Страница:
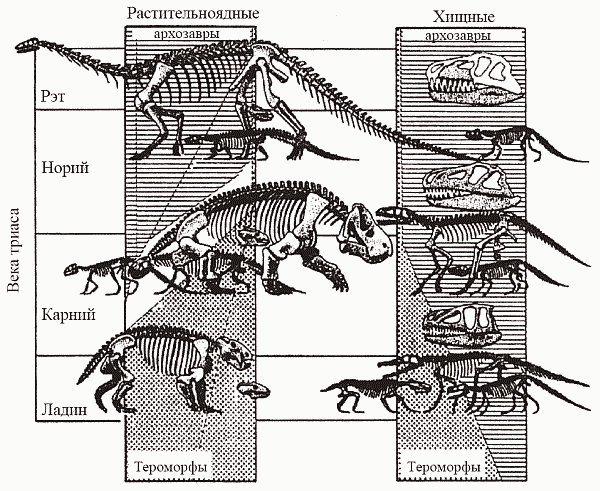
Рис. 41. Замещение тероморфов завроморфами-архозаврами в триасовых фаунах (по Dzik, 1997). Процесс шел параллельно и среди фитофагов (замещение дицинодонтов и ринхозавров на растительноядных динозавров-завропод), и среди хищников (замещение цинодонтов на текодонтов)
А что же другая, завроморфная, ветвь амниот? Они появились в том же позднем карбоне, что и тероморфы, но на протяжении всего палеозоя пребывали на вторых ролях. Правда, растительноядные анапсиды парейазавры (рис. 40, е) стали заметным элементом позднепермских экосистем, но в целом группа занимала явно подчиненное положение... И вот в начале мезозоя завроморфы берут реванш – да какой! В триасе представители тероморфной линии оказываются вытесненными на глубокую перифирию эволюционной сцены, а их место занимают завроморфы-диапсиды (рис. 41); последние к тому же осваивают такие ниши, которые до сих пор амниотам даже не снились – море и воздушное пространство. Триасовый упадок тероморфов, совсем уж было подошедших к тому, чтобы превратиться в маммалий, и внезапный расцвет «настоящих ящеров» – одна из самых интригующих загадок палеонтологии. В.А. Красилов удачно назвал эту ситуацию «мезозойским зигзагом». Упрощенные объяснения типа «иссушения климата» смотрятся тут абсолютно неубедительными – хотя бы потому, что пермь была временем даже более аридным, чем триас...В триасе возникает целый ряд морских групп, эвриапсид: дельфиноподобные ихтиозавры, более похожие на ластоногих плезиозавров (всем хорошо известны эласмозавры с очень длинной шеей и маленькой головой – «змея, продетая сквозь черепаху», но в юре существовали и короткошеие плиозавры с черепом длиной до 3 м), а также менее известные нотозавры и плакодонты, несколько похожие на тюленей (рис. 42, а–г). Их появление в это время вполне логично следует из рептилийной физиологии. Говорят, что змея по вкусу больше всего похожа на цыпленка. Это чистая правда: у завроморфов (к коим принадлежат змеи) большая часть мускулатуры представляет собой «белое мясо», как грудные мышцы курицы. Белая мускулатура бедна (по сравнению с красной) миоглобином и митохондриями, и энергоснабжение мышечных сокращений в ней достигается в основном за счет гликолитического (анаэробного) расщепления глюкозы до молочной кислоты. Для этой реакции не нужен кислород, а скорость ее мало зависит от температуры, что особенно важно для животного, не обладающего гомойотермией. Однако (мы скажем в который раз!) за любое преимущество надо платить. Накапливающаяся в результате гликолиза молочная кислота – метаболический яд, и буквально через 1–2 мин интенсивной работы белая мускулатура практически теряет способность к дальнейшим сокращениям; самопроизвольный распад молочной кислоты (и восстановление работоспособности) требует нескольких часов. Белые мышцы принципиально не годятся для продолжительной работы.
Что касается богатых миоглобином красных мышц, составляющих основу мускулатуры маммалий, то тут ситуация иная. Они содержат огромное количество митохондрий (собственно, они и придают этой ткани красный цвет), и в них молочная кислота – лишь промежуточный продукт, который затем в митохондриях «дожигается» кислородом до углекислого газа и воды с выделением огромного количества энергии. Казалось бы, выгодно во всех отношениях (так что непонятно, зачем вообще кто-то сохраняет белые мышцы) – ан нет: реакции кислородного дыхания требуют повышенной температуры среды. Мало того, что для этого еще нужно выработать гомойотермию, так потом еще почти весь прибыток энергии придется потратить на собственный обогрев! Не зря млекопитающее потребляет (в покое) примерно в 10 раз больше пищи, чем рептилия равного с ним веса – а пищу эту, между прочим, надо еще поймать... В общем, «мамы всякие нужны, мамы всякие важны»: белая мускулатура хорошо (и энергетически дешево) работает «на рывке»[58] , а красная – при длительных постоянных нагрузках; животное с белыми мышцами – хороший «спринтер», а с красными – «стайер».
Вернемся к появлению в триасе разнообразных морских рептилий (кстати, и амфибии-лабиринтодонты тогда же сделали попытку освоить море). Многие исследователи (например, Р. Кэрролл) полагают, что переход к водному образу жизни происходит у низших амниот особенно легко именно в силу низкой интенсивности их метаболизма, способности выдерживать недостаток кислорода (из-за использования анаэробных процессов для работы мышц) и низкой температуры тела (не выше, чем у окружающей воды). Иными словами, рептилии исходно предрасположены к обитанию в водной среде.
Судя по наблюдениям над современными морскими игуанами, передвижение и питание в воде не требует от них структурной или физиологической перестройки; более того, при водной локомоции метаболические затраты этих ящериц составляют лишь четверть от тех, что потребны для перемещения по суше (благо тело в воде ничего не весит – по закону Архимеда).[59] Рептилии без проблем становятся вторичноводными всякий раз, когда жить в воде становится выгоднее с точки зрения обилия пищи и/или отсутствия врагов и конкурентов. Так, в юре возникли морские крокодилы (ноги их превратились в настоящие ласты, и имелся специальный хвостовой плавник, как у ихтиозавра), в раннем мелу – морские черепахи, близкие к нынеживущим, а в позднем мелу – мозазавры, исполинские (до 12 м длиной) морские ящерицы-вараны (рис. 42, д–е).
На суше ситуация складывается принципиально иначе, чем в море: тут белая мускулатура создает для завроморфов серьезные проблемы. Мы уже упоминали о различиях в строении поясов конечностей у тероморфов и завроморфов. Конечности могут быть расположены сбоку от тела (такое их положение называется латеральным) или подведены под него (парасагитально); в первом случае бедренная (или плечевая) кость направлена параллельно земле, а во втором – вертикально (рис. 43, а). Отсюда следуют различия в типе локомоции: рептилии «враскорячку» ползают на брюхе (отсюда происходит русское название группы – «пресмыкающиеся»), тогда как у маммалий тело высоко приподнято над землей, а конечность, двигаясь в плоскости, параллельной оси тела, способна делать широкий шаг (рис. 43, б). Весьма существенно то, что при парасагитальном положении конечностей (маммальный вариант) вес тела принимают на себя (продольно!) кости скелета конечностей, так что мышцам надо лишь не давать этим «подпоркам» отклоняться от вертикального положения; при латеральных же (рептилийных) конечностях тело «подвешено» на самих мышцах, и животное как бы постоянно выполняет упражнение «отжимание от пола» – удовольствие ниже среднего.

Рис. 42. Мезозойские морские рептилии: эвриапсиды (а–г) и диапсиды (д–е) (по Фентон, 1997); а – ихтиозавр; б – плакодонт; в – плезиозавр-эласмозавр; г —плезиозавр-плиозавр; д – мозазавр; е – морской крокодил
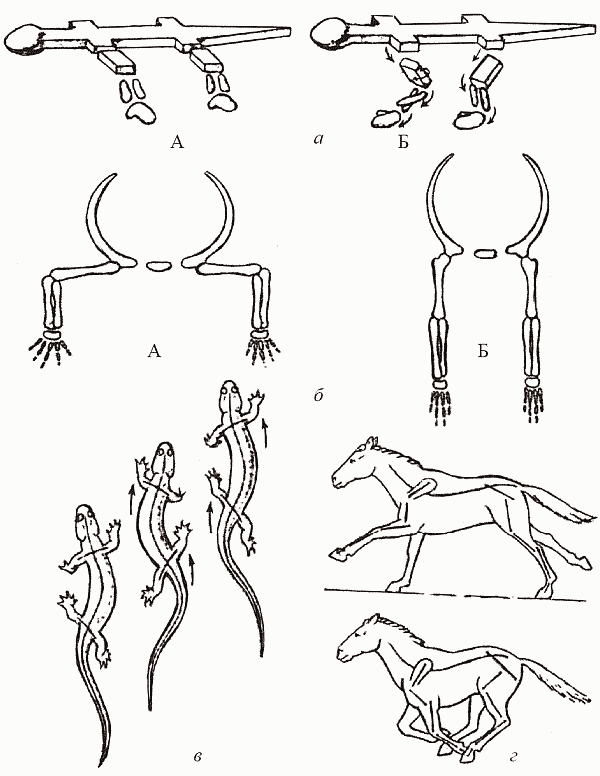
Рис. 43. Локомоция рептилий и маммалий: а – латеральное (А) и парасагитальное (Б) положение конечностей; б – постановка тела у рептилий (А) и маммалий (Б); в – «рептильный» шаг; г – «маммальный» шаг (по Шилову, 1994)
Казалось бы, маммальное строение поясов конечностей безусловно «лучше». В действительности же свои плюсы есть и в рептилийном варианте. Ведь если мускулатура по преимуществу белая, ориентированная «на рывок», то проще бо?льшую часть времени проводить, расслаблено лежа на брюхе, и лишь изредка совершать броски за добычей. При маммальной же постановке конечностей приходится постоянно поддерживать тело на весу; это, во-первых, практически исключает саму возможность использовать белую мускулатуру (которая «устает»), а во-вторых, заставляет тратить много энергии вхолостую (мы помним, что млекопитающее в покое расходует примерно в 10 раз больше энергии, чем рептилия равного размера). Однако когда все плюсы и минусы просуммированы, оказывается, что на суше для завроморфов (с их белой мускулатурой и латеральной конечностью) открыты лишь «профессии» подстерегающего хищника или пассивно защищенного – панцирем или слоем воды – крупного фитофага; остальные экологические ниши оказываются в распоряжении «теплых», активных, высокоподвижных тероморфов.
Ситуация поменялась в триасе, когда на эволюционную арену вышла группа рептилий, являющая собой подлинный венец всей завроморфной линии эволюции – архозавры. К этому подклассу принадлежат крокодилы и вымершие текодонты, динозавры и птерозавры; прямо от него ведут свое происхождение птицы. Из курса зоологии вы помните, что архозавры (о строении которых мы судим в основном по крокодилам) обладают целым рядом уникальных для рептилий продвинутых черт: у них уже полностью разделены правый и левый желудочки сердца, зубы их альвеолярного типа – сидящие в отдельных ячейках (что позволяет аллигаторам иметь «клыки» до 5 см длиной), возникает вторичное нёбо – дополнительная костная перемычка под первичным нёбом, отделяющая дыхательные пути от ротовой полости. Однако для триасовой победы архозавров над тероморфами наиболее существенным оказалось, по всей видимости, именно видоизменение конечностей и их поясов (в этом смысле крокодилы как раз не показательны – их конечности очень примитивны, близки к предковому для архозавров типу), которое весьма неожиданным образом разрешило проблему скоростной локомоции.
Дело в том, что по ряду анатомических причин (они были детально обсуждены еще А. Ромером в 1922 году) перевести в парасагитальное положение задние конечности, которые причленены к позвоночнику, намного проще, чем передние, соединенные с грудиной. Достаточно сказать, что тероморфы сумели «подвести под тело» задние ноги еще в перми (у горгонопсид), а вот полная парасагитальность выработалась в этой линии много позже – лишь у триасовых цинодонтов. Итоговая же скорость передвижения «доцинодонтного» тероморфа, как легко догадаться, все равно лимитируется именно «медленными» – латеральными – передними конечностями.
Архозавры появились в поздней перми, почти одновременно с терапсидами, и одновременно же с ними выработали парасагитальность задних конечностей. А вот дальше архозавры, вместо того чтобы возиться с преобразованием пояса передних конечностей, вообще отказались от их использования для движения и выработали принципиально новый тип локомоции – бипедальный (двуногое хождение). О потенциальных скоростных возможностях такого двуногого диапсида дают некоторое представление современные страусы... Бипедальность тоже требует ряда серьезных анатомических перестроек (прежде всего создания опорного таза с консолидированными позвонками крестцового отдела), однако сделать это, как ни странно, оказалось легче, чем решить простенькую на первый взгляд задачу снятия ограничений в подвижности плечевого сустава – на что тероморфам пришлось потратить почти 100 млн лет.[60]
Так в триасе возникла жизненная форма высокоскоростного бипедального существа. Именно «двуногость» открыла динозаврам путь к 130-миллионолетнему владычеству на суше (есть много четвероногих растительноядных динозавров, но все они произошли от двуногих предков, «опустившихся на четвереньки»). Среди наземных хищников в крупном размерном классе эта жизненная форма стала вообще единственной и, однажды сформировавшись, практически не менялась на протяжении всего мезозоя (главный персонаж «Парка юрского периода» – позднемеловой динозавр Tyrannosaurus rex – внешне мало чем отличается от крупного триасового текодонта Ornithosuchus). Более того, впоследствии именно бипедальная локомоция позволила двум линиям архозавров – птерозаврам и птицам – независимо преобразовать переднюю конечность в машущее крыло и освоить активный полет. Впрочем, «это уже совсем другая история»...
Глава 11
Поздний мезозой: эволюция наземных позвоночных (2). Завроморфный мир. Маммализация териодонтов. Динозавры и их вымирание.
Итак, мы знаем, что с самого момента появления истинно наземных позвоночных – амниот – произошло их разделение на две эволюционные ветви: тероморфов и завроморфов. Обе группы возникли одновременно в позднем карбоне, но первоначальный успех оказался на стороне тероморфов (с их неороговевшими покровами, благоприятствующими активной терморегуляции, относительно большим мозгом и высоким уровнем метаболизма); они доминировали в позднем карбоне и перми. Однако в триасе высшие завроморфы (архозавры) одержали верх над высшими тероморфами (терапсидами), начисто вытеснив их из крупного размерного класса. «Изобретенная» архозаврами жизненная форма бипедального хищника оказалась столь совершенной, что на протяжении всего мезозоя наземных хищников иного типа не возникало вовсе, а крупный размерный класс оказался полностью закрыт и для тероморфов, и для прочих (не-архозавровых) завроморфов. Судя по всему, мы тут опять имеем дело со случаем «абсолютного оружия» (это уже знакомо нам по ситуациям с вендским зоопланктоном и карбоновыми стрекозами). Терапсидам удалось-таки «выставить на конкурс» свою модель высокоподвижного хищника (с двумя парами парасагитальных конечностей, с красной мускулатурой и частичной теплокровностью), однако они поспели к шапочному разбору, когда экологические ниши были уже поделены.


Очень интересно сопоставить структуру сообщества наземных, вторично-водных и летающих позвоночных. Если на суше господствовали амниоты с самым активным метаболизмом – тероморфы и архозавры (последние имели четырехкамерное сердце и, как мы увидим дальше, по-своему решили проблему гомойотермии), то водная среда оказалась вотчиной рептилий с низким уровнем обмена и белой мускулатурой, ориентированной на бескислородные процессы. Для архозавров она была малоподходящей (водный образ жизни ведут лишь самые архаичные их представители, фитозавры и крокодилы), зато здесь процвели низшие диапсиды (эвриапсиды) и анапсиды: ихтиозавры, плезиозавры, плакодонты, морские черепахи. Нет такой группы диапсид, которая не «делегировала» бы своих представителей в воду – были даже водные клювоголовые, плеврозавры.
Все водные рептилии – хищники (среди них не было не только фитофагов, но даже и планктоноядов вроде усатых китов или китовой акулы), и все крупные – заметно крупнее своих сухопутных предков (если таковые известны). Весьма показательны в этом последнем отношении «ящерицы» (в широком смысле), на суше практически никогда не выходившие из малого размерного класса, но породившие гигантских морских мозазавров и пресноводных хампсозавров (жизненная форма «крокодил»). Начиная с триаса водные рептилии оккупировали верхние трофические этажи морских экосистем, полностью вытеснив оттуда и рыб, и головоногих – эти предыдущие «владыки морей» служили им основной пищей. Ситуация изменилась лишь в начале мела, когда появились акулы современного типа; возможно, именно они конкурентно вытеснили самых «рыбообразных» из рептилий – ихтиозавров.
С освоением воздуха картина, как и следовало ожидать, была обратная (по сравнению с морем): здесь преуспели именно высшие архозавры, имевшие самый активный метаболизм, – птицы и птерозавры, которые тоже почти наверняка были теплокровными. Так, мелкий птерозавр Sordes pilosus (что в переводе с латыни означает «нечисть волосатая») был покрыт чем-то вроде волосяного покрова не вполне ясной природы; трудно представить себе, зачем он мог понадобиться, если не для теплоизоляции. В пользу теплокровности птерозавров может косвенно свидетельствовать и строение их костей, полых и несущих перфорации – как у птиц, у которых кости пронизаны воздушными мешками (обеспечивающими бесперебойность дыхания во время машущего полета). Если дыхательная система в этих двух группах построена аналогичным образом, то и интенсивность обмена у птерозавров должна быть очень высока.

У динозавров тело было приподнято над землей гораздо больше, чем у любых современных рептилий с их широко расставлеными в стороны конечностями. Об этом свидетельствуют, в числе прочего, их узкие следовые дорожки – как у маммалий и птиц. Иногда указывают на то, что динозавры просто не могли нести многотонное тело на своих не до конца выпрямленных («раскоряченных» по-рептильному) конечностях – это невозможно: надо, чтобы вес принимали на себя не мышцы, а вертикально поставленные кости (см. выше). Верно. Но все-таки это, пожалуй, не причина, а следствие. Ведь «прямохождение» выработалось уже у самых первых динозавров – небольших существ с комплекцией кенгуру, перед которыми проблема избыточного веса еще не стояла. Скорее наоборот: само происхождение динозавров было связано с общей активизацией обмена веществ. Это открыло им путь к парасагитальности (для нее, как мы помним, необходима продолжительная мышечная активность; следовательно – красная мускулатура, следовательно – аэробный метаболизм и теплокровность, и т.д.), а уже парасагитальность, в свою очередь, позволила наращивать размеры тела аж до завроподных 30 тонн.
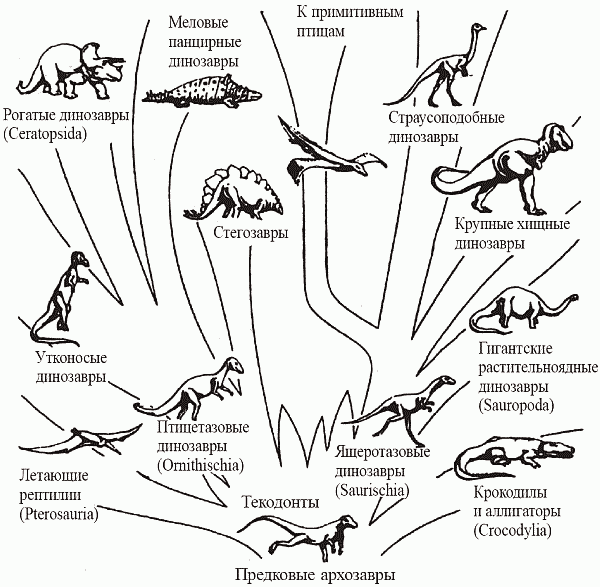
Рис. 44. Филогения архозавров (по Romer, 1950)
Эволюция архозавров (рис. 44) выглядит так. От пермо-триасовых текодонтов обособились к концу триаса все остальные отряды: крокодилы, птерозавры и два отряда динозавров – ящеротазовые (Saurischia) и птицетазовые (Ornitischia). К ящеротазовым относятся такие на первый взгляд несхожие друг с другом формы, как хищные двуногие тероподы (размером от тиранозавра – 6 м высотой, самый крупный наземный хищник всех времен – до прекурзора, очаровательного создания величиной с ворону) и четвероногие растительноядные завроподы – самые большие наземные животные (диплодок достигал 26 м в длину при весе 30 т).[61] К птицетазовым принадлежат исключительно растительноядные формы, по большей части крупные и очень крупные: двуногие гадрозавры (утконосые динозавры) и игуанодоны, а также вторично-четвероногие анкилозавры (панцирные динозавры), стегозавры и цератопсиды (рогатые динозавры) (рис. 45).Рис. 45. Наземные архозавры – динозавры и текодонты (по Фентон, 1997); а —диплодок; б – его череп; в – тиранозавр; г – мелкая теропода Ornitholestes; д – гадрозавр; е – трицератопс; ж – стегозавр; з – анкилозавр; и – текодонт
От мелких хищных теропод позднее произошли птицы. Следует отметить, что текодонты еще в триасе «изобрели» практически все жизненные формы, которыми архозавры пользовались потом на протяжении всего мезозоя: хищные бипедальные орнитозухии принципиально не отличались от теропод, растительноядные этозавры с мощным панцирем весьма походили на четвероногих пти-цетазовых, а полуводные фитозавры – это просто крокодил-крокодилом. Подчеркнем: указанные пары (например, фитозавр – крокодил) не связаны сколь-нибудь тесным родством, речь идет именно о реализации определенных экологических стратегий (рис. 46).Рис. 46. Триасовые текодонты – экологические аналоги более поздних архозавров (динозавров и крокодилов): а – этозавр Steganolepis (аналог анкилозавров); б – псевдозух Ornithosuchus (аналог теропод-карнозавров); в – фитозавр Parasuchus(аналог крокодилов) (по Кэрроллу, 1992)
Когда говорят о структуре мезозойского сообщества наземных позвоночных, сразу замечают, что крупный размерный класс (Э. Олсон назвал его «доминантным сообществом») в нем был полностью сформирован архозаврами: и фитофаги, и хищники в нем представлены сперва текодонтами, потом динозаврами. Реже обращают внимание на другое обстоятельство: малый размерный класс («субдоминантное сообщество») оказался для архозавров почти закрытым – ровно в той же степени, как крупный – для тероморфов. Среди малоразмерных существ (менее 1 м) доминировали териодонты (и их прямые потомки – млекопитающие), а на вторых ролях выступали низшие диапсиды – ящерицы и клювоголовые (ныне от этой группы уцелела лишь гаттерия); питались они насекомыми и, реже, друг другом (фитофагии в малом размерном классе не было вовсе). Итак, высшие завроморфы (динозавры) и высшие тероморфы (маммалии) с самого начала сформировали «параллельные миры», практически не взаимодействовавшие между собой.Очень интересно сопоставить структуру сообщества наземных, вторично-водных и летающих позвоночных. Если на суше господствовали амниоты с самым активным метаболизмом – тероморфы и архозавры (последние имели четырехкамерное сердце и, как мы увидим дальше, по-своему решили проблему гомойотермии), то водная среда оказалась вотчиной рептилий с низким уровнем обмена и белой мускулатурой, ориентированной на бескислородные процессы. Для архозавров она была малоподходящей (водный образ жизни ведут лишь самые архаичные их представители, фитозавры и крокодилы), зато здесь процвели низшие диапсиды (эвриапсиды) и анапсиды: ихтиозавры, плезиозавры, плакодонты, морские черепахи. Нет такой группы диапсид, которая не «делегировала» бы своих представителей в воду – были даже водные клювоголовые, плеврозавры.
Все водные рептилии – хищники (среди них не было не только фитофагов, но даже и планктоноядов вроде усатых китов или китовой акулы), и все крупные – заметно крупнее своих сухопутных предков (если таковые известны). Весьма показательны в этом последнем отношении «ящерицы» (в широком смысле), на суше практически никогда не выходившие из малого размерного класса, но породившие гигантских морских мозазавров и пресноводных хампсозавров (жизненная форма «крокодил»). Начиная с триаса водные рептилии оккупировали верхние трофические этажи морских экосистем, полностью вытеснив оттуда и рыб, и головоногих – эти предыдущие «владыки морей» служили им основной пищей. Ситуация изменилась лишь в начале мела, когда появились акулы современного типа; возможно, именно они конкурентно вытеснили самых «рыбообразных» из рептилий – ихтиозавров.
С освоением воздуха картина, как и следовало ожидать, была обратная (по сравнению с морем): здесь преуспели именно высшие архозавры, имевшие самый активный метаболизм, – птицы и птерозавры, которые тоже почти наверняка были теплокровными. Так, мелкий птерозавр Sordes pilosus (что в переводе с латыни означает «нечисть волосатая») был покрыт чем-то вроде волосяного покрова не вполне ясной природы; трудно представить себе, зачем он мог понадобиться, если не для теплоизоляции. В пользу теплокровности птерозавров может косвенно свидетельствовать и строение их костей, полых и несущих перфорации – как у птиц, у которых кости пронизаны воздушными мешками (обеспечивающими бесперебойность дыхания во время машущего полета). Если дыхательная система в этих двух группах построена аналогичным образом, то и интенсивность обмена у птерозавров должна быть очень высока.
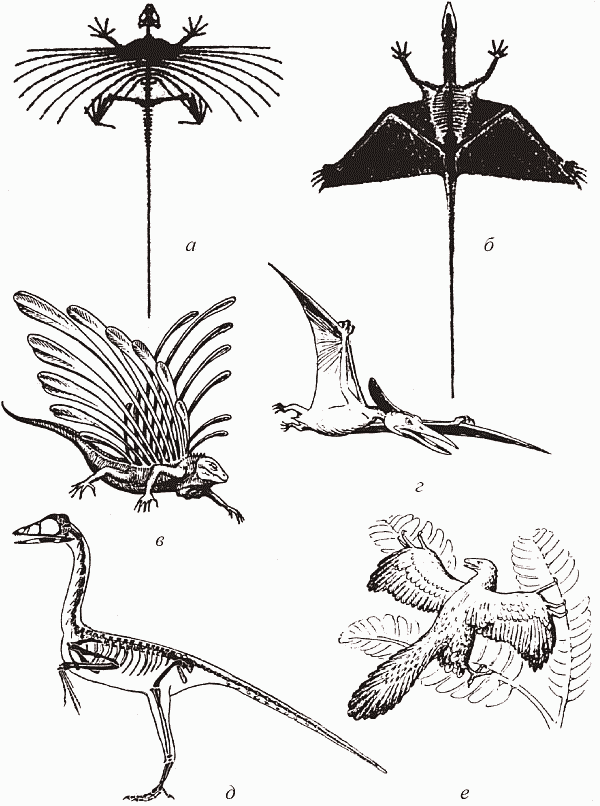
Рис. 47. Завоевание воздуха рептилиями: а, б, в – планеры из числа триасовых низших диапсид; г – птерозавры; д, е – предки птиц (по Dzik, 1997, и Фентон, 1997): а – Icarosaurus siefkei; б – Sharovipteryx mirabilis; в – Longisquama insignis; г – птеранодон; д – протоавис; е – археоптерикс
Самое интересное, что в триасе (и даже в конце перми) низшие диапсиды («ящерицы») совершили целый ряд попыток завоевания воздуха, некоторые из которых кажутся крайне экзотичными (рис. 47, а–в), однако это были варианты планирующего полета: на активный, машуший, полет у этих рептилий явно не хватает энергетики. Появившиеся в конце триаса воздушные архозавры (птерозавры) разом пресекли эти неуклюжие потуги низших диапсид. Первыми птерозаврами были хвостатые рамфоринхи, размером от воробья до вороны; в дальнейшем птерозавры становились все более крупными (например, птеродактили были не меньше орла, а у их верхнемеловