Страница:
Итак, подтвердив и развив теорию Агассиса, геологи оказались перед лицом проблемы – в чем же причина оледенений? Что вызывало рост ледниковых покровов прошлого, и почему, распространившись почти на треть суши, они вдруг начинали отступать? «Астрономические» гипотезы усматривали причину этих изменений в периодическом сокращении количества тепла, поступающего на Землю от Солнца. Другая группа гипотез акцентировала внимание на том, что оледенение – лишь одно из звеньев глобальной климатической системы, и система «ледниковый щит – океан – атмосфера» работает как единая гигантская машина; суть дела, заключали они, не в количестве тепла, поступающем на Землю, а в том, насколько равномерно это тепло распределено по поверхности планеты.
Солярная гипотеза, предполагающая периодические падения светимости Солнца, не имеет ныне сторонников: согласно мнению астрофизиков, звезды спектрального класса G-2, к которому относится наше Солнце, к подобным глупым шуткам совершенно не склонны. Зато весьма популярна теория астронома М. Миланковича (1924), связывающая оледенения с изменениями летней инсоляции (поступления солнечной радиации) в высоких широтах обеих полушарий, что, в свою очередь, обусловлено циклическими изменениям трех параметров орбитального движения Земли вокруг Солнца (вариациями наклона земной оси и пр.). Сделав поправки на эффекты менявшегося альбедо (отражательной способности земной поверхности), он рассчитал сдвиги в географическом положении границ ледниковых покровов за последний миллион лет, которые неплохо совпали с периодизацией европейских оледенений. Как на инсоляционных кривых Миланковича, так и на палеоклиматическом графике Пенка и Брюкнера ледниковые эпохи запечатлелись в виде коротких резких пульсаций, отделенных друг от друга длинными интервалами, и при этом «Великое межледниковье» (между минделем и риссом) занимает на графике то же место, что и предсказанный Миланковичем длительный теплый интервал (рисунок 59). Впоследствии картина оказалась гораздо более сложной, чем это представлялось в тридцатые-сороковые годы, однако ныне существование 100 000-летних климатических циклов, порождаемых орбитальными возмущениями, имеет вполне солидное обоснование.
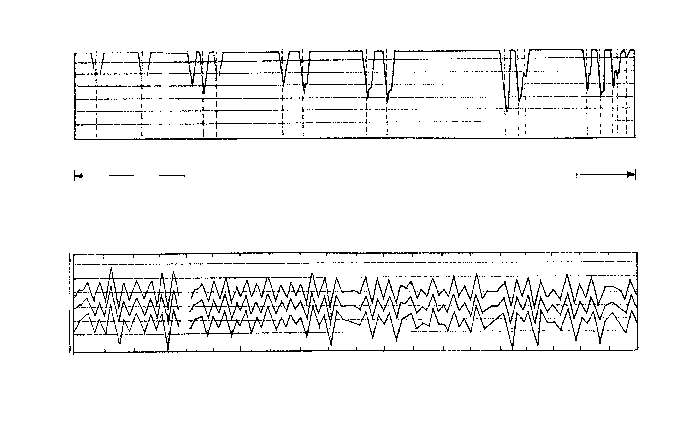
ИСУНОК 59. Проверка теории Миланковича: сопоставление климатической истории Европы (вверху) с инсоляционными кривыми Миланковича, рассчитанными для 55, 60 и 65( (внизу).
Теория Миланковича (в ее современных вариантах) удовлетворительно описывает динамику похолоданий и потеплений внутри ледникового периода, однако, к сожалению, не отвечает на вопрос о наступлении самого этого периода: вполне очевидно, что вся эта астрономическая циклика была точно такой же и в предшествующие плейстоцену эпохи, но никаких оледенений при этом не порождала. Поэтому с середины 50-х годов стал расти интерес к «земным» гипотезам оледенений, переносящим акцент на динамические взаимодействия в системе «оледенение – океан – атмосфера»; мы уже упоминали об «обратных парниковых эффектах», порождаемых изменениями атмосферного соотношения CO2/O2, и о работе морских течений при различном расположении материков.
Одной из самых интересных представляется гипотеза климатологов М. Юинга и У. Донна (1956). Задавшись вопросом – а почему оледенение не возникает сейчас, когда температурные условия Арктики вроде бы вполне тому благоприятствуют, – они сочли это следствием дефицита осадков [80]. Главный тезис их гипотезы: решающее условие возникновения оледенения в Арктике – усиление притока несущих влагу воздушных масс и усиление снегопадов; от этого ледник начинает нарастать, альбедо увеличивается, температура падает... ну, дальше – ясно. Вопрос: что же за фактор повышал влажность в арктических широтах? Ответ: освобождение Ледовитого океана от его ледового панциря в результате усиления притока теплой воды из Северной Атлантики. При отсутствии покрова морских льдов этот океан должен становиться мощнейшим испарителем, воздух над ним – «заряжаться» водяным паром, а интенсивность снегопадов на окружающей сушей – резко возрастать; рост альбедо доводит падение температуры до ледниковой эпохи. А вот дальше – самое интересное; в некоторый момент похолодание достигает той точки, когда Ледовитый океан вновь замерзает, и тогда начинается дегляциация: потеряв главный источник атмосферного питания, ледниковые покровы начинают «съеживаться». Ледник тает, и при этом уровень океана повышается, ветви теплого Северо-Атлантического течения вновь прорываются в Арктику, растапливают ее морские льды – и цикл начинается по новой.
Существование этой парадоксальной автоколебательной системы, в которой оледенение порождается потеплением, а дегляциация – похолоданием, нашло недавно косвенное подтверждение при изучении донных отложений Атлантики: оказалось, что в плейстоцене Гольфстрим периодически исчезал; при этом выяснилось, что усиление Гольфстрима действительно совпадает с периодами похолоданий, и наоборот. Вообще один из главных вопросов, на которые приходится отвечать «земным» гипотезам (подразумевающим примерное постоянство инсоляционного баланса планеты) – каким образом прекращается разрастание ледника, процесс, который по идее должен идти с положительной обратной связью? Ведь рост ледника приводит (через увеличение альбедо) к падению температуры, что еще увеличивает ледник – и так до тех пор, пока льдом не покроется вся планета... Один из наиболее убедительных ответов состоит в том, что по достижении ледником некого порогового размера над ним (именно из-за высокого альбедо) возникает постоянно действующий антициклон (область высокого атмосферного давления), который усиливается по мере роста ледника и в конце концов лишает его «питания» – осадков. Таким образом, климат перигляциальных (окружающих ледник) территорий должен быть холодным и сухим – что полностью подтверждается палеонтологическими данными. В этих ландшафтах складываются весьма специфическая фаунистическая группировка с сочетанием криофильных (холодолюбивых) и ксерофильных (сухолюбивых) элементов, которую А.Я. Тугаринов (1929) назвал тундростепной; типично тундровые (влаголюбивые) элементы встречаются здесь лишь близ водотоков.
Сейчас группировки, сходные с перигляциальными, сохранились в виде так называемых реликтовых степей – островков среди таежного и лесотундрового ландшафта, приуроченных к южным склонам гор северо-восточной Сибири и Аляски, а также в холодных засушливых высокогорьях Центральной Азии (здесь их называют «пастбищем яков»). Реликтовые степи, так же как и тундра, представляют собой безлесые сериальные стадии к хвойным таежным климаксам – соответственно, ксеро– и гидросерию; соотношение площадей, занимаемых в ареале сукцессионной системы сообществами ксеро– и гидросериального ряда, определяется климатической ситуацией. В перигляциальном ландшафте, где вся вода вымерзает на массе ледника (как в морозилке холодильника) и ситуация складывается фактически аридная, доминируют криоксерофильные тундростепи, а настоящие тундры существуют лишь в виде приводных сообществ. Когда же ледниковая эпоха заканчивается и наступает межледниковье, ситуация меняется на обратную. Ледник тает, высвобождая массы воды, и доминировать на осовобождающейся ото льда территории начинают сообщества гидроряда (тундра – это в некотором смысле чудовищно разросшееся таежное моховое болото), тундростепи же съеживаются до крохотных островков «реликтовых степей» на сухих прогреваемых южных склонах.
Тундростепь отличалась тем, что травяной ярус ее формировали в основном не мхи (как в тундре), а злаки; здесь складывался крайне криофильный вариант уже знакомого нам (по главе 13) травяного биома с его высокой биомассой пастбищных копытных и хищников – мамонтовой фауной. В ее составе были причудливо смешаны виды, приуроченные ныне к тундре (северный олень, овцебык, леминги), к степям (сайгак, лошадь, верблюд, бизон, суслики), а также виды, характерные лишь для этого сообщества и исчезнувшие вместе с ним (мамонт, шерстистый носорог, саблезубый тигр – смилодон, гигантская гиена – рисунок 60). Исчезновение мамонтовой фауны, произошедшее в начале голоцена, одни исследователи связывают с климатическими изменениями, другие же считают этих животных жертвами человека («охотников на мамонтов»). Сторонники гипотезы «антропогенного вымирания» резонно указывают, что все предыдущие межледниковья, когда еще не было человека, криофильная мамонтовая фауна пережила вполне спокойно. Сторонники гипотезы «климатических воздействий» – опять-таки резонно – возражают, что голоценовое вымирание было наиболее масштабным не в относительно густо заселенной Евразии, а в практически безлюдной в те времена Северной Америке (человек проник сюда лишь около 10-12 тысяч лет назад из Азии через Берингов пролив); на прародине же человечества – в африканских саваннах – никаких вымираний вообще не было. Кроме того, вымирание захватило не только крупных травоядных и хищников, но и целую кучу маммальной мелочи, которая никак не могла быть для кроманьонцев ни добычей, ни врагами, подлежащими целенаправленному уничтожению.
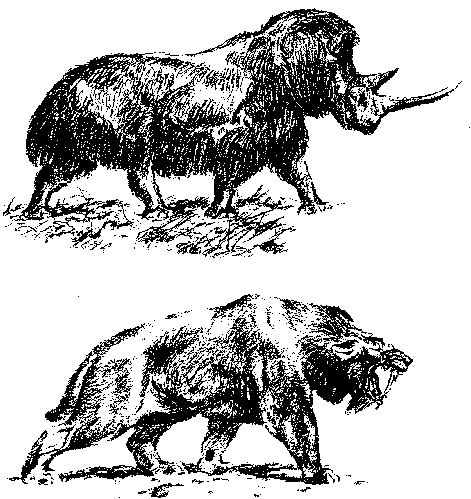
РИСУНОК 60. Вымершие представители мамонтовой фауны: (а) – шерстистый носорог, (б) – гигантская саблезубая кошка смилодон.
Представляется, что наиболее близок к истине В.В. Жерихин (1993): «Каждый тип травяного биома производен от вполне определенной сукцессионной системы с лесным климаксом [в случае тундростепи – от тайги: К.Е.]. При вторичном сильном сокращении площади травяных сообществ они могут полностью утратить комплекс [поддерживающих их] крупных травоядных, а тем самым и эндогенную стабильность. В этом случае они могут вновь приобрести статус сериальных. Ярким примером могут служить современные реликтовые тундростепи, сохранившиеся в таежных сукцессионных системах после полного исчезновения тундростепного биома.» В момент таяния ледника и резкого увлажнения климата расширяются моховые тундры и сокращаются злаковые тундростепи, служащие пастбищем для мамонтовой фауны. Дополнительные неприятности для популяций этих животных создает то, что тундростепной ландшафт оказывается «нарезанным» на «острова»: и из теоретической экологии, и из современной практики заповедного дела известно, что для крупных животных несколько мелких резерватов хуже одного крупного (равного им по площади). Вот в этих-то, кризисных, условиях человек мог нанести мамонтовой фауне последний удар: выборочно уничтожая крупных копытных, он значительно ускорил превращение тундростепей в лесные сообщества – а дальше процесс этот пошел неостановимо, с положительной обратной связью, пока не исчез весь этот фаунистический комплекс (хотя часть его сохраняется ныне в фауне тундр и степей). Отметим, что дольше всего мамонт выжил на острове Врангеля (открытый недавно карликовый подвид, около 1,5 м в холке, вымер 5 тыс. лет назад – против 10-12 тыс. лет на континенте), где и поныне широко распространены реликтовые степи.
Самое же интересное – что итоговое воздействие катастрофических (по любым меркам) плейстоценовых оледенений на биоту Северного полушария оказалось совершенно ничтожным. Да, вымерло некоторое количество млекопитающих из мамонтовой фауны, но во-первых темпы этого вымирания не превышают средних по кайнозою, а во-вторых, как мы теперь знаем, мамонтовая фауна вымерла скорее в результате прекращения оледенения. Известен лишь один вымерший вид четвертичных насекомых (если не считать гигантского кожного овода, паразитировавший на мамонте, и нескольких видов североамериканских жуков-навозников – те исчезли вместе со своими хозяевами и прокормителями); что же касается растений, то они, похоже, не пострадали вовсе. Создается отчетливое впечатление, что в плейстоцене менялось лишь географическое распространение экосистем (широколиственные леса временно отступали к югу, а на севере изменялось соотношение площадей, занятых сообществами гидро– и ксеросериального ряда) и отдельных видов (в перигляциальных сообществах Европы появлялись жуки, ограниченные ныне степями Якутии и Тибетом) [81]. Все это лишний раз свидетельствует о том, что экосистемы в норме обладают колоссальной устойчивостью, и разрушить их внешними воздействиями – даже катастрофическими – практически невозможно. Особенно замечательно плейстоценовая ситуация смотрится на фоне «тихих» внутрисистемных кризисов вроде среднемеловой ангиоспермизации – заведомо не связанной ни с какими импактами и драматическими перестройками климата, но вызвавшей обвальные вымирания в наземных и пресноводных сообществах.
С другой стороны, влияние плейстоценовых оледенений на климат планеты отнюдь не ограничивалось высокими ее широтами. Разрастание ледниковых щитов близ полюса тут же аукалось на экваторе невиданным иссушением тропического пояса: установлено, что дождевые тропические леса Южной Америки периодически съеживались до нескольких десятков крохотных пятачков-"резерватов" в среднем течении Амазонки, а всю эту территорию занимали сухие саванны. Более того: есть серьезные основания полагать, что пустыни наиболее распространенного ныне на Земле средиземноморского типа [82]возникли лишь в плейстоцене. Если пустыни берегового и центральноазиатского типа, грубо говоря, являются пустынями всегда, то средиземноморские пустыни становятся таковыми временно, в зависимости от глобальной климатической обстановки, т.е. от взаимодействий в системе «оледенение – атмосфера – океан». Например, иссушение Сахары (наступление песков на саванну) идет буквально на наших глазах: первые европейские путешественники застали озеро Чад настоящим внутренним морем, а в реках нагорья Тибести, что в самом центре Сахары, еще в 20-е годы нашего века жили крокодилы.
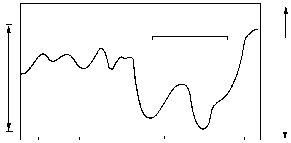
РИСУНОК 61. Климат последнего тысячелетия.
Многие из этих климатических изменений повторялись «в миниатюре» на памяти человечества. На рисунке 61 представлена кривая зимних температур в Европе за последнее тысячелетие. Во время так называемого «Малого ледникового периода» (1450-1850 гг) ледники повсеместно наступали, и их размеры превосходили современные (снежный покров появлялся, например, в горах Эфиопии, где его ныне не бывает). Во время же предшествовавшего тому Атлантического оптимума (900-1300 гг) ледники сократились, и климат был заметно мягче нынешнего (вспомните: именно в эти времена викинги назвали Гренландию «Зеленой страной»); следствием потепления в высоких широтах стало увеличение количества осадков, выпадающих в аридном поясе.
Итак, на Севере стало тепло и, как писано в одном хорошем романе о викингах, «наступили времена изобилия и достатка, когда собирался такой прекрасный урожай ржи, а улов сельди был настолько велик, что большинство людей легко могли прокормить себя [что в Средневековье случалось нечасто – К.Е.]». Итог этого «изобилия и достатка» – норманская экспансия в Европе: «датская дань» [83]в Англии, варяжские дружины при всех состоятельных государях, колонизация Исландии и Гренландии, плавания в Америку. То же самое – на Юге, где стало влажно, пустыня обратилась в степь, и такое же «изобилие и достаток» посетило кочевые скотоводческие народы Центральной Азии; итог – «Монгольское нашествие» от Китая до Адриатики. На это же время приходится и расцвет городских цивилизаций в африканских саваннах – Канем, Гао, Гана, Мали, Ифе... О климатических воздействиях на судьбы человеческих цивилизаций можно говорить много, однако тут я уже явно начинаю отбивать хлеб у историков; как раз в такие моменты Шахерезада «прекращала дозволенные ей речи» – и была совершенно права.
14-а (дополнительная). Историческая биогеография. Викариантная модель и концепция «оттесненных реликтов». Фитоспрединг.
Солярная гипотеза, предполагающая периодические падения светимости Солнца, не имеет ныне сторонников: согласно мнению астрофизиков, звезды спектрального класса G-2, к которому относится наше Солнце, к подобным глупым шуткам совершенно не склонны. Зато весьма популярна теория астронома М. Миланковича (1924), связывающая оледенения с изменениями летней инсоляции (поступления солнечной радиации) в высоких широтах обеих полушарий, что, в свою очередь, обусловлено циклическими изменениям трех параметров орбитального движения Земли вокруг Солнца (вариациями наклона земной оси и пр.). Сделав поправки на эффекты менявшегося альбедо (отражательной способности земной поверхности), он рассчитал сдвиги в географическом положении границ ледниковых покровов за последний миллион лет, которые неплохо совпали с периодизацией европейских оледенений. Как на инсоляционных кривых Миланковича, так и на палеоклиматическом графике Пенка и Брюкнера ледниковые эпохи запечатлелись в виде коротких резких пульсаций, отделенных друг от друга длинными интервалами, и при этом «Великое межледниковье» (между минделем и риссом) занимает на графике то же место, что и предсказанный Миланковичем длительный теплый интервал (рисунок 59). Впоследствии картина оказалась гораздо более сложной, чем это представлялось в тридцатые-сороковые годы, однако ныне существование 100 000-летних климатических циклов, порождаемых орбитальными возмущениями, имеет вполне солидное обоснование.
ИСУНОК 59. Проверка теории Миланковича: сопоставление климатической истории Европы (вверху) с инсоляционными кривыми Миланковича, рассчитанными для 55, 60 и 65( (внизу).
Теория Миланковича (в ее современных вариантах) удовлетворительно описывает динамику похолоданий и потеплений внутри ледникового периода, однако, к сожалению, не отвечает на вопрос о наступлении самого этого периода: вполне очевидно, что вся эта астрономическая циклика была точно такой же и в предшествующие плейстоцену эпохи, но никаких оледенений при этом не порождала. Поэтому с середины 50-х годов стал расти интерес к «земным» гипотезам оледенений, переносящим акцент на динамические взаимодействия в системе «оледенение – океан – атмосфера»; мы уже упоминали об «обратных парниковых эффектах», порождаемых изменениями атмосферного соотношения CO2/O2, и о работе морских течений при различном расположении материков.
Одной из самых интересных представляется гипотеза климатологов М. Юинга и У. Донна (1956). Задавшись вопросом – а почему оледенение не возникает сейчас, когда температурные условия Арктики вроде бы вполне тому благоприятствуют, – они сочли это следствием дефицита осадков [80]. Главный тезис их гипотезы: решающее условие возникновения оледенения в Арктике – усиление притока несущих влагу воздушных масс и усиление снегопадов; от этого ледник начинает нарастать, альбедо увеличивается, температура падает... ну, дальше – ясно. Вопрос: что же за фактор повышал влажность в арктических широтах? Ответ: освобождение Ледовитого океана от его ледового панциря в результате усиления притока теплой воды из Северной Атлантики. При отсутствии покрова морских льдов этот океан должен становиться мощнейшим испарителем, воздух над ним – «заряжаться» водяным паром, а интенсивность снегопадов на окружающей сушей – резко возрастать; рост альбедо доводит падение температуры до ледниковой эпохи. А вот дальше – самое интересное; в некоторый момент похолодание достигает той точки, когда Ледовитый океан вновь замерзает, и тогда начинается дегляциация: потеряв главный источник атмосферного питания, ледниковые покровы начинают «съеживаться». Ледник тает, и при этом уровень океана повышается, ветви теплого Северо-Атлантического течения вновь прорываются в Арктику, растапливают ее морские льды – и цикл начинается по новой.
Существование этой парадоксальной автоколебательной системы, в которой оледенение порождается потеплением, а дегляциация – похолоданием, нашло недавно косвенное подтверждение при изучении донных отложений Атлантики: оказалось, что в плейстоцене Гольфстрим периодически исчезал; при этом выяснилось, что усиление Гольфстрима действительно совпадает с периодами похолоданий, и наоборот. Вообще один из главных вопросов, на которые приходится отвечать «земным» гипотезам (подразумевающим примерное постоянство инсоляционного баланса планеты) – каким образом прекращается разрастание ледника, процесс, который по идее должен идти с положительной обратной связью? Ведь рост ледника приводит (через увеличение альбедо) к падению температуры, что еще увеличивает ледник – и так до тех пор, пока льдом не покроется вся планета... Один из наиболее убедительных ответов состоит в том, что по достижении ледником некого порогового размера над ним (именно из-за высокого альбедо) возникает постоянно действующий антициклон (область высокого атмосферного давления), который усиливается по мере роста ледника и в конце концов лишает его «питания» – осадков. Таким образом, климат перигляциальных (окружающих ледник) территорий должен быть холодным и сухим – что полностью подтверждается палеонтологическими данными. В этих ландшафтах складываются весьма специфическая фаунистическая группировка с сочетанием криофильных (холодолюбивых) и ксерофильных (сухолюбивых) элементов, которую А.Я. Тугаринов (1929) назвал тундростепной; типично тундровые (влаголюбивые) элементы встречаются здесь лишь близ водотоков.
Сейчас группировки, сходные с перигляциальными, сохранились в виде так называемых реликтовых степей – островков среди таежного и лесотундрового ландшафта, приуроченных к южным склонам гор северо-восточной Сибири и Аляски, а также в холодных засушливых высокогорьях Центральной Азии (здесь их называют «пастбищем яков»). Реликтовые степи, так же как и тундра, представляют собой безлесые сериальные стадии к хвойным таежным климаксам – соответственно, ксеро– и гидросерию; соотношение площадей, занимаемых в ареале сукцессионной системы сообществами ксеро– и гидросериального ряда, определяется климатической ситуацией. В перигляциальном ландшафте, где вся вода вымерзает на массе ледника (как в морозилке холодильника) и ситуация складывается фактически аридная, доминируют криоксерофильные тундростепи, а настоящие тундры существуют лишь в виде приводных сообществ. Когда же ледниковая эпоха заканчивается и наступает межледниковье, ситуация меняется на обратную. Ледник тает, высвобождая массы воды, и доминировать на осовобождающейся ото льда территории начинают сообщества гидроряда (тундра – это в некотором смысле чудовищно разросшееся таежное моховое болото), тундростепи же съеживаются до крохотных островков «реликтовых степей» на сухих прогреваемых южных склонах.
Тундростепь отличалась тем, что травяной ярус ее формировали в основном не мхи (как в тундре), а злаки; здесь складывался крайне криофильный вариант уже знакомого нам (по главе 13) травяного биома с его высокой биомассой пастбищных копытных и хищников – мамонтовой фауной. В ее составе были причудливо смешаны виды, приуроченные ныне к тундре (северный олень, овцебык, леминги), к степям (сайгак, лошадь, верблюд, бизон, суслики), а также виды, характерные лишь для этого сообщества и исчезнувшие вместе с ним (мамонт, шерстистый носорог, саблезубый тигр – смилодон, гигантская гиена – рисунок 60). Исчезновение мамонтовой фауны, произошедшее в начале голоцена, одни исследователи связывают с климатическими изменениями, другие же считают этих животных жертвами человека («охотников на мамонтов»). Сторонники гипотезы «антропогенного вымирания» резонно указывают, что все предыдущие межледниковья, когда еще не было человека, криофильная мамонтовая фауна пережила вполне спокойно. Сторонники гипотезы «климатических воздействий» – опять-таки резонно – возражают, что голоценовое вымирание было наиболее масштабным не в относительно густо заселенной Евразии, а в практически безлюдной в те времена Северной Америке (человек проник сюда лишь около 10-12 тысяч лет назад из Азии через Берингов пролив); на прародине же человечества – в африканских саваннах – никаких вымираний вообще не было. Кроме того, вымирание захватило не только крупных травоядных и хищников, но и целую кучу маммальной мелочи, которая никак не могла быть для кроманьонцев ни добычей, ни врагами, подлежащими целенаправленному уничтожению.
РИСУНОК 60. Вымершие представители мамонтовой фауны: (а) – шерстистый носорог, (б) – гигантская саблезубая кошка смилодон.
Представляется, что наиболее близок к истине В.В. Жерихин (1993): «Каждый тип травяного биома производен от вполне определенной сукцессионной системы с лесным климаксом [в случае тундростепи – от тайги: К.Е.]. При вторичном сильном сокращении площади травяных сообществ они могут полностью утратить комплекс [поддерживающих их] крупных травоядных, а тем самым и эндогенную стабильность. В этом случае они могут вновь приобрести статус сериальных. Ярким примером могут служить современные реликтовые тундростепи, сохранившиеся в таежных сукцессионных системах после полного исчезновения тундростепного биома.» В момент таяния ледника и резкого увлажнения климата расширяются моховые тундры и сокращаются злаковые тундростепи, служащие пастбищем для мамонтовой фауны. Дополнительные неприятности для популяций этих животных создает то, что тундростепной ландшафт оказывается «нарезанным» на «острова»: и из теоретической экологии, и из современной практики заповедного дела известно, что для крупных животных несколько мелких резерватов хуже одного крупного (равного им по площади). Вот в этих-то, кризисных, условиях человек мог нанести мамонтовой фауне последний удар: выборочно уничтожая крупных копытных, он значительно ускорил превращение тундростепей в лесные сообщества – а дальше процесс этот пошел неостановимо, с положительной обратной связью, пока не исчез весь этот фаунистический комплекс (хотя часть его сохраняется ныне в фауне тундр и степей). Отметим, что дольше всего мамонт выжил на острове Врангеля (открытый недавно карликовый подвид, около 1,5 м в холке, вымер 5 тыс. лет назад – против 10-12 тыс. лет на континенте), где и поныне широко распространены реликтовые степи.
Самое же интересное – что итоговое воздействие катастрофических (по любым меркам) плейстоценовых оледенений на биоту Северного полушария оказалось совершенно ничтожным. Да, вымерло некоторое количество млекопитающих из мамонтовой фауны, но во-первых темпы этого вымирания не превышают средних по кайнозою, а во-вторых, как мы теперь знаем, мамонтовая фауна вымерла скорее в результате прекращения оледенения. Известен лишь один вымерший вид четвертичных насекомых (если не считать гигантского кожного овода, паразитировавший на мамонте, и нескольких видов североамериканских жуков-навозников – те исчезли вместе со своими хозяевами и прокормителями); что же касается растений, то они, похоже, не пострадали вовсе. Создается отчетливое впечатление, что в плейстоцене менялось лишь географическое распространение экосистем (широколиственные леса временно отступали к югу, а на севере изменялось соотношение площадей, занятых сообществами гидро– и ксеросериального ряда) и отдельных видов (в перигляциальных сообществах Европы появлялись жуки, ограниченные ныне степями Якутии и Тибетом) [81]. Все это лишний раз свидетельствует о том, что экосистемы в норме обладают колоссальной устойчивостью, и разрушить их внешними воздействиями – даже катастрофическими – практически невозможно. Особенно замечательно плейстоценовая ситуация смотрится на фоне «тихих» внутрисистемных кризисов вроде среднемеловой ангиоспермизации – заведомо не связанной ни с какими импактами и драматическими перестройками климата, но вызвавшей обвальные вымирания в наземных и пресноводных сообществах.
С другой стороны, влияние плейстоценовых оледенений на климат планеты отнюдь не ограничивалось высокими ее широтами. Разрастание ледниковых щитов близ полюса тут же аукалось на экваторе невиданным иссушением тропического пояса: установлено, что дождевые тропические леса Южной Америки периодически съеживались до нескольких десятков крохотных пятачков-"резерватов" в среднем течении Амазонки, а всю эту территорию занимали сухие саванны. Более того: есть серьезные основания полагать, что пустыни наиболее распространенного ныне на Земле средиземноморского типа [82]возникли лишь в плейстоцене. Если пустыни берегового и центральноазиатского типа, грубо говоря, являются пустынями всегда, то средиземноморские пустыни становятся таковыми временно, в зависимости от глобальной климатической обстановки, т.е. от взаимодействий в системе «оледенение – атмосфера – океан». Например, иссушение Сахары (наступление песков на саванну) идет буквально на наших глазах: первые европейские путешественники застали озеро Чад настоящим внутренним морем, а в реках нагорья Тибести, что в самом центре Сахары, еще в 20-е годы нашего века жили крокодилы.
РИСУНОК 61. Климат последнего тысячелетия.
Многие из этих климатических изменений повторялись «в миниатюре» на памяти человечества. На рисунке 61 представлена кривая зимних температур в Европе за последнее тысячелетие. Во время так называемого «Малого ледникового периода» (1450-1850 гг) ледники повсеместно наступали, и их размеры превосходили современные (снежный покров появлялся, например, в горах Эфиопии, где его ныне не бывает). Во время же предшествовавшего тому Атлантического оптимума (900-1300 гг) ледники сократились, и климат был заметно мягче нынешнего (вспомните: именно в эти времена викинги назвали Гренландию «Зеленой страной»); следствием потепления в высоких широтах стало увеличение количества осадков, выпадающих в аридном поясе.
Итак, на Севере стало тепло и, как писано в одном хорошем романе о викингах, «наступили времена изобилия и достатка, когда собирался такой прекрасный урожай ржи, а улов сельди был настолько велик, что большинство людей легко могли прокормить себя [что в Средневековье случалось нечасто – К.Е.]». Итог этого «изобилия и достатка» – норманская экспансия в Европе: «датская дань» [83]в Англии, варяжские дружины при всех состоятельных государях, колонизация Исландии и Гренландии, плавания в Америку. То же самое – на Юге, где стало влажно, пустыня обратилась в степь, и такое же «изобилие и достаток» посетило кочевые скотоводческие народы Центральной Азии; итог – «Монгольское нашествие» от Китая до Адриатики. На это же время приходится и расцвет городских цивилизаций в африканских саваннах – Канем, Гао, Гана, Мали, Ифе... О климатических воздействиях на судьбы человеческих цивилизаций можно говорить много, однако тут я уже явно начинаю отбивать хлеб у историков; как раз в такие моменты Шахерезада «прекращала дозволенные ей речи» – и была совершенно права.
14-а (дополнительная). Историческая биогеография. Викариантная модель и концепция «оттесненных реликтов». Фитоспрединг.
Биогеография – наука о закономерностях географического распространения живых существ. Вполне очевидно, что всякое живое существо экологически связано с определенным типом ландшафта, а если смотреть шире – с одной из природных (ландшафтно-климатических) зон: белый медведь – с арктическими льдами, лось – с тайгой, верблюд – с пустыней. С другой стороны, ареал (область распространения) некоей группы организмов в той или иной степени отражает и историю его формирования: район происхождения, время расселения, действие преград (неполнозубые, широконосые обезьяны и кавиморфные грызуны ограничены Южной Америкой, что отражает долгую изоляцию этого континента, нарушенную лишь недавно возникновением Панамского перешейка). В соответствии с этими двумя аспектами биогеографию подразделяют на ландшафтную (которая является фактически разделом экологии) и историческую. Например, верблюды распространены ныне в пустынях Азии – Центральной (двугорбый бактриан) и Передней (одногорбый дромадер); так вот, ландшафтная биогеография призвана ответить на вопрос о том, почему верблюды живут в зоне пустынь (а не в субтропических лесах или, скажем, в тайге), а историческая – почему верблюды распространены лишь в Азии и отсутствуют в Австралии и Северной Америке (где есть вполне вроде бы подходящие для них пустыни).
Возможных вариантов тут два: то ли верблюды этих континентов по каким-то причинам не достигли, то ли они здесь раньше жили, но вымерли. Но можно ли прийти к определенным историческим выводам, исходя лишь из особенностей их современного распространения? Оказывается, можно. Ближайшие родственники верблюдов – ламы – распространены в Южной Америке, так что каким бы путем ни расселялись мозоленогие (хоть из Азии в Южную Америку, хоть наоборот – из Южной Америки в Азию), миновать Северную Америку они никак не могли. Соответственно, на этом материке мозоленогие наверняка жили, но вымерли, а вот Австралии этот подотряд, видимо, не достиг – так же, как все прочие плацентарные млекопитающие (кроме летучих мышей). Ископаемые находки подтверждают эту реконструкцию (верблюды произошли в Северной Америке и исчезли там совсем недавно), однако так бывает далеко не всегда.
Давайте попытаемся реконструировать историю сумчатых, исходя лишь из их современного распространения – Австралия и Южная Америка (опоссум проник в Северную Америку из Южной недавно, по панамскому мосту). В прежние годы, когда положение материков принималось неизменным, приходилось предполагать, что сумчатые возникли в Северном полушарии, затем проникли в Австралию (из Азии) и Южную Америку (из Северной), после чего вымерли на северных материках. По нынешним временам же логично предположить, что группа эта никогда не проникала на территорию Арктогеи (ну, кроме опоссума), будучи на протяжении всей своей истории изолированной на южных материках – так же, как, например, «южноамериканские копытные». Она возникла в конце мела, когда фрагмент Гондваны, объединявший Южную Америку, Австралию и Антарктиду, был уже изолирован от остального мира, и расселилась по нему до эоцена, когда от него отделилась Австралия. Ископаемые находки, однако, рисуют иную картину. Сумчатые действительно появились в Южной Америке в позднем мелу, а потом через Антарктиду проникли в Австралию; родиной их, однако, является Северная Америка, откуда они – через Гренландию – заселили также Европу, где дожили до неогена, так и не проникнув ни в Азию, ни в Африку (рисунок 62).
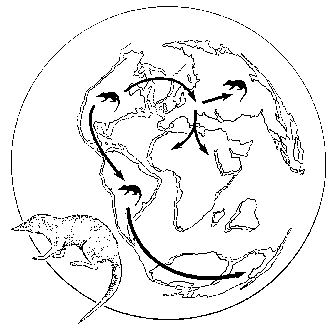
РИСУНОК 62. Географическая история сумчатых: пути расселения из центра происхождения (по палеонтологическим данным).
Рассмотренная ситуация наглядно демонстрирует два момента. Во-первых, насколько опасно в историко-биогеографических реконструкциях опираться лишь на современное распространение группы (что делается биогеографами сплошь и рядом). Во-вторых, насколько эти реконструкции зависят от концепций, господствующих в геологии – о корректности каковых концепций биогеографы (естественно!) профессионально судить не могут и вынуждены многое принимать «на веру». В этом смысле весьма показательна история изучения дизъюнктивных (разорванных) трансокеанических ареалов.
Многие животные и растения распространены лишь на южных оконечностях разобщенных ныне океанами материков Южного полушария (рисунок 63). Происхождение этих дизъюнкций издавна интриговало биогеографов. В XIX веке предполагали, что эти живые существа проникали из одной части их современного ареала в другую по неким «временным сухопутным мостам» – обширным участкам суши между континентами (расположение последних считалось тогда неизменным), которые впоследствии погрузились в океан. Типичный пример такого рода реконструкций – Лемурия, гипотетическая суша в Индийском океане, «созданная» биогеографами для того, чтобы напрямую «провести» из Африки в Индию лемуров, слонов, панголинов и т.п. Надо заметить, что сторонники этого направления быстро вошли во вкус и, по едкому замечанию Ч. Дарвина, принялись печь подобные «временные мосты», как блины.
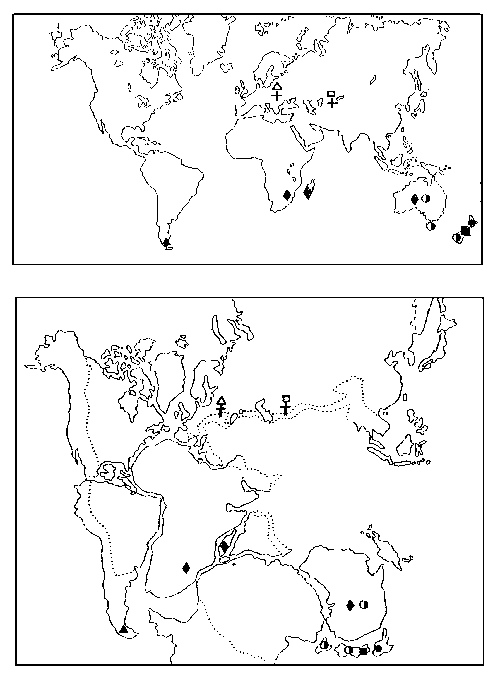
РИСУНОК 63. Пример южнополушарной дизъюнкции: распространение родов пауков-археид по современным материкам (а) и при нанесении его на карту Гондваны (б). Ископаемые рода (из эоценового Балтийского янтаря и из юры Казахстана) помечены крестиком.
Хотя ныне эти реконструкции представляют лишь исторический интерес (они, помимо всего прочего, противоречат концепции изостазии), рациональное зерно в их «констатирующей части» определенно содержалось. Так, ботаник Дж. Гукер (1860) на огромном материале продемонстрировал высокое единство биоты материков Южного полушария (вспомним нотофагусовые леса на юге Австралии, в Новой Зеландии и в Чили). Последователи Гукера предполагали автохтонное (на месте своего нынешнего распространения) развитие биоты Южных континентов – изолированных от Северных и соединенных между собою некими сухопутными связями; предполагалось, что одним из основных источников формирования этой биоты могла служить Антарктида – еще не покрытая тогда ледником и имевшая существенно больший размер, чем ныне. Итак, в рамках этой концепции основное внимание уделялось единству Южной биоты и обеспечивающим ее механизмам расселения.
Иной взгляд на природу южнополушарных дизъюнкций высказывал зоолог А. Уоллес (1876). Он полагал, что основные формообразовательные процессы идут на блоке Северных континентов, откуда более молодые и продвинутые формы животных и растений оттесняют более древних и архаичных все дальше к югу. Южные оконечности материков Южного полушария являются, таким образом, «последней линией обороны» этих реликтовых (сохранившихся от более древних эпох) групп, которые в прежние времена имели гораздо более широкое, обычно панконтинентальное, распространение. С этой точки зрения нет принципиальной разницы между трансокеаническими дизъюнкциями и дизъюнкциями, расположенными на одном материке (например, европейско-дальневосточными, как у жерлянок): и те, и другие возникают в результате сокращения исходного ареала и вымирания «соединительных звеньев». Концепцию Уоллеса (ее позднее назвали теорией оттесненных реликтов) подтверждали ископаемые находки – мнгие «крайне-южные» ныне животные и растения (араукарии, саговники, двоякодышащие рыбы, бескилевые птицы и т.д.) действительно обитали раньше на северных континентах. Пауки-археиды (рисунок 63), например, были первоначально открыты в ископаемом состоянии – в эоценовом балтийском янтаре, – и лишь спустя несколько десятков лет в Южном полушарии были найдены их современные представители. В рамках этой концепции (в отличие от Гукеровской) акцент переносился на архаичность и реликтовый характер Южной биоты.
Модель Уоллеса, хорошо обоснованная палеонтологическими данными и не вводящая таких «избыточных сущностей» (по Оккаму), как сухопутные межматериковые соединения непонятной геологической природы, долго считалась главенствующей. Однако с тех пор, как в геологии возобладала тектоника плит, среди биогеографов возродился интерес к Гукеровской модели: ведь теперь уже не было нужды воздвигать между материками какие-то «сухопутные мосты» – достаточно сдвинуть между собою сами материки. Эта модель позволяет рассматривать разнообразные южнополушарные дизьюнкции как прямой результат раскола Гондваны и сохранения на ее разошедшихся в результате континентального дрейфа фрагментах представителей некогда единой биоты этого мезозойского праматерика (рисунок 63, б). Гукеровские построения считают одним из основных источников наиболее популярной ныне биогеографической концепции – викариантной биогеографии: ее задача состоит в том, чтобы согласовать последовательность ветвления филогенетического древа таксона с рисуемой геофизиками последовательностью расхождения фрагментов Пангеи.
Возможных вариантов тут два: то ли верблюды этих континентов по каким-то причинам не достигли, то ли они здесь раньше жили, но вымерли. Но можно ли прийти к определенным историческим выводам, исходя лишь из особенностей их современного распространения? Оказывается, можно. Ближайшие родственники верблюдов – ламы – распространены в Южной Америке, так что каким бы путем ни расселялись мозоленогие (хоть из Азии в Южную Америку, хоть наоборот – из Южной Америки в Азию), миновать Северную Америку они никак не могли. Соответственно, на этом материке мозоленогие наверняка жили, но вымерли, а вот Австралии этот подотряд, видимо, не достиг – так же, как все прочие плацентарные млекопитающие (кроме летучих мышей). Ископаемые находки подтверждают эту реконструкцию (верблюды произошли в Северной Америке и исчезли там совсем недавно), однако так бывает далеко не всегда.
Давайте попытаемся реконструировать историю сумчатых, исходя лишь из их современного распространения – Австралия и Южная Америка (опоссум проник в Северную Америку из Южной недавно, по панамскому мосту). В прежние годы, когда положение материков принималось неизменным, приходилось предполагать, что сумчатые возникли в Северном полушарии, затем проникли в Австралию (из Азии) и Южную Америку (из Северной), после чего вымерли на северных материках. По нынешним временам же логично предположить, что группа эта никогда не проникала на территорию Арктогеи (ну, кроме опоссума), будучи на протяжении всей своей истории изолированной на южных материках – так же, как, например, «южноамериканские копытные». Она возникла в конце мела, когда фрагмент Гондваны, объединявший Южную Америку, Австралию и Антарктиду, был уже изолирован от остального мира, и расселилась по нему до эоцена, когда от него отделилась Австралия. Ископаемые находки, однако, рисуют иную картину. Сумчатые действительно появились в Южной Америке в позднем мелу, а потом через Антарктиду проникли в Австралию; родиной их, однако, является Северная Америка, откуда они – через Гренландию – заселили также Европу, где дожили до неогена, так и не проникнув ни в Азию, ни в Африку (рисунок 62).
РИСУНОК 62. Географическая история сумчатых: пути расселения из центра происхождения (по палеонтологическим данным).
Рассмотренная ситуация наглядно демонстрирует два момента. Во-первых, насколько опасно в историко-биогеографических реконструкциях опираться лишь на современное распространение группы (что делается биогеографами сплошь и рядом). Во-вторых, насколько эти реконструкции зависят от концепций, господствующих в геологии – о корректности каковых концепций биогеографы (естественно!) профессионально судить не могут и вынуждены многое принимать «на веру». В этом смысле весьма показательна история изучения дизъюнктивных (разорванных) трансокеанических ареалов.
Многие животные и растения распространены лишь на южных оконечностях разобщенных ныне океанами материков Южного полушария (рисунок 63). Происхождение этих дизъюнкций издавна интриговало биогеографов. В XIX веке предполагали, что эти живые существа проникали из одной части их современного ареала в другую по неким «временным сухопутным мостам» – обширным участкам суши между континентами (расположение последних считалось тогда неизменным), которые впоследствии погрузились в океан. Типичный пример такого рода реконструкций – Лемурия, гипотетическая суша в Индийском океане, «созданная» биогеографами для того, чтобы напрямую «провести» из Африки в Индию лемуров, слонов, панголинов и т.п. Надо заметить, что сторонники этого направления быстро вошли во вкус и, по едкому замечанию Ч. Дарвина, принялись печь подобные «временные мосты», как блины.
РИСУНОК 63. Пример южнополушарной дизъюнкции: распространение родов пауков-археид по современным материкам (а) и при нанесении его на карту Гондваны (б). Ископаемые рода (из эоценового Балтийского янтаря и из юры Казахстана) помечены крестиком.
Хотя ныне эти реконструкции представляют лишь исторический интерес (они, помимо всего прочего, противоречат концепции изостазии), рациональное зерно в их «констатирующей части» определенно содержалось. Так, ботаник Дж. Гукер (1860) на огромном материале продемонстрировал высокое единство биоты материков Южного полушария (вспомним нотофагусовые леса на юге Австралии, в Новой Зеландии и в Чили). Последователи Гукера предполагали автохтонное (на месте своего нынешнего распространения) развитие биоты Южных континентов – изолированных от Северных и соединенных между собою некими сухопутными связями; предполагалось, что одним из основных источников формирования этой биоты могла служить Антарктида – еще не покрытая тогда ледником и имевшая существенно больший размер, чем ныне. Итак, в рамках этой концепции основное внимание уделялось единству Южной биоты и обеспечивающим ее механизмам расселения.
Иной взгляд на природу южнополушарных дизъюнкций высказывал зоолог А. Уоллес (1876). Он полагал, что основные формообразовательные процессы идут на блоке Северных континентов, откуда более молодые и продвинутые формы животных и растений оттесняют более древних и архаичных все дальше к югу. Южные оконечности материков Южного полушария являются, таким образом, «последней линией обороны» этих реликтовых (сохранившихся от более древних эпох) групп, которые в прежние времена имели гораздо более широкое, обычно панконтинентальное, распространение. С этой точки зрения нет принципиальной разницы между трансокеаническими дизъюнкциями и дизъюнкциями, расположенными на одном материке (например, европейско-дальневосточными, как у жерлянок): и те, и другие возникают в результате сокращения исходного ареала и вымирания «соединительных звеньев». Концепцию Уоллеса (ее позднее назвали теорией оттесненных реликтов) подтверждали ископаемые находки – мнгие «крайне-южные» ныне животные и растения (араукарии, саговники, двоякодышащие рыбы, бескилевые птицы и т.д.) действительно обитали раньше на северных континентах. Пауки-археиды (рисунок 63), например, были первоначально открыты в ископаемом состоянии – в эоценовом балтийском янтаре, – и лишь спустя несколько десятков лет в Южном полушарии были найдены их современные представители. В рамках этой концепции (в отличие от Гукеровской) акцент переносился на архаичность и реликтовый характер Южной биоты.
Модель Уоллеса, хорошо обоснованная палеонтологическими данными и не вводящая таких «избыточных сущностей» (по Оккаму), как сухопутные межматериковые соединения непонятной геологической природы, долго считалась главенствующей. Однако с тех пор, как в геологии возобладала тектоника плит, среди биогеографов возродился интерес к Гукеровской модели: ведь теперь уже не было нужды воздвигать между материками какие-то «сухопутные мосты» – достаточно сдвинуть между собою сами материки. Эта модель позволяет рассматривать разнообразные южнополушарные дизьюнкции как прямой результат раскола Гондваны и сохранения на ее разошедшихся в результате континентального дрейфа фрагментах представителей некогда единой биоты этого мезозойского праматерика (рисунок 63, б). Гукеровские построения считают одним из основных источников наиболее популярной ныне биогеографической концепции – викариантной биогеографии: ее задача состоит в том, чтобы согласовать последовательность ветвления филогенетического древа таксона с рисуемой геофизиками последовательностью расхождения фрагментов Пангеи.