Страница:
Многие авторы полагают, что речь идет о климатических изменениях. Действительно, в конце мела (маастрихтский век) началось заметное похолодание, а осушение морских шельфов вело к установлению более континентального климата с резкими температурными перепадами – инерциально-гомойотермным динозаврам все это и вправду было совершенно «не в масть». Однако больший интерес, на мой взгляд, представляют «биотические» гипотезы, касающиеся экосистемных перестроек. Среди неспециалистов достаточно популярные гипотезы о том, что растительноядные динозавры якобы не сумели приспособиться к новой диете – распространившейся в середине мела цветковой растительности: отравились вырабатываемыми цветковыми алкалоидами или активно концентрируемыми их тканями тяжелыми металлами
[64], сточили зубы о содержащиеся в их клетках кристаллы кремнезема, и т.п. Все это представляется абсолютно несерьезным – хотя бы потому, что на поздний мел просто приходится максимальный расцвет группы: из этого времени известно столько же видов динозавров, сколько за всю предшествующую историю – с триаса по конец раннего мела. Именно тогда, кстати, появляются специализированные потребители высокоабразивных кормов (типа злаков) с зубными батареями. Иное дело – события, происходившие в мелу внутри сообщества позвоночных.
Мы с вами помним, что основой 120-миллионолетнего «мирного сосуществования» динозавров и млекопитающих было полное разделение их экологических ниш в соответствии с различиями в размерных классах. В мелком размерном классе, формируемом млекопитающими, в те времена не существовало настоящих плотоядных форм – только насекомоядные и неспециализированные хищники, типа ежей или опоссумов. Ситуация поменялась в мелу, когда на эволюционную арену вышли териевые млекопитающие, имевшие заметно более высокий уровень метаболизма, нежели триасовые и юрские прототерии. На этой метаболической основе «истинно-гомойотермным» териям впервые удается создать фитофага в малом размерном классе – условно говоря, «крысу». Событие поистине революционное – легко понять, насколько теперь расширяется пищевая база субдоминантного сообщества [65]. Теперь в нем с неуклонностью должен появиться и управляющий блок из специализированных хищников – столь же условно говоря, «фокстерьер». И вот тут-то – совершенно неожиданно! – начинаются крупные неприятности у динозавров – ибо детеныши-то их являются членами не доминантного, а субдоминантного сообщества. Детеныш динозавра, не обладающий еще (в силу своих размеров) инерциальной гомойотермией – это просто большая ящерица, лакомая добыча для такого вот, круглосуточно активного, «фокстерьера».
Защитить детеныша – задача чепуховая... но только на первый взгляд. Это (как нам известно) хорошо удается слонам и крупным копытным, ведущим стадный образ жизни, но с динозаврами ситуация иная. Легко наладить охрану кладки (некоторые динозавры в позднем мелу действительно отрабатывают такие типы поведения), однако когда детеныш имеет размер кролика, а родители – ростом со слона, то его быстрее раздавишь, чем защитишь от нападения [66]. Сократить этот разрыв в размерах невозможно (размер детеныша ограничен максимальными размерами яйца: если сделать его слишком большим, скорлупа не выдержит давления жидкости, а если увеличить толщину скорлупы, зародыш задохнется), и молодь динозавров обречена оставаться членом субдоминантного сообщества. Интересно, кстати, что именно в это время хищные динозавры-тероподы совершают ряд попыток войти в малый размерный класс, чтобы тоже использовать такой новый пищевой ресурс, как «крысы», однако все они оканчиваются неудачей: сообщество уже сформировано, ниши поделены, а тероподы-крошки поспели к шапочному разбору – в точности как тероморфы в триасе.
Этот сценарий событий, предполагающий постепенное угасание динозавров за счет полной утери ими малого размерного класса (А.С. Раутиан образно назвал малый размерный класс «меристемой группы» – имея в виду, что именно из него исходят все эволюционные новации), представляется наиболее убедительным. Последнюю точку в истории «драконов мезозоя» действительно могло поставить маастрихтское похолодание и континентализация климата, однако это была именно «последняя соломинка, сломавшая спину верблюда». В любом случае, большинство палеозоологов не видит никаких оснований к тому, чтобы объяснять эти события внеземными (чуть было не сказал – «потусторонними») причинами.
12. Мезозойские биоценотические кризисы. «Ангиоспермизация мира» (средний мел) и «Великое вымирание» (конец мела). Импактные и биотические гипотезы.
Мы с вами помним, что основой 120-миллионолетнего «мирного сосуществования» динозавров и млекопитающих было полное разделение их экологических ниш в соответствии с различиями в размерных классах. В мелком размерном классе, формируемом млекопитающими, в те времена не существовало настоящих плотоядных форм – только насекомоядные и неспециализированные хищники, типа ежей или опоссумов. Ситуация поменялась в мелу, когда на эволюционную арену вышли териевые млекопитающие, имевшие заметно более высокий уровень метаболизма, нежели триасовые и юрские прототерии. На этой метаболической основе «истинно-гомойотермным» териям впервые удается создать фитофага в малом размерном классе – условно говоря, «крысу». Событие поистине революционное – легко понять, насколько теперь расширяется пищевая база субдоминантного сообщества [65]. Теперь в нем с неуклонностью должен появиться и управляющий блок из специализированных хищников – столь же условно говоря, «фокстерьер». И вот тут-то – совершенно неожиданно! – начинаются крупные неприятности у динозавров – ибо детеныши-то их являются членами не доминантного, а субдоминантного сообщества. Детеныш динозавра, не обладающий еще (в силу своих размеров) инерциальной гомойотермией – это просто большая ящерица, лакомая добыча для такого вот, круглосуточно активного, «фокстерьера».
Защитить детеныша – задача чепуховая... но только на первый взгляд. Это (как нам известно) хорошо удается слонам и крупным копытным, ведущим стадный образ жизни, но с динозаврами ситуация иная. Легко наладить охрану кладки (некоторые динозавры в позднем мелу действительно отрабатывают такие типы поведения), однако когда детеныш имеет размер кролика, а родители – ростом со слона, то его быстрее раздавишь, чем защитишь от нападения [66]. Сократить этот разрыв в размерах невозможно (размер детеныша ограничен максимальными размерами яйца: если сделать его слишком большим, скорлупа не выдержит давления жидкости, а если увеличить толщину скорлупы, зародыш задохнется), и молодь динозавров обречена оставаться членом субдоминантного сообщества. Интересно, кстати, что именно в это время хищные динозавры-тероподы совершают ряд попыток войти в малый размерный класс, чтобы тоже использовать такой новый пищевой ресурс, как «крысы», однако все они оканчиваются неудачей: сообщество уже сформировано, ниши поделены, а тероподы-крошки поспели к шапочному разбору – в точности как тероморфы в триасе.
Этот сценарий событий, предполагающий постепенное угасание динозавров за счет полной утери ими малого размерного класса (А.С. Раутиан образно назвал малый размерный класс «меристемой группы» – имея в виду, что именно из него исходят все эволюционные новации), представляется наиболее убедительным. Последнюю точку в истории «драконов мезозоя» действительно могло поставить маастрихтское похолодание и континентализация климата, однако это была именно «последняя соломинка, сломавшая спину верблюда». В любом случае, большинство палеозоологов не видит никаких оснований к тому, чтобы объяснять эти события внеземными (чуть было не сказал – «потусторонними») причинами.
12. Мезозойские биоценотические кризисы. «Ангиоспермизация мира» (средний мел) и «Великое вымирание» (конец мела). Импактные и биотические гипотезы.
Как уже было сказано, история наземных экосистем – это фактически история трех групп живых организмов: сосудистых растений, насекомых и позвоночных-тетрапод, а также их взаимоотношений. Основу окружающего нас кайнозойского мира составляют цветковые растения и экологически связанные с ними группы насекомых (таких, как, например, опылители – пчелы, мухи, бабочки), а также общественные насекомые (муравьи и термиты); наиболее заметными членами сообщества позвоночных являются плацентарные млекопитающие и птицы. Если говорить о водоемах, то тут доминируют настоящие костистые рыбы; верхние трофические этажи в морях формируют морские млекопитающие (китообразные и ластоногие) и акулы современного типа.
Мезозойский мир выглядел совершенно иначе. Растительную основу его составляли голосеменные (помимо хвойных – очень разнообразные гинкговые, саговники и родственые им беннетиты), в сообществе позвоночных доминировали архозавры – динозавры на земле, птерозавры в воздухе. Рыбы представлены в основном ганоидами [67], в морях главными хищниками являлись эвриапсидные рептилии (ихтиозавры и плезиозавры), а также головоногие (аммониты и белемниты). Различия среди насекомых не менее существенны: в те времена доминировали занимающие ныне подчиненное положение тараканы, цикады, сетчатокрылые и скорпионницы. Когда же именно возник этот наш кайнозойский мир? Произошла ли эта смена более или менее единовременно во всех группах, либо была растянутой во времени? Было ли это плавным замещением «ветеранов» «новобранцами», или, напротив, кризисом, сопряженным с массовыми вымираниями мезозойских групп?
О резких изменениях планетарной биоты в конце мелового периода ученые знали уже в XVIII веке; происходившие тогда события часто называют «Великим вымиранием». Наибольшее внимание обычно привлекает исчезновение гигантских рептилий (динозавров и птерозавров на суше, плезиозавров и мозозавров в море), однако помимо них в это время вымирают аммониты и белемниты, иноцерамы и рудисты (крупные – размером до метра – двустворки) и еще множество морских групп. Особено сильно пострадал планктон: раковинные простейшие – радиолярии и форамениферы, одноклеточные водоросли с известковым (кокколитофориды) и кремнеземовым (диатомеи) скелетом. При этом меловые виды обычны и разнообразны вплоть до последнего миллиметра отложений перед мезозойско-кайнозойской границей. Из данных магнитостратиграфии следует, что наибольшие изменения фауны соответствуют периоду обращенной полярности магнитного поля Земли (так называемый интервал 39), длившемуся не более 500 тыс лет. Иными словами, смена мезозойской биоты на кайнозойскую произошла внезапно и очень быстро, что наводит на мысль о какой-то глобальной катастрофе; а поскольку изменения эти охватили как морские, так и сухопутные группы, экологически не связанные между собою, логично связать ее с какой-то внешней по отношению к биосфере причиной – земной (катастрофический вулканизм и т.п.) или космической (вспышка сверхновой, многократное усиление излучения из космоса из-за переполюсовки магнитного поля планеты, падение гигантского астероида и т.п.).
Здесь следует сделать одно отступление, касающееся методологии. Мы с вами уже говорили о том, что основу европейского научного мышления составляет «Бритва Оккама» (см. главу 1-а) – из нескольких конкурирующих гипотез выбирается наиболее простая, не вводящая «избыточных сущностей». Всё так, но только вот представления о «простоте» и о том, какие из сущностей являются «избыточными» могут довольно сильно различаться у представителей разных наук.
Применительно к нашему конкретному случаю это означает следующее. Для физика более простыми, а потому более привлекательными, будут «однофакторные», импактные, гипотезы (извергнулись разом все вулканы – и всё живое отравилось серой и тяжелыми металлами; упал астероид – и поднятая им пыль настолько замутнила атмосферу, что растения перестали фотосинтезировать); представления же о системных свойствах биосферы являются «избыточными».
Иное дело – биолог; для него вполне очевидно, что экосистемы и биосфера являют собою не механические наборы живых организмов (вроде террариумов на Птичьем рынке), а высокоцелостные гомеостаты, обладающие своим собственным поведением, способные кондиционировать среду своего обитания и эффективно противостоять внешним возмущениям. Поэтому с точки зрения биолога более простыми будут гипотезы, предполагающие внутренние причины экосистемных смен – даже наиболее резких, кризисных (превращения червеобразной гусеницы в твердую неподвижную куколку, а куколки – в бабочку выглядят со стороны явными «кризисами» в процессе индивидуального развития насекомого), а внешние воздействия – «избыточной сущностью», которую можно привлекать лишь после того, как исчерпаны все иные, «внутрибиосферные» варианты объяснения. Это различие между «физиками» и «биологами» (названия эти, разумеется, условны) носит в некотором смысле мировоззренческий характер, и оттого лучше загодя обозначить свою позицию; я, как легко догадаться, занимаю «биологическую» позицию – положение обязывает...
Итак, возвращаемся к «Великому вымиранию». В середине 60-х годов при исследовании разреза пограничных мел-кайнозойских отложений в Губбио (Италия) в тонком слое глины, соответствующем фаунистической границе (его возраст – 67 млн лет), была обнаружена необычайно высокая концентрация редкого металла иридия (Ir), в 20 раз превышающая его среднее содержание в земной коре. Впоследствии такие иридиевые аномалии на мел-кайнозойской границе были найдены во множестве мест по всему миру, а содержание металла в некоторых из них превышает фоновое в 120 раз; протяженность периода, когда накапливались эти осадки, была (судя по толщине их слоя) очень невелика – не более 10 тысяч лет. В земной коре иридий редок потому, что он хорошо растворим в железе: почти весь земной иридий сконцентрирован сейчас в ядре планеты. Соответственно, его довольно много в веществе железных метеоритов, представляющих собой фрагменты планетных ядер.
На этом основании Л. Алварес (1980) предположил, что иридиевая аномалия – следствие удара о Землю крупного астероида, вещество которого рассеялось по всей ее поверхности (астероидная гипотеза). Чтобы дать наблюдаемое количество иридия, требуется тело внеземной природы диаметром приблизительно 10 км и массой 1010 т; при его падении на сушу возникла бы воронка диметром около 100 км (главным претендентом на эту роль считают кратер Чикксулуб на Юкатанском полуострове). Расчеты показывают, что при таком ударе в атмосферу было выброшено огромное количество пылевидного материала (в 60 раз больше массы самого астероида). Альварес и его сторонники полагают, что эта пыль обращалась вокруг Земли несколько лет, прежде чем выпала обратно на поверхность. Это плотное пылевое облако, сквозь которое слабо проходит солнечный свет, должно было сильно ослабить фотосинтез, что привело к гибели растений (прежде всего -планктонных водорослей, имеющих очень короткий жизненный цикл), а затем и питающихся ими животных. Кроме того, это должно было вызвать резкое охлаждение поверхности планеты («астероидная зима»). В воде океанов из-за ослабления фотосинтеза накапливается CO2, что ведет к повышению ее кислотности, а это, в свою очередь, – к растворению раковин карбонатсекретирующих организмов (от фораменифер до иноцерамов).
Р. Кэрролл (1993) в своем учебнике палеонтологии пишет: «Ученые-физики, признавая основные выводы Альвареса, оспаривают некоторые детали. Специалисты по ископаемым остаткам обычно настроены очень критически»; А.Г. Пономаренко весьма точно отразил отношение палеозоологов к «импактным» гипотезам, завершив соответствующую лекцию словами: «К несчастью, воззрения эти в последние годы распространились, как чума». Палеонтологи, конечно, не могут профессионально оценить математические модели, из коих следует принципиальная, теоретическая возможность наступления «астероидной зимы» (хотя по некоторым расчетам пыль должна была осесть не через несколько лет, а уже через несколько недель, а гидрохимики напоминают о существовании в морской воде карбонатно-бикарбонатного буфера, изменить кислотность которого чрезвычайно трудно), но вот о конкретных событиях конца мела им очень даже есть чего сказать. Суть дела в том, что наличие причинной связи между иридиевой аномалией (которая факт) и позднемеловым вымиранием (которое тоже факт), вообще-то говоря, решительно ни из чего не следует.
Начнем с того, что иридиевых аномалий в отложениях самого различного возраста найдено (с той поры, как их стали целенаправленно искать) уже несколько десятков – но только они никак не связаны с крупными фаунистическими сменами. И наоборот – все попытки обнаружить следы астероидных импактов в горизонтах, соответствующих другим крупным вымираниям, таким, например, как пермо-триасовое (по справедливости «Великим» следовало бы назвать именно его – оно было куда более масштабным, чем мел-палеогеновое) ни к чему не привели. Существует целый ряд точно датированных метеоритных кратеров даже более крупного размера, чем предполагаемый Альваресом (до 300 км в диаметре), и при этом достоверно известно, что ничего серьезного с биотой Земли в те моменты не происходило. Скурпулезное, «по миллиметрам», изучение пограничных разрезов показало, что синхронность иридиевой аномалии и «Великого вымирания» сильно преувеличена. Массовое вымирание морских организмов было «мгновенным» лишь по геологическим меркам и продолжалось, по разным оценкам, от 10 до 100 тысячелетий (а вовсе не годы – как это должно было быть по импактным моделям). Последовательность исчезновения планктонных организмов в разных местах неодинакова, а пики вымирания могут расходиться со временем иридиевой аномалии на десятки тысяч лет, причем многие группы (например, белемниты) вымирают до аномалии, а не после нее.
В этом смысле весьма показательна ситуация с Эльтанинским астероидом (около 4 км в поперечнике), упавшим в позднем плиоцене, около 2.5 млн лет назад, на шельф между Южной Америкой и Антарктидой; остатки астероида были недавно подняты из образовавшегося в морском дне кратера. Последствия этого падения выглядят вполне катастрофическими – километровые цунами забрасывали морскую фауну вглубь суши; именно тогда на андийском побережье возникли очень странные захоронения фауны со смесью морских и сухопутных форм, а в антарктических озерах вдруг появляются чисто морские диатомовые водоросли. Что же касается отдаленных, эволюционно значимых последствий, то их просто не было: следы этого импакта заключены внутри одной стратиграфической зоны. То есть – абсолютно никаких вымираний за всеми этими ужасными пертурбациями не последовало.
Обращает на себя внимание тот факт, что из всех групп, ставших жертвами «Великого вымирания», наземными были лишь динозавры (которые к тому же, как мы помним, вымерли несколько позже иридиевой аномалии); не морской группой были и вымершие тогда же птерозавры, но те, судя по всему, были трофически связаны с морем. (Кстати, упоминавшееся выше грандиозное пермо-триасовое вымирание было чисто морским – на суше тогда тоже не произошло ничего примечательного.) Если же обратиться к меловой континентальной биоте, то окажется, что радикальные изменения в составе ее ключевых групп (растений и насекомых) произошли примерно на 30 млн лет раньше – в конце раннего мела; А.Г. Пономаренко удачно назвал эти события «ангиоспермизацией мира», выделив важнейшую их черту – массовое распространение ангиоспермов (цветковых растений). Здесь можно вспомнить, что ботаники иногда стремятся использовать свою собственную временную шкалу и делить историю растительности не на «палеозой, мезозой и кайнозой», а на «палеофит, мезофит и кайнофит». Так вот, граница между «мезофитом» и «кайнофитом» проходит внутри мелового периода: с точки зрения ботаника (и – как мы увидим дальше – энтомолога) поздний мел – это фактически уже кайнозой.
Появление и распространение покрытосеменных – одно из ключевых событий в истории биосферы, а вызванные им изменения сопоставимы по своим масштабам с появлением наземных (сосудистых) растений или животных с минеральным скелетом. Нынешний мир – ангиоспермный; голосеменным и споровым пришел в мелу такой же конец, как в позднем палеозое – водорослевым матам (при появлении сосудистой растительности). Ныне голосеменные растут лишь там, где им «позволяют»: сосна растет на песках и болоте, а лиственница – на вечной мерзлоте совсем не оттого, что им там хорошо, а оттого, что на эти неудобья «не позарился» никто из цветковых. В некоторых ландшафтах (в тропических лесах и в пустынях) голосеменных нет вовсе [68].
Если задать ботанику вопрос – какие черты покрытосеменных привели их к эволюционному успеху, он наверняка начнет говорить о двойном оплодотворении, о триплоидном эндосперме и т.п.; это всё, конечно, верно – но немножко не о том. «С точки зрения биосферы» наиболее существенны, пожалуй, три свойства цветковых, позволившие им радикально изменить самую структуру мезозойских экосистем.
1/ Склонность к неотении (неотения – обретение организмом половозрелости на ранних (личиночных) стадиях своего индивидуального развития), то есть – в данном конкретном случае – к образованию травянистых форм, которые по сути дела есть древесные проростки, ставшие способными к размножению; напомним, что голосеменные «траву» не образуют вовсе – ни сейчас, ни в прошлом. Именно травянистые двудольные являются самыми агрессивными эксплерентами («сорными растениями»), мгновенно захватывающими места нарушений растительного покрова, а травянистые однодольные, склонные к образованию многолетней дернины, наиболее эффективно препятствуют эрозии.
2. Появление «мясистых» листьев с большим объемом фотосинтезирующей паренхимы, вызвавшее появление листогрызущих насекомых. Дело в том, что до того насекомые потребляли лишь генеративные органы, содержимое сосудов, даже древесину – но только не листья: в листе голосеменного (даже такого, как гинкго), попросту говоря, нечего есть. Ситуация менятся лишь с появлением цветковых – объеденные листья и мины (ходы, проедаемые личинками насекомых в толще листа) известны с самого начала позднего мела (сеноманский век). На покрытосеменных сформировалась общирная фауна листогрызущих насекомых, и уже после этого они перешли на голосеменные, папоротники и хвощи.
3. Энтомофилия цветков, открывающая покрытосеменным путь к сложной коэволюции с насекомыми. Собственно говоря, сам цветок покрытосеменных, судя по всему, сформировался в процессе адаптаций к насекомоопылению для защиты завязи от повреждений ее опылителями.
Ангиоспермы впервые появляются в палеонтологической летописи в начале раннего мела, в неокоме; на протяжении барремского и аптского веков они представлены единичными находками, в альбе идет стремительное нарастание их числа, а уже в самом начале позднего мела, в сеномане, цветковые составляют большую часть ископаемых флор – то есть наступает «кайнофит» (рисунок 50). Среднемеловая экспансия цветковых была столь стремительной (она фактически уложилась в ничтожный по геологический меркам отрезок от 115 до 108 млн лет), что Дарвин в свое время в сердцах назвал этот эпизод палеонтологической истории «отвратительной тайной – abominable mistery». Делались попытки объяснить это тем, что цветковые на самом-то деле появились на Земле давным-давно, минимум в юре – просто они обитали лишь в горах (не попадая по этой причине в палеонтологическую летопись), а в середине мела заселили, наконец, приводные местообитания, благоприятствующие попаданию в захоронения. Вообще такой «период скрытого существования» сбрасывать со счетов никак нельзя – но в данном случае эта гипотеза не проходит: дело в том, что пыльца покрытосеменных (которая, ясное дело, летает везде, и ее не «упрячешь» в горы) появляется в летописи одновременно с их макроостатками. По всем признакам, цветковые действительно возникли в начале мела, причем именно в приводных местообитаниях.
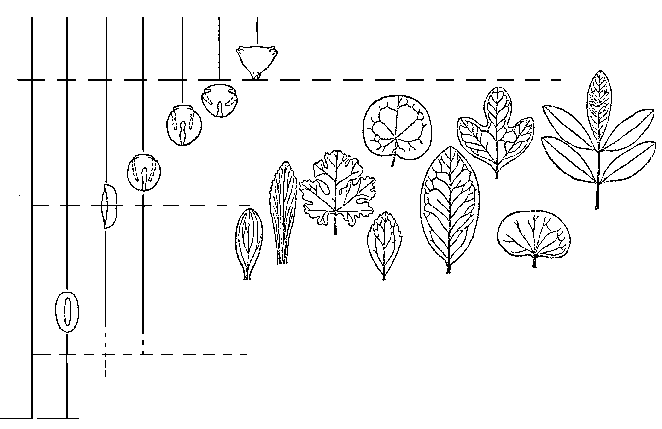
РИСУНОК 50. Распределение в меловых отложениях основных типов пыльцы и листьев покрытосеменных.
В 70-х годах советские палеоэнтомологи исследовали множество местонахождений меловых насекомых в Сибири и Монголии, «закрасив» тем самым самое крупное «белое пятно» в геологической истории класса. Оказалось, что у насекомых график, который отражает долю вымерших таксонов в фаунах разных времен, ведет себя довольно неожиданным образом (рисунок 51). Доля вымерших семейств мало меняется на протяжении большей части мезозоя, и на границе юры и мела составляет около 60% ; затем, в течении раннего мела, доля эта стремительно падает от 60 до 20% , в позднем мелу это падение резко замедляется (от 20 до 5%), а к началу кайнозоя кривая вновь «выходит на плато», только уже в районе 5%. Иными словами, у насекомых самая резкая фаунистическая смена, связанная с массовым вымиранием мезозойских групп, приходится не на конец мела, а на его середину – в точности как у растений. На этом основании В.В. Жерихин (1978) предложил следующую модель среднемелового биоценотического кризиса.
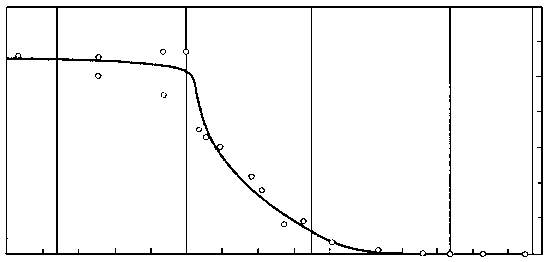
РИСУНОК 51. Эволюция насекомых в мезозое: процент вымерших семейств от общего числа семейств насекомых в позднемезозойских и кайнозойских фаунах.
Покрытосеменные возникли в начале мелового периода, первоначально как ценофобы – растения, не входящие в закономерные, сложившиеся в длительной коэволюции сукцессионные ряды [69]. Они росли в качестве «сорной» растительности по свободным от других растений участкам (на речных отмелях, береговых оползнях, гарях), которые соседствовали с гораздо более обширными участками, занятыми зрелыми сообществами мезофитной растительности. Покрытосеменные были исходно энтомофильными и смогли сформироваться лишь в окружении энтомофильных же голосеменных (таких, как беннетиты, саговники, кейтониевые), используя уже сложившийся к тому времени комплекс их опылителей. Но если покрытосеменные оказались среди пионерных растений мезофита единственными энтомофилами (а, судя по всему, так оно и было), то это должно было дать им гиганские преимущества: именно для локально распределенных, не образующих сплошных зарослей пионеров насекомоопыление особенно выгодно; впрочем, возможно, имело место и прямое конкурентное вытеснение покрытосеменными энтомофильных голосеменных из пионерных сообществ – за счет более эффективного привлечения насекомых, лучшей защиты от повреждения завязи опылителями и т.д.
Основой стремительной среднемеловой экспансии цветковых стало то, что им удалось закрепиться в качестве нормальной пионерной растительности – для чего решающими факторами стали их исходная энтомофильность и наличие среди них травянистых форм (что во много раз убыстряет зарастание поврежденных участков). Покрытосеменные не пытались потеснить всю мезофитную растительность, что было абсолютно нереально; они «всего-навсего» конкурентно вытеснили прежних пионеров – и тем самым блокировали все последующие стадии мезофитной экогенетической сукцессии. Разрушение существовавших в то время закономерных сукцессионных рядов (попросту говоря – мезофитная растительность продолжала существовать там, где она существует, но потеряла способность восстанавливаться после экзогенных нарушений), вызвало полный развал мезозойских наземных экосистем и массовое вымирание входивших в них животных; наиболее интенсивно эта деструкция шла в альбе (захватывая конец апта и начало сеномана).
В.В. Жерихин (1978) обратил внимание на любопытнейшую черту энтомофаун этого времени: многие семейства, характерные для мезозоя, обнаруживаются в них в последний раз, тогда как сколько-нибудь заметного появления новых групп не отмечается, и при этом появляется целый ряд представителей реликтовых для того времени (раннемезозойских и даже позднепалеозойских) групп. Это «всплывание реликтов» считают одним из наиболее ярких симптомов кризиса, охватившего среднемеловые сообщества: вымирание доминирующих раннемеловых групп (наиболее тесно связанных с существующей сукцессионной системой) создает экологический вакуум, который ненадолго заполняют древние формы, оттесненные к тому времени «на обочину жизни».
Мезозойский мир выглядел совершенно иначе. Растительную основу его составляли голосеменные (помимо хвойных – очень разнообразные гинкговые, саговники и родственые им беннетиты), в сообществе позвоночных доминировали архозавры – динозавры на земле, птерозавры в воздухе. Рыбы представлены в основном ганоидами [67], в морях главными хищниками являлись эвриапсидные рептилии (ихтиозавры и плезиозавры), а также головоногие (аммониты и белемниты). Различия среди насекомых не менее существенны: в те времена доминировали занимающие ныне подчиненное положение тараканы, цикады, сетчатокрылые и скорпионницы. Когда же именно возник этот наш кайнозойский мир? Произошла ли эта смена более или менее единовременно во всех группах, либо была растянутой во времени? Было ли это плавным замещением «ветеранов» «новобранцами», или, напротив, кризисом, сопряженным с массовыми вымираниями мезозойских групп?
О резких изменениях планетарной биоты в конце мелового периода ученые знали уже в XVIII веке; происходившие тогда события часто называют «Великим вымиранием». Наибольшее внимание обычно привлекает исчезновение гигантских рептилий (динозавров и птерозавров на суше, плезиозавров и мозозавров в море), однако помимо них в это время вымирают аммониты и белемниты, иноцерамы и рудисты (крупные – размером до метра – двустворки) и еще множество морских групп. Особено сильно пострадал планктон: раковинные простейшие – радиолярии и форамениферы, одноклеточные водоросли с известковым (кокколитофориды) и кремнеземовым (диатомеи) скелетом. При этом меловые виды обычны и разнообразны вплоть до последнего миллиметра отложений перед мезозойско-кайнозойской границей. Из данных магнитостратиграфии следует, что наибольшие изменения фауны соответствуют периоду обращенной полярности магнитного поля Земли (так называемый интервал 39), длившемуся не более 500 тыс лет. Иными словами, смена мезозойской биоты на кайнозойскую произошла внезапно и очень быстро, что наводит на мысль о какой-то глобальной катастрофе; а поскольку изменения эти охватили как морские, так и сухопутные группы, экологически не связанные между собою, логично связать ее с какой-то внешней по отношению к биосфере причиной – земной (катастрофический вулканизм и т.п.) или космической (вспышка сверхновой, многократное усиление излучения из космоса из-за переполюсовки магнитного поля планеты, падение гигантского астероида и т.п.).
Здесь следует сделать одно отступление, касающееся методологии. Мы с вами уже говорили о том, что основу европейского научного мышления составляет «Бритва Оккама» (см. главу 1-а) – из нескольких конкурирующих гипотез выбирается наиболее простая, не вводящая «избыточных сущностей». Всё так, но только вот представления о «простоте» и о том, какие из сущностей являются «избыточными» могут довольно сильно различаться у представителей разных наук.
Применительно к нашему конкретному случаю это означает следующее. Для физика более простыми, а потому более привлекательными, будут «однофакторные», импактные, гипотезы (извергнулись разом все вулканы – и всё живое отравилось серой и тяжелыми металлами; упал астероид – и поднятая им пыль настолько замутнила атмосферу, что растения перестали фотосинтезировать); представления же о системных свойствах биосферы являются «избыточными».
Иное дело – биолог; для него вполне очевидно, что экосистемы и биосфера являют собою не механические наборы живых организмов (вроде террариумов на Птичьем рынке), а высокоцелостные гомеостаты, обладающие своим собственным поведением, способные кондиционировать среду своего обитания и эффективно противостоять внешним возмущениям. Поэтому с точки зрения биолога более простыми будут гипотезы, предполагающие внутренние причины экосистемных смен – даже наиболее резких, кризисных (превращения червеобразной гусеницы в твердую неподвижную куколку, а куколки – в бабочку выглядят со стороны явными «кризисами» в процессе индивидуального развития насекомого), а внешние воздействия – «избыточной сущностью», которую можно привлекать лишь после того, как исчерпаны все иные, «внутрибиосферные» варианты объяснения. Это различие между «физиками» и «биологами» (названия эти, разумеется, условны) носит в некотором смысле мировоззренческий характер, и оттого лучше загодя обозначить свою позицию; я, как легко догадаться, занимаю «биологическую» позицию – положение обязывает...
Итак, возвращаемся к «Великому вымиранию». В середине 60-х годов при исследовании разреза пограничных мел-кайнозойских отложений в Губбио (Италия) в тонком слое глины, соответствующем фаунистической границе (его возраст – 67 млн лет), была обнаружена необычайно высокая концентрация редкого металла иридия (Ir), в 20 раз превышающая его среднее содержание в земной коре. Впоследствии такие иридиевые аномалии на мел-кайнозойской границе были найдены во множестве мест по всему миру, а содержание металла в некоторых из них превышает фоновое в 120 раз; протяженность периода, когда накапливались эти осадки, была (судя по толщине их слоя) очень невелика – не более 10 тысяч лет. В земной коре иридий редок потому, что он хорошо растворим в железе: почти весь земной иридий сконцентрирован сейчас в ядре планеты. Соответственно, его довольно много в веществе железных метеоритов, представляющих собой фрагменты планетных ядер.
На этом основании Л. Алварес (1980) предположил, что иридиевая аномалия – следствие удара о Землю крупного астероида, вещество которого рассеялось по всей ее поверхности (астероидная гипотеза). Чтобы дать наблюдаемое количество иридия, требуется тело внеземной природы диаметром приблизительно 10 км и массой 1010 т; при его падении на сушу возникла бы воронка диметром около 100 км (главным претендентом на эту роль считают кратер Чикксулуб на Юкатанском полуострове). Расчеты показывают, что при таком ударе в атмосферу было выброшено огромное количество пылевидного материала (в 60 раз больше массы самого астероида). Альварес и его сторонники полагают, что эта пыль обращалась вокруг Земли несколько лет, прежде чем выпала обратно на поверхность. Это плотное пылевое облако, сквозь которое слабо проходит солнечный свет, должно было сильно ослабить фотосинтез, что привело к гибели растений (прежде всего -планктонных водорослей, имеющих очень короткий жизненный цикл), а затем и питающихся ими животных. Кроме того, это должно было вызвать резкое охлаждение поверхности планеты («астероидная зима»). В воде океанов из-за ослабления фотосинтеза накапливается CO2, что ведет к повышению ее кислотности, а это, в свою очередь, – к растворению раковин карбонатсекретирующих организмов (от фораменифер до иноцерамов).
Р. Кэрролл (1993) в своем учебнике палеонтологии пишет: «Ученые-физики, признавая основные выводы Альвареса, оспаривают некоторые детали. Специалисты по ископаемым остаткам обычно настроены очень критически»; А.Г. Пономаренко весьма точно отразил отношение палеозоологов к «импактным» гипотезам, завершив соответствующую лекцию словами: «К несчастью, воззрения эти в последние годы распространились, как чума». Палеонтологи, конечно, не могут профессионально оценить математические модели, из коих следует принципиальная, теоретическая возможность наступления «астероидной зимы» (хотя по некоторым расчетам пыль должна была осесть не через несколько лет, а уже через несколько недель, а гидрохимики напоминают о существовании в морской воде карбонатно-бикарбонатного буфера, изменить кислотность которого чрезвычайно трудно), но вот о конкретных событиях конца мела им очень даже есть чего сказать. Суть дела в том, что наличие причинной связи между иридиевой аномалией (которая факт) и позднемеловым вымиранием (которое тоже факт), вообще-то говоря, решительно ни из чего не следует.
Начнем с того, что иридиевых аномалий в отложениях самого различного возраста найдено (с той поры, как их стали целенаправленно искать) уже несколько десятков – но только они никак не связаны с крупными фаунистическими сменами. И наоборот – все попытки обнаружить следы астероидных импактов в горизонтах, соответствующих другим крупным вымираниям, таким, например, как пермо-триасовое (по справедливости «Великим» следовало бы назвать именно его – оно было куда более масштабным, чем мел-палеогеновое) ни к чему не привели. Существует целый ряд точно датированных метеоритных кратеров даже более крупного размера, чем предполагаемый Альваресом (до 300 км в диаметре), и при этом достоверно известно, что ничего серьезного с биотой Земли в те моменты не происходило. Скурпулезное, «по миллиметрам», изучение пограничных разрезов показало, что синхронность иридиевой аномалии и «Великого вымирания» сильно преувеличена. Массовое вымирание морских организмов было «мгновенным» лишь по геологическим меркам и продолжалось, по разным оценкам, от 10 до 100 тысячелетий (а вовсе не годы – как это должно было быть по импактным моделям). Последовательность исчезновения планктонных организмов в разных местах неодинакова, а пики вымирания могут расходиться со временем иридиевой аномалии на десятки тысяч лет, причем многие группы (например, белемниты) вымирают до аномалии, а не после нее.
В этом смысле весьма показательна ситуация с Эльтанинским астероидом (около 4 км в поперечнике), упавшим в позднем плиоцене, около 2.5 млн лет назад, на шельф между Южной Америкой и Антарктидой; остатки астероида были недавно подняты из образовавшегося в морском дне кратера. Последствия этого падения выглядят вполне катастрофическими – километровые цунами забрасывали морскую фауну вглубь суши; именно тогда на андийском побережье возникли очень странные захоронения фауны со смесью морских и сухопутных форм, а в антарктических озерах вдруг появляются чисто морские диатомовые водоросли. Что же касается отдаленных, эволюционно значимых последствий, то их просто не было: следы этого импакта заключены внутри одной стратиграфической зоны. То есть – абсолютно никаких вымираний за всеми этими ужасными пертурбациями не последовало.
Обращает на себя внимание тот факт, что из всех групп, ставших жертвами «Великого вымирания», наземными были лишь динозавры (которые к тому же, как мы помним, вымерли несколько позже иридиевой аномалии); не морской группой были и вымершие тогда же птерозавры, но те, судя по всему, были трофически связаны с морем. (Кстати, упоминавшееся выше грандиозное пермо-триасовое вымирание было чисто морским – на суше тогда тоже не произошло ничего примечательного.) Если же обратиться к меловой континентальной биоте, то окажется, что радикальные изменения в составе ее ключевых групп (растений и насекомых) произошли примерно на 30 млн лет раньше – в конце раннего мела; А.Г. Пономаренко удачно назвал эти события «ангиоспермизацией мира», выделив важнейшую их черту – массовое распространение ангиоспермов (цветковых растений). Здесь можно вспомнить, что ботаники иногда стремятся использовать свою собственную временную шкалу и делить историю растительности не на «палеозой, мезозой и кайнозой», а на «палеофит, мезофит и кайнофит». Так вот, граница между «мезофитом» и «кайнофитом» проходит внутри мелового периода: с точки зрения ботаника (и – как мы увидим дальше – энтомолога) поздний мел – это фактически уже кайнозой.
Появление и распространение покрытосеменных – одно из ключевых событий в истории биосферы, а вызванные им изменения сопоставимы по своим масштабам с появлением наземных (сосудистых) растений или животных с минеральным скелетом. Нынешний мир – ангиоспермный; голосеменным и споровым пришел в мелу такой же конец, как в позднем палеозое – водорослевым матам (при появлении сосудистой растительности). Ныне голосеменные растут лишь там, где им «позволяют»: сосна растет на песках и болоте, а лиственница – на вечной мерзлоте совсем не оттого, что им там хорошо, а оттого, что на эти неудобья «не позарился» никто из цветковых. В некоторых ландшафтах (в тропических лесах и в пустынях) голосеменных нет вовсе [68].
Если задать ботанику вопрос – какие черты покрытосеменных привели их к эволюционному успеху, он наверняка начнет говорить о двойном оплодотворении, о триплоидном эндосперме и т.п.; это всё, конечно, верно – но немножко не о том. «С точки зрения биосферы» наиболее существенны, пожалуй, три свойства цветковых, позволившие им радикально изменить самую структуру мезозойских экосистем.
1/ Склонность к неотении (неотения – обретение организмом половозрелости на ранних (личиночных) стадиях своего индивидуального развития), то есть – в данном конкретном случае – к образованию травянистых форм, которые по сути дела есть древесные проростки, ставшие способными к размножению; напомним, что голосеменные «траву» не образуют вовсе – ни сейчас, ни в прошлом. Именно травянистые двудольные являются самыми агрессивными эксплерентами («сорными растениями»), мгновенно захватывающими места нарушений растительного покрова, а травянистые однодольные, склонные к образованию многолетней дернины, наиболее эффективно препятствуют эрозии.
2. Появление «мясистых» листьев с большим объемом фотосинтезирующей паренхимы, вызвавшее появление листогрызущих насекомых. Дело в том, что до того насекомые потребляли лишь генеративные органы, содержимое сосудов, даже древесину – но только не листья: в листе голосеменного (даже такого, как гинкго), попросту говоря, нечего есть. Ситуация менятся лишь с появлением цветковых – объеденные листья и мины (ходы, проедаемые личинками насекомых в толще листа) известны с самого начала позднего мела (сеноманский век). На покрытосеменных сформировалась общирная фауна листогрызущих насекомых, и уже после этого они перешли на голосеменные, папоротники и хвощи.
3. Энтомофилия цветков, открывающая покрытосеменным путь к сложной коэволюции с насекомыми. Собственно говоря, сам цветок покрытосеменных, судя по всему, сформировался в процессе адаптаций к насекомоопылению для защиты завязи от повреждений ее опылителями.
Ангиоспермы впервые появляются в палеонтологической летописи в начале раннего мела, в неокоме; на протяжении барремского и аптского веков они представлены единичными находками, в альбе идет стремительное нарастание их числа, а уже в самом начале позднего мела, в сеномане, цветковые составляют большую часть ископаемых флор – то есть наступает «кайнофит» (рисунок 50). Среднемеловая экспансия цветковых была столь стремительной (она фактически уложилась в ничтожный по геологический меркам отрезок от 115 до 108 млн лет), что Дарвин в свое время в сердцах назвал этот эпизод палеонтологической истории «отвратительной тайной – abominable mistery». Делались попытки объяснить это тем, что цветковые на самом-то деле появились на Земле давным-давно, минимум в юре – просто они обитали лишь в горах (не попадая по этой причине в палеонтологическую летопись), а в середине мела заселили, наконец, приводные местообитания, благоприятствующие попаданию в захоронения. Вообще такой «период скрытого существования» сбрасывать со счетов никак нельзя – но в данном случае эта гипотеза не проходит: дело в том, что пыльца покрытосеменных (которая, ясное дело, летает везде, и ее не «упрячешь» в горы) появляется в летописи одновременно с их макроостатками. По всем признакам, цветковые действительно возникли в начале мела, причем именно в приводных местообитаниях.
РИСУНОК 50. Распределение в меловых отложениях основных типов пыльцы и листьев покрытосеменных.
В 70-х годах советские палеоэнтомологи исследовали множество местонахождений меловых насекомых в Сибири и Монголии, «закрасив» тем самым самое крупное «белое пятно» в геологической истории класса. Оказалось, что у насекомых график, который отражает долю вымерших таксонов в фаунах разных времен, ведет себя довольно неожиданным образом (рисунок 51). Доля вымерших семейств мало меняется на протяжении большей части мезозоя, и на границе юры и мела составляет около 60% ; затем, в течении раннего мела, доля эта стремительно падает от 60 до 20% , в позднем мелу это падение резко замедляется (от 20 до 5%), а к началу кайнозоя кривая вновь «выходит на плато», только уже в районе 5%. Иными словами, у насекомых самая резкая фаунистическая смена, связанная с массовым вымиранием мезозойских групп, приходится не на конец мела, а на его середину – в точности как у растений. На этом основании В.В. Жерихин (1978) предложил следующую модель среднемелового биоценотического кризиса.
РИСУНОК 51. Эволюция насекомых в мезозое: процент вымерших семейств от общего числа семейств насекомых в позднемезозойских и кайнозойских фаунах.
Покрытосеменные возникли в начале мелового периода, первоначально как ценофобы – растения, не входящие в закономерные, сложившиеся в длительной коэволюции сукцессионные ряды [69]. Они росли в качестве «сорной» растительности по свободным от других растений участкам (на речных отмелях, береговых оползнях, гарях), которые соседствовали с гораздо более обширными участками, занятыми зрелыми сообществами мезофитной растительности. Покрытосеменные были исходно энтомофильными и смогли сформироваться лишь в окружении энтомофильных же голосеменных (таких, как беннетиты, саговники, кейтониевые), используя уже сложившийся к тому времени комплекс их опылителей. Но если покрытосеменные оказались среди пионерных растений мезофита единственными энтомофилами (а, судя по всему, так оно и было), то это должно было дать им гиганские преимущества: именно для локально распределенных, не образующих сплошных зарослей пионеров насекомоопыление особенно выгодно; впрочем, возможно, имело место и прямое конкурентное вытеснение покрытосеменными энтомофильных голосеменных из пионерных сообществ – за счет более эффективного привлечения насекомых, лучшей защиты от повреждения завязи опылителями и т.д.
Основой стремительной среднемеловой экспансии цветковых стало то, что им удалось закрепиться в качестве нормальной пионерной растительности – для чего решающими факторами стали их исходная энтомофильность и наличие среди них травянистых форм (что во много раз убыстряет зарастание поврежденных участков). Покрытосеменные не пытались потеснить всю мезофитную растительность, что было абсолютно нереально; они «всего-навсего» конкурентно вытеснили прежних пионеров – и тем самым блокировали все последующие стадии мезофитной экогенетической сукцессии. Разрушение существовавших в то время закономерных сукцессионных рядов (попросту говоря – мезофитная растительность продолжала существовать там, где она существует, но потеряла способность восстанавливаться после экзогенных нарушений), вызвало полный развал мезозойских наземных экосистем и массовое вымирание входивших в них животных; наиболее интенсивно эта деструкция шла в альбе (захватывая конец апта и начало сеномана).
В.В. Жерихин (1978) обратил внимание на любопытнейшую черту энтомофаун этого времени: многие семейства, характерные для мезозоя, обнаруживаются в них в последний раз, тогда как сколько-нибудь заметного появления новых групп не отмечается, и при этом появляется целый ряд представителей реликтовых для того времени (раннемезозойских и даже позднепалеозойских) групп. Это «всплывание реликтов» считают одним из наиболее ярких симптомов кризиса, охватившего среднемеловые сообщества: вымирание доминирующих раннемеловых групп (наиболее тесно связанных с существующей сукцессионной системой) создает экологический вакуум, который ненадолго заполняют древние формы, оттесненные к тому времени «на обочину жизни».