сосуд кристаллик, непосредственно инициировавший его начало.
13. Кайнозой: наступление криоэры. Новые типы сообществ – тропические леса и травяные биомы. Эволюция млекопитающих и появление человека.
Если вернуться к истокам среднемелового кризиса – самому возникновению покрытосеменных, – то здесь обнаруживается еще один интересный аспект, на который обратил внимание А.Г. Пономаренко (1997). Дело в том, что, по данным последних лет, серьезные эволюционные изменения среди насекомых начинаются несколько раньше широкого распространения покрытосеменных – с конца юры. Это, казалось бы, противоречит гипотезе о причинной связи между этими явлениями – но только на первый взгляд. Именно в это время начинается очередная «...зация» – ангиоспермизация голосеменных, и широко распространяются проангиоспермы (термин В.А. Красилова, 1989) – голосеменные с отдельными чертами цветковых (таковы, например, гнетовое Eoantha с крупными, явно энтомофильными «цветами», или беннетит Manlaia, имитирующий осоку – см. рисунок 52). «С точки зрения экосистемы» точное систематическое положение всех этих проангиоспермов не столь уж важно – все они могут в некоторой степени играть экологическую роль настоящих покрытосеменных.
Неудивительно, что параллельно с энтомофильными голосеменными появляются антофильные насекомые; имитаторам цветковых в растительном мире сопутствуют столь же многочисленные имитаторы бабочек среди насекомых (таковы, например, Calligrammatidae из сетчатокрылых, или Aneuretopsychidae из скорпионниц – см. рисунок 52). Важнее же всего то, что здесь складывается система с положительной обратной связью: наличие у некого беннетита комплекса опыляющих его насекомых стимулирует появление энтомофилии у его соседей-голосеменных, что, в свою очередь, расширяет экологическую нишу этих насекомых-опылителей, и т.д. Каждый шаг на этом пути увеличивает вероятность следующего и делает появление покрытосеменных (т.е. растений с полным синдромом ангиоспермных признаков) совершенно неизбежным; какая же конкретно группа голосеменных даст начало «настоящим цветковым» (равно как – какие именно из мекоптероидных насекомых породят «настоящих бабочек»), вообще-то говоря, не столь уж важно. Именно эти взаимно индуцируемые процессы Пономаренко и назвал «ангиоспермизацией мира».
РИСУНОК 52. Голосеменные-имитаторы цветковых и насекомые-имитаторы бабочек. (а) – гнетовое Dinophyton, (б) – гнетовое Eoantha, (в) – беннетит Baisia, (г) – сетчатокрылое Kalligrammatidae.
«Ангиоспермизация мира» захватывает не только насекомых, но и все прочие компоненты континентальных экосистем. С начала мела на суше стремительно распространяются птицы (это особенно хорошо видно по укрупнению птерозавров – те полностью уходят в нишу крупных парителей) и териевые млекопитающие; в пресных водоемах настоящие костистые рыбы вытесняют ганоидов. После апт-альбского кризиса мир обретает вполне знакомый нам облик; правда, в нем наличествуют динозавры (и не просто «наличествуют» – они вполне процветает на протяжении большей части позднего мела), однако судьба этой группы была рассмотрена нами выше (глава 11), и нет оснований напрямую связывать ее вымирание с драматическими изменениями в морской биоте на границе мела и кайнозоя. Таким образом, мезозойско-кайнозойская смена на суше произошла много раньше, чем в море.
Ну, а каковы же всё-таки причины позднемелового «Великого вымирания»? Если не падение астероида и прочие «импакты» – то что же? Давайте прежде всего попытаемся найти какие-нибудь общие черты для этого события и для другого морского вымирания – пермско-триасового. Оно было самым крупным в истории Земли – тогда в морских группах вымерло 50% семейств, 70% родов и более 90% видов (для сравнения: при мел-кайнозойском вымирании исчезло менее четверти семейств). Помимо прямого исчезновения множества палеозойских групп (трилобиты, палеозойские кораллы – ругозы и табуляты) произошла радикальная смена структуры морских экосистем (роль основных рифостроителей перешла от мшанок к кораллам, донных фильтраторов – от брахиопод к двустворчатым моллюскам, и т.п.). Причины этой грандиозной фаунистической смены остаются загадкой; стандартные объяснения вроде «глобального катастрофического вулканизма» или морской регрессии, резко сократившей площадь шельфов (основной зоны обитания морских организмов) производят впечатление явно придуманных «за неимением лучшего». Мне лично наиболее остроумной и логичной (хотя и не бесспорной) представляется гипотеза Х. Таппан – специалиста по фитопланктону.
В 1986 году Таппан опубликовала работу под названием «Фитопланктон: за солью на планетарном столе (Phytoplancton: below the salt at the global table)»; название это требует пояснения. В средневековом английском замке представители всех социальных слоев ели за общим длинным столом. Дворяне, рыцари и почетные гости сидели во главе, и большая солонка, стоящая примерно посередине стола, как бы отделяла их места от мест слуг, солдат и «гостей второй свежести», занимающих другой его конец; места эти называли, соответственно, «перед солью» и «за солью» («above and below the salt»). Кушанья, как легко догадаться, подавали сперва на тот конец, что «перед солью», а «за соль» доходило – уж что останется...
Так вот, по мысли Таппан, наземная биота планеты всегда занимает место «во главе стола», а океанская – оказывается «за солью». Необходимые для жизнедеятельности фитопланктона биогены (прежде всего азот и фосфор) поступают с суши, за счет эрозии; наземная растительность, между тем, стремится подавить эрозию и оставить как можно больше биогенов для собственного потребления (помните – замыкание геохимических циклов?). Таким образом, все заметные эволюционные успехи наземной растительности замедляют поступление биогенов в океан, что со временем вызывает вымирание соответствующих групп фитопланктона; фитопланктон же – основа первого трофического уровня любых морских экосистем, так что дальше разваливается вся цепь. Следует помнить, однако, что инерция такой системы огромна, и между «прогрессивным изменением» в наземной растительности и вызванным им «фитопланктонным шоком» в океане проходит ощутимый срок.
Подойдя с этих позиций к пермо-триасовому и мел-палеогеновому морским вымираниям, мы увидим, что в поздней перми и в позднем мелу действительно произошли два серьезных изменения в структуре наземной растительности, прямо влияющие на регуляцию потока биогенов с суши. В перми высшая растительность впервые начинает заселять водоразделы, тормозя их прежде беспрепятственный размыв; этому способствует возникновение засухоустойчивых голосеменных (типа вольциевых) с пикноксилической древесиной и мелкими, сильно склеротизованными листьями (а также, как мы помним, появление насекомых с водными личинками – см. главу 9). В позднем мелу появляется трава, образующая дернину – наиболее эффективный из известных подавителей эрозии.
Что же касается импактов, то они, по всей видимсти, не представляют угрозы для сбалансированных, «здоровых» экосистем, но вот если те уже находятся в состоянии кризиса (вроде планктона, столкнувшегося с серьезными глобальными изменениями в порядке поступления биогенов), то кратковременная «астероидная зима» вполне способна сыграть роль «последней соломинки, ломающей спину верблюда». Тут возможна аналогия с выпадением соли из пересыщенного раствора: никому ведь не придет в голову считать причиной этого процесса упавший в сосуд кристаллик, непосредственно инициировавший его начало.
13. Кайнозой: наступление криоэры. Новые типы сообществ – тропические леса и травяные биомы. Эволюция млекопитающих и появление человека.
Мезозой, как мы помним из главы 9, был термоэрой и отличался «райским» климатом: теплым и выровненным по всей Земле, с ослабленным температурным градиентом между тропиками и полюсом. В рамках термоэр неоднократно происходили свои похолодания (например, в маастрихте), однако масштабы даже крупнейших из них не идут ни в какое сравнение с ледниковыми эпохами криоэр. В самом начале кайнозоя (в палеоцене и эоцене) климатическая ситуация оставалась еще прежней, мезозойской, а затем, в олигоцене, начались изменения, приведшие в конце концов к... так и тянет сказать: «к тому самому климатическому безобразию, в каковом мы теперь имеем счастье прозябать (в буквальном смысле этого слова)». Одной из главных причин этих изменений считают перестройку глобальной системы морских течений, вызванную дрейфом континентов, что затруднило теплообмен между полярными и экваториальными бассейнами.
Согласно моделям климатологов, наиболее интенсивный теплообмен будет достигнут при таком взаиморасположении материков и полюсов, когда планета имеет свободные от суши полюса и экватор (вдоль которого складывалась бы под действием кориолисовых сил единая экваториальная циркуляция). Меридионально развернутые материки, расположенные в средних широтах, отводят рукава экваториальной циркуляции в высокие широты, откуда вдоль противоположных их берегов возвращается в экваториальную зону холодная вода из полярных бассейнов – возвращается сразу, не застаиваясь в полярных циркуляциях. Как легко видеть (рисунок 53, а), мезозойская ситуация была в этом смысле близка к идеальной.
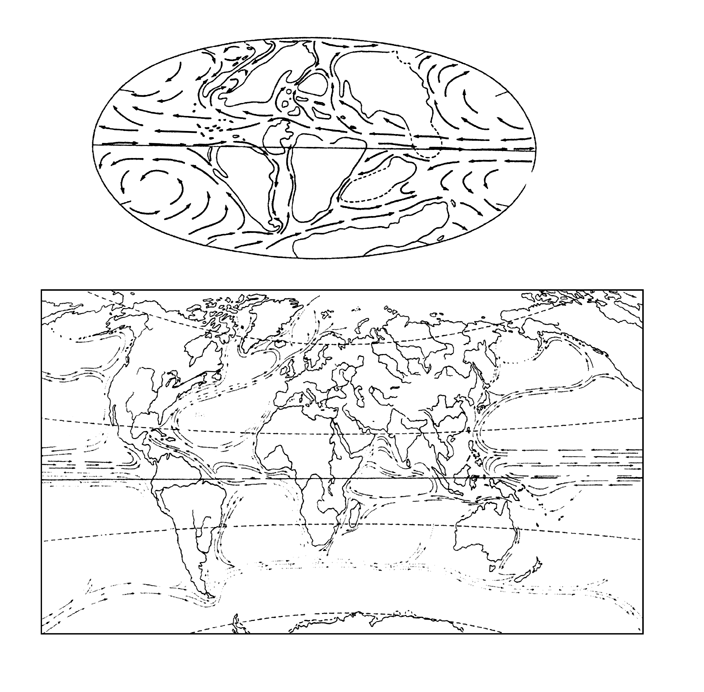
РИСУНОК 53. Материки и морские течения. Мезозойское (средний мел) (а) и современное (б) расположение материков.
В кайнозое картина становится близка к обратной, и причиною тому – перемещения материков, на которые раскололся суперконтинент Гондвана (рисунок 53, б). К концу эоцена движущаяся на север Индия натыкается на Азию, а кора в месте их столкновения сминается в Гималаи. Чуть позже в Евразию втыкается с юго-запада Африка (с еще не отделившейся от нее Аравией) – в результате поднимаются горные хребты южной Европы и Иранское нагорье. Океан Тетис, отделявший в мезозое северные (лавразийские) материки от южных (гондванских) – а именно по нему тогда шла экваториальная циркуляция – исчезает; ныне от Тетиса остались лишь изолированные морские бассейны (Средиземное, Черное и южная часть Каспийского моря), не имеющие постоянной связи с мировым океаном [70]. В южном полушарии Антарктида последовательно отрывается от Австралии (эоцен) и Южной Америки (олигоцен), «наползает» на Южный полюс, а вокруг нее складывается замкнутая антарктическая циркуляция; все это приводит к тому, что в олигоцене в Трансантарктических горах возникают первые ледники, которые в миоцене покрывают единым ледяным щитом весь материк. В плиоцене возникает Панамский перешеек – соединяющий Южную Америку с Северной, но разделяющий Атлантический и Тихий океаны, – и глобальная экваториальная циркуляция оказывается полностью разрушенной (ныне мы имеем лишь один ее сегмент – в центральной части Тихого океана). Разрушение экваториальной циркуляции, возникновение «антарктического холодильника» на Южном полюсе и замыкание северного полярного бассейна – все это приводит тому, что в четвертичном периоде (в плейстоцене) на материках Северного полушария развиваются грандиозные покровные оледенения (см. главу 14).
Надо заметить, что отсутствие в мезозое холодных климатических зон давно не вызывает сомнения у палеоклиматологов. Однако они зачастую не обращают внимания на то, что при более равномерном, чем ныне, распределении тепла по поверхности планеты не должно было существовать и климата типа современного тропического. Но тогда и сообщества, сложившиеся в условиях не существовавших ранее типов климата (бореального с одной стороны и тропического – с другой), должны быть эволюционно наиболее молоды... Если в некий момент на полюсах стало «слишком холодно», а на экваторе – «слишком жарко», то реликты предыдущей эпохи имеют наилучшие шансы выжить прежде всего в субтропиках, господствовавших ранее по всей Земле. Специальный анализ, проделанный В.В. Жерихиным (1978), показал, что так оно и есть. Судя по всему, некоторые субтропические сообщества (такие, как, например, леса из южного бука в Новой Зеландии, южной Австралии и андийской Южной Америке) сохранили не просто отдельные реликты раннего кайнофита, а самую структуру раннекайнофитных биоценозов.
Что же касается тропических лесов (мы будем называть их южноамериканским словом гилея), то укоренившиеся представления о глубокой древности их биоты являются очередным «научным предрассудком»; они связаны в основном с тем, что понятие «тропики» зачастую употребляют нечетко. В астрономических тропиках – т.е. между тропиками Рака и Козерога – действительно есть сообщества с весьма архаичными элементами (например, индонезийские «туманные леса» из древовидных папоротников), однако все они приурочены к горам, и существуют, на самом деле, в условиях субтропического или даже умеренного климата. Если же рассмотреть лишь климатические тропики, т.е. экосистемы, реально существующие в условиях тропического климата (с температурой самого холодного месяца не менее 18 С) – гилею и саванны, то окажется, что их биота практически лишена архаичных черт. Мы уже упоминали (глава 12) об отсутствии в составе тропических лесов голосеменных, столь характерных для мезозойских сообществ; ныне те приурочены либо к субтропикам (саговники, араукарии, таксодиум), либо к умеренной зоне (секвойя, гинкго).
Подсчеты Жерихина показали, что практически все по-настоящему древние группы насекомых (такие, как скорпионницы или сетчатокрылые) избегают тропиков. Общее число семейств, не встречающихся в истинных, климатических, тропиках очень велико, тогда как чисто тропических (в этом смысле) семейств среди насекомых – как это ни удивительно – ничтожно мало. Разумеется, тропическая энтомофауна чрезвычайно богата, и есть множество преимущественно тропических семейств – однако почти все они эволюционно молоды и не обнаружены в ископаемых донеогеновых фаунах, либо представлены в них очень скудно.
Еще более интересны в этом плане приводимые Жерихиным (1980) данные по экологической структуре тропических лесов:
"Сообщества типа современной гилеи с их сложной ярусной структурой и высокой степенью полидоминантности [71]вообще не могут существовать без участия филогенетически молодых групп. Полидоминантность в вечнозеленых лесах [в отличие от листопадных – К.Е.] поддерживается только наличием специализированных опылителей (пчелы, антофильные птицы и рукокрылые), поскольку анемофилия эффективна лишь для самых высокоствольных деревьев...
Деструкция органического вещества как в гилейных, так и в саванновых сообществах производится прежде всего термитами... Препятствуя образованию подстилки и выраженного гумусового горизонта (и тем самым – развитого травяного яруса) они, по-видимому, обусловили превращение многих травянистых растений в эпифиты и формирование эпифитных консорций, очень типичных для гилеи. Распределяя органическое вещество в мощном слое почвенного профиля более или менее равномерно, они обеспечивают возможность развития корневых систем на самых различных уровнях, в том числе и глубоких, что позволяет существовать гигантским деревьям с глубоко погруженными корнями...
В вечнозеленых лесах, где листья, не сменяясь, существуют на дереве по нескольку лет, требуется эффективное предотвращение сильного повреждения листвы фитофагами; в противном случае деятельность листового насоса не сможет обеспечить существование дерева. Функцию подавления численности открытоживущих фитофагов выполняют чрезвычайно обильные в гилее муравьи..." Все перечисленные выше группы, «определяющие лицо» тропических экоситем – и опылители, и термиты, и муравьи – эволюционно очень молоды и не характерны для раннекайнофитных сообществ. Все это и привело Жерихина к заключению, что тропические сообщества – наравне с бореальными – принадлежат к числу самых молодых на Земле; судя по всему, они возникли не раньше эоцена, а окончательное их формирование произошло уже в неогене.
С уверенностью говорить о времени формирования гилейных экосистем трудно из-за очень слабой палеонтологической изученности современной тропческой зоны. Что же касается бореальных сообществ – например, криофильных (холодолюбивых) лесов с доминированием хвойных, типа тайги, – то они, по всей видимости, сложились в палеогене как высотный пояс в горах, а при неогеновом похолодании широко распространились на равнинах Северного полушария. В палеонтологической летописи они впервые достоверно появляются в верхнем миоцене Канадского архипелага (остров Миен); их состав напоминал современные лесотундровые редколесья.
Еще одна характерная для кайнозоя черта – травяные биомы (типа степей и саванн), существование которых основано на коэволюции злаков и крупных травоядных млекопитающих: при ряде условий (в засушливых и малоплодородных районах) животные способны воспрепятствовать восстановлению лесной растительности. В доэоценовые времена, судя по пыльцевым спектрам, сообществ такого типа на Земле не было. По мнению Жерихина (1993), эти биомы первоначально представляли собою сериальные сообщества, ведущие к соответствующим древесным климаксам (стадии с господством трав – например, луга – имеются во всех современных сукцессионных системах), однако затем возникла уникальная ситуация: фитофаги, изымая прирост биомассы (в современных степях это изъятие доходит до 60% растительной продукции – абсолютный рекорд), оказались способны остановить экогенетическую сукцессию на безлесой стадии. Дальнейшая коэволюция млекопитающих и трав привела к стабилизации этих своеобразных «зоогенных климаксов» : необходимый для климаксного сообщества нулевой баланс по органике обеспечивается в них именно деятельностью животных-фитофагов. Основные черты, отличающие эти травяные климаксы и от сериальных травяных сообществ, и от лесов – исключительно высокая продуктивность, очень быстрый оборот органического вещества и наличие легко мобилизуемого запаса этой органики в необычайно плодородных почвах (типа черноземов).
Известно, что основой существования этих сообществ является замечательная коадаптация доминирующих в растительном покрове злаков и фитофагов: злаки не имеют эффективной защиты от поедания (вроде колючек, ядовитости и т.п.), зато способны компенсировать обгрызание надземных частей резким ускорением их регенерации. Поэтому регуляция в таком сообществе достигается предельно просто: при снижении пресса фитофагов первичная продукция тоже автоматически снижается – и наоборот. Основную роль в пастбищных цепях этих экосистем играют млекопитающие, прежде всего – различные копытные (потомки кондиляртр – рисунок 55, а) и хищные (потомки креодонтов – рисунок 56, а); об эволюции сообщества позвоночных – чуть далее. Иное дело – детритные цепи. Возникновение стабильных травяных сообществ было бы просто невозможно без появления целого комплекса насекомых-копрофагов, перерабатывающих огромную массу экскрементов травоядных [72]. В более древних (палеогеновых) лесных сообществах, где млекопитающие не достигали такой биомассы, как пастбищные копытные, с этой работой справлялись низшие навозники. Важнейшие же современные группы копрофагов (высшие навозники и навозные мухи) в заметных количествах появляются в захоронениях с конца олигоцена. В это же время складывается и не менее важный комплекс насекомых-некрофагов (мясные мухи и жуки-мертвоеды), ответственный за эффективную переработку трупов травоядных млекопитающих.
Сообщество наземных позвоночных в кайнозое развивалось независимо на трех разобщенных территориях, фаунистические контакты между которыми практически отсутствовали. Австралия (с ее сумчатыми и однопроходными) изолирована и поныне, а Южная Америка сохраняла свою обособленность от остальной суши вплоть до плиоцена, когда возник Панамский перешеек; отсюда и проистекает современное разделение мира на три зоогеографические области: Нотогея (Австралия), Неогея (Южная Америка) и Арктогея (Евразия, Африка и Северная Америка). Так вот, по мнению Жерихина (1993), во всех этих трех областях травяные биомы возникали независимо, на базе совершенно различных комплексов крупных млекопитающих; собственно говоря, есть серьезные основания полагать, что млекопитающие по-настоящему вышли в крупный размерный класс лишь в травяных биомах.
Раньше всего (в среднем эоцене) этот процесс начался в Южной Америке. Там среди исходно листоядных «южноамериканских копытных» [73]возникают первые травоядные формы, а также появляются гигантские травоядные броненосцы-глиптодонты, смахивающие на небольшой танк (рисунок 54, а). В среднем же эоцене в Южной Америке впервые обнаруживаются и пыльцевые спектры с высоким содержанием пыльцы злаков, палеопочвы степного типа, а также фоссилизированные навозные шары, принадлежащие жукам-навозникам. Позднее, в олигоцене и особенно в миоцене, здесь возникает в высшей степени своеобразный комплекс пастбищных травоядных. Он включал неполнозубых (глиптодонтов и наземных ленивцев), «южноамериканских копытных» (различные литоптерны демонстрирует сильное конвергентное сходство частью с лошадьми, частью с верблюдами, у пиротериев много общего со слонами, а среди нотоунгулят были формы, схожие и с носорогами, и с бегемотами, и с кроликами – риснок 54, б-г), а также гиганских кавиморфных грызунов (некоторые из этих родственников морской свинки достигали размеров носорога) и просуществовал вплоть до установления в плиоцене сухопутной связи с Северной Америкой.

РИСУНОК 54. Третичные млекопитающие Южной Америки: неполнозубые (а), «южноамериканские копытные» (б-г) и хищные сумчатые (д-е). (а) – глиптодонт, (б) – литоптерна Thoatherium, (в) – литоптерна Toxodon, (г) – нотоунгулят Macrauchenia, (д) – «сумчатый волк» Prothylacynus, (е) – «сумчатый саблезубый тигр» Thylacosmilus; черепа Thylacosmilus (ж) и «северной» саблезубой кошки Smilodon (з) (в одном масштабе).
Что же касается хищников, то они в древней южноамериканской фауне всегда были в дефиците. Ни один из здешних отрядов плацентарных по не вполне понятным причинам так и не дал плотоядных форм – эту роль исполняли исключительно сумчатые. Довольно разнообразные боргиениды несколько напоминали собак (а еще больше – тилацина, тасманийского «сумчатого волка»), а тилакосмилюс вполне заслуживает названия «сумчатый саблезубый тигр» и являет собою поразительный пример конвергенции с саблезубыми кошками Северного полушария (рисунок 54, д-е). Дефицит маммальных хищников (на «несбалансированность» здешних фаун обратили внимание А.С. Раутиан и Н.Н. Каландадзе, 1987) привел к тому, что эту нишу заполняли самые неожиданные персонажи. Так, с палеоцена по миоцен здесь существовали себекозухии – сухопутные крокодилы с высокой и узкой мордой (предполагают, что их образ жизни напоминал современных комодоских варанов), а в эоцене появились дожившие до плейстоцена фороракосы – гигантские (до 3 м ростом) нелетающие хищные птицы, принадлежащие к журавлеобразным.
В Австралии (Нотогее) формирование травяного биома началось много позже, в неогене; здесь явно сыграл роль дрейф этого континента по направлению от полюса к экватору – в результате значительная часть его территории попала в условия засушливого климата. Основу здешнего сообщества пастбищных млекопитающих составили крупные травоядные сумчатые – кенгуру и вымершие на памяти человека дипротодонты (их иногда, из-за двух крупных резцов, не слишком удачно называют «кроликами ростом с носорога»). Как и в древней южноамериканской фауне, здесь отчетливо прослеживается дефицит хищников: известны лишь два крупноразмерных хищных млекопитающих – тилацин (тасманийский сумчатый волк) и древесный тилаколео (которого по аналогии можно назвать «сумчатым леопардом»). Недостаток маммальных хищников возмещался (опять-таки как в Южной Америке) за счет рептилий: исполинских варанов-мегаланий длиною до 7 м и сухопутных крокодилов, сходных по образу жизни с себекозухиями; хищных нелетающих птиц здесь не возникло, однако некоторые из австралийских страусов, судя по всему, выполняли роль падальщиков.
Согласно моделям климатологов, наиболее интенсивный теплообмен будет достигнут при таком взаиморасположении материков и полюсов, когда планета имеет свободные от суши полюса и экватор (вдоль которого складывалась бы под действием кориолисовых сил единая экваториальная циркуляция). Меридионально развернутые материки, расположенные в средних широтах, отводят рукава экваториальной циркуляции в высокие широты, откуда вдоль противоположных их берегов возвращается в экваториальную зону холодная вода из полярных бассейнов – возвращается сразу, не застаиваясь в полярных циркуляциях. Как легко видеть (рисунок 53, а), мезозойская ситуация была в этом смысле близка к идеальной.
РИСУНОК 53. Материки и морские течения. Мезозойское (средний мел) (а) и современное (б) расположение материков.
В кайнозое картина становится близка к обратной, и причиною тому – перемещения материков, на которые раскололся суперконтинент Гондвана (рисунок 53, б). К концу эоцена движущаяся на север Индия натыкается на Азию, а кора в месте их столкновения сминается в Гималаи. Чуть позже в Евразию втыкается с юго-запада Африка (с еще не отделившейся от нее Аравией) – в результате поднимаются горные хребты южной Европы и Иранское нагорье. Океан Тетис, отделявший в мезозое северные (лавразийские) материки от южных (гондванских) – а именно по нему тогда шла экваториальная циркуляция – исчезает; ныне от Тетиса остались лишь изолированные морские бассейны (Средиземное, Черное и южная часть Каспийского моря), не имеющие постоянной связи с мировым океаном [70]. В южном полушарии Антарктида последовательно отрывается от Австралии (эоцен) и Южной Америки (олигоцен), «наползает» на Южный полюс, а вокруг нее складывается замкнутая антарктическая циркуляция; все это приводит к тому, что в олигоцене в Трансантарктических горах возникают первые ледники, которые в миоцене покрывают единым ледяным щитом весь материк. В плиоцене возникает Панамский перешеек – соединяющий Южную Америку с Северной, но разделяющий Атлантический и Тихий океаны, – и глобальная экваториальная циркуляция оказывается полностью разрушенной (ныне мы имеем лишь один ее сегмент – в центральной части Тихого океана). Разрушение экваториальной циркуляции, возникновение «антарктического холодильника» на Южном полюсе и замыкание северного полярного бассейна – все это приводит тому, что в четвертичном периоде (в плейстоцене) на материках Северного полушария развиваются грандиозные покровные оледенения (см. главу 14).
Надо заметить, что отсутствие в мезозое холодных климатических зон давно не вызывает сомнения у палеоклиматологов. Однако они зачастую не обращают внимания на то, что при более равномерном, чем ныне, распределении тепла по поверхности планеты не должно было существовать и климата типа современного тропического. Но тогда и сообщества, сложившиеся в условиях не существовавших ранее типов климата (бореального с одной стороны и тропического – с другой), должны быть эволюционно наиболее молоды... Если в некий момент на полюсах стало «слишком холодно», а на экваторе – «слишком жарко», то реликты предыдущей эпохи имеют наилучшие шансы выжить прежде всего в субтропиках, господствовавших ранее по всей Земле. Специальный анализ, проделанный В.В. Жерихиным (1978), показал, что так оно и есть. Судя по всему, некоторые субтропические сообщества (такие, как, например, леса из южного бука в Новой Зеландии, южной Австралии и андийской Южной Америке) сохранили не просто отдельные реликты раннего кайнофита, а самую структуру раннекайнофитных биоценозов.
Что же касается тропических лесов (мы будем называть их южноамериканским словом гилея), то укоренившиеся представления о глубокой древности их биоты являются очередным «научным предрассудком»; они связаны в основном с тем, что понятие «тропики» зачастую употребляют нечетко. В астрономических тропиках – т.е. между тропиками Рака и Козерога – действительно есть сообщества с весьма архаичными элементами (например, индонезийские «туманные леса» из древовидных папоротников), однако все они приурочены к горам, и существуют, на самом деле, в условиях субтропического или даже умеренного климата. Если же рассмотреть лишь климатические тропики, т.е. экосистемы, реально существующие в условиях тропического климата (с температурой самого холодного месяца не менее 18 С) – гилею и саванны, то окажется, что их биота практически лишена архаичных черт. Мы уже упоминали (глава 12) об отсутствии в составе тропических лесов голосеменных, столь характерных для мезозойских сообществ; ныне те приурочены либо к субтропикам (саговники, араукарии, таксодиум), либо к умеренной зоне (секвойя, гинкго).
Подсчеты Жерихина показали, что практически все по-настоящему древние группы насекомых (такие, как скорпионницы или сетчатокрылые) избегают тропиков. Общее число семейств, не встречающихся в истинных, климатических, тропиках очень велико, тогда как чисто тропических (в этом смысле) семейств среди насекомых – как это ни удивительно – ничтожно мало. Разумеется, тропическая энтомофауна чрезвычайно богата, и есть множество преимущественно тропических семейств – однако почти все они эволюционно молоды и не обнаружены в ископаемых донеогеновых фаунах, либо представлены в них очень скудно.
Еще более интересны в этом плане приводимые Жерихиным (1980) данные по экологической структуре тропических лесов:
"Сообщества типа современной гилеи с их сложной ярусной структурой и высокой степенью полидоминантности [71]вообще не могут существовать без участия филогенетически молодых групп. Полидоминантность в вечнозеленых лесах [в отличие от листопадных – К.Е.] поддерживается только наличием специализированных опылителей (пчелы, антофильные птицы и рукокрылые), поскольку анемофилия эффективна лишь для самых высокоствольных деревьев...
Деструкция органического вещества как в гилейных, так и в саванновых сообществах производится прежде всего термитами... Препятствуя образованию подстилки и выраженного гумусового горизонта (и тем самым – развитого травяного яруса) они, по-видимому, обусловили превращение многих травянистых растений в эпифиты и формирование эпифитных консорций, очень типичных для гилеи. Распределяя органическое вещество в мощном слое почвенного профиля более или менее равномерно, они обеспечивают возможность развития корневых систем на самых различных уровнях, в том числе и глубоких, что позволяет существовать гигантским деревьям с глубоко погруженными корнями...
В вечнозеленых лесах, где листья, не сменяясь, существуют на дереве по нескольку лет, требуется эффективное предотвращение сильного повреждения листвы фитофагами; в противном случае деятельность листового насоса не сможет обеспечить существование дерева. Функцию подавления численности открытоживущих фитофагов выполняют чрезвычайно обильные в гилее муравьи..." Все перечисленные выше группы, «определяющие лицо» тропических экоситем – и опылители, и термиты, и муравьи – эволюционно очень молоды и не характерны для раннекайнофитных сообществ. Все это и привело Жерихина к заключению, что тропические сообщества – наравне с бореальными – принадлежат к числу самых молодых на Земле; судя по всему, они возникли не раньше эоцена, а окончательное их формирование произошло уже в неогене.
С уверенностью говорить о времени формирования гилейных экосистем трудно из-за очень слабой палеонтологической изученности современной тропческой зоны. Что же касается бореальных сообществ – например, криофильных (холодолюбивых) лесов с доминированием хвойных, типа тайги, – то они, по всей видимости, сложились в палеогене как высотный пояс в горах, а при неогеновом похолодании широко распространились на равнинах Северного полушария. В палеонтологической летописи они впервые достоверно появляются в верхнем миоцене Канадского архипелага (остров Миен); их состав напоминал современные лесотундровые редколесья.
Еще одна характерная для кайнозоя черта – травяные биомы (типа степей и саванн), существование которых основано на коэволюции злаков и крупных травоядных млекопитающих: при ряде условий (в засушливых и малоплодородных районах) животные способны воспрепятствовать восстановлению лесной растительности. В доэоценовые времена, судя по пыльцевым спектрам, сообществ такого типа на Земле не было. По мнению Жерихина (1993), эти биомы первоначально представляли собою сериальные сообщества, ведущие к соответствующим древесным климаксам (стадии с господством трав – например, луга – имеются во всех современных сукцессионных системах), однако затем возникла уникальная ситуация: фитофаги, изымая прирост биомассы (в современных степях это изъятие доходит до 60% растительной продукции – абсолютный рекорд), оказались способны остановить экогенетическую сукцессию на безлесой стадии. Дальнейшая коэволюция млекопитающих и трав привела к стабилизации этих своеобразных «зоогенных климаксов» : необходимый для климаксного сообщества нулевой баланс по органике обеспечивается в них именно деятельностью животных-фитофагов. Основные черты, отличающие эти травяные климаксы и от сериальных травяных сообществ, и от лесов – исключительно высокая продуктивность, очень быстрый оборот органического вещества и наличие легко мобилизуемого запаса этой органики в необычайно плодородных почвах (типа черноземов).
Известно, что основой существования этих сообществ является замечательная коадаптация доминирующих в растительном покрове злаков и фитофагов: злаки не имеют эффективной защиты от поедания (вроде колючек, ядовитости и т.п.), зато способны компенсировать обгрызание надземных частей резким ускорением их регенерации. Поэтому регуляция в таком сообществе достигается предельно просто: при снижении пресса фитофагов первичная продукция тоже автоматически снижается – и наоборот. Основную роль в пастбищных цепях этих экосистем играют млекопитающие, прежде всего – различные копытные (потомки кондиляртр – рисунок 55, а) и хищные (потомки креодонтов – рисунок 56, а); об эволюции сообщества позвоночных – чуть далее. Иное дело – детритные цепи. Возникновение стабильных травяных сообществ было бы просто невозможно без появления целого комплекса насекомых-копрофагов, перерабатывающих огромную массу экскрементов травоядных [72]. В более древних (палеогеновых) лесных сообществах, где млекопитающие не достигали такой биомассы, как пастбищные копытные, с этой работой справлялись низшие навозники. Важнейшие же современные группы копрофагов (высшие навозники и навозные мухи) в заметных количествах появляются в захоронениях с конца олигоцена. В это же время складывается и не менее важный комплекс насекомых-некрофагов (мясные мухи и жуки-мертвоеды), ответственный за эффективную переработку трупов травоядных млекопитающих.
Сообщество наземных позвоночных в кайнозое развивалось независимо на трех разобщенных территориях, фаунистические контакты между которыми практически отсутствовали. Австралия (с ее сумчатыми и однопроходными) изолирована и поныне, а Южная Америка сохраняла свою обособленность от остальной суши вплоть до плиоцена, когда возник Панамский перешеек; отсюда и проистекает современное разделение мира на три зоогеографические области: Нотогея (Австралия), Неогея (Южная Америка) и Арктогея (Евразия, Африка и Северная Америка). Так вот, по мнению Жерихина (1993), во всех этих трех областях травяные биомы возникали независимо, на базе совершенно различных комплексов крупных млекопитающих; собственно говоря, есть серьезные основания полагать, что млекопитающие по-настоящему вышли в крупный размерный класс лишь в травяных биомах.
Раньше всего (в среднем эоцене) этот процесс начался в Южной Америке. Там среди исходно листоядных «южноамериканских копытных» [73]возникают первые травоядные формы, а также появляются гигантские травоядные броненосцы-глиптодонты, смахивающие на небольшой танк (рисунок 54, а). В среднем же эоцене в Южной Америке впервые обнаруживаются и пыльцевые спектры с высоким содержанием пыльцы злаков, палеопочвы степного типа, а также фоссилизированные навозные шары, принадлежащие жукам-навозникам. Позднее, в олигоцене и особенно в миоцене, здесь возникает в высшей степени своеобразный комплекс пастбищных травоядных. Он включал неполнозубых (глиптодонтов и наземных ленивцев), «южноамериканских копытных» (различные литоптерны демонстрирует сильное конвергентное сходство частью с лошадьми, частью с верблюдами, у пиротериев много общего со слонами, а среди нотоунгулят были формы, схожие и с носорогами, и с бегемотами, и с кроликами – риснок 54, б-г), а также гиганских кавиморфных грызунов (некоторые из этих родственников морской свинки достигали размеров носорога) и просуществовал вплоть до установления в плиоцене сухопутной связи с Северной Америкой.
РИСУНОК 54. Третичные млекопитающие Южной Америки: неполнозубые (а), «южноамериканские копытные» (б-г) и хищные сумчатые (д-е). (а) – глиптодонт, (б) – литоптерна Thoatherium, (в) – литоптерна Toxodon, (г) – нотоунгулят Macrauchenia, (д) – «сумчатый волк» Prothylacynus, (е) – «сумчатый саблезубый тигр» Thylacosmilus; черепа Thylacosmilus (ж) и «северной» саблезубой кошки Smilodon (з) (в одном масштабе).
Что же касается хищников, то они в древней южноамериканской фауне всегда были в дефиците. Ни один из здешних отрядов плацентарных по не вполне понятным причинам так и не дал плотоядных форм – эту роль исполняли исключительно сумчатые. Довольно разнообразные боргиениды несколько напоминали собак (а еще больше – тилацина, тасманийского «сумчатого волка»), а тилакосмилюс вполне заслуживает названия «сумчатый саблезубый тигр» и являет собою поразительный пример конвергенции с саблезубыми кошками Северного полушария (рисунок 54, д-е). Дефицит маммальных хищников (на «несбалансированность» здешних фаун обратили внимание А.С. Раутиан и Н.Н. Каландадзе, 1987) привел к тому, что эту нишу заполняли самые неожиданные персонажи. Так, с палеоцена по миоцен здесь существовали себекозухии – сухопутные крокодилы с высокой и узкой мордой (предполагают, что их образ жизни напоминал современных комодоских варанов), а в эоцене появились дожившие до плейстоцена фороракосы – гигантские (до 3 м ростом) нелетающие хищные птицы, принадлежащие к журавлеобразным.
В Австралии (Нотогее) формирование травяного биома началось много позже, в неогене; здесь явно сыграл роль дрейф этого континента по направлению от полюса к экватору – в результате значительная часть его территории попала в условия засушливого климата. Основу здешнего сообщества пастбищных млекопитающих составили крупные травоядные сумчатые – кенгуру и вымершие на памяти человека дипротодонты (их иногда, из-за двух крупных резцов, не слишком удачно называют «кроликами ростом с носорога»). Как и в древней южноамериканской фауне, здесь отчетливо прослеживается дефицит хищников: известны лишь два крупноразмерных хищных млекопитающих – тилацин (тасманийский сумчатый волк) и древесный тилаколео (которого по аналогии можно назвать «сумчатым леопардом»). Недостаток маммальных хищников возмещался (опять-таки как в Южной Америке) за счет рептилий: исполинских варанов-мегаланий длиною до 7 м и сухопутных крокодилов, сходных по образу жизни с себекозухиями; хищных нелетающих птиц здесь не возникло, однако некоторые из австралийских страусов, судя по всему, выполняли роль падальщиков.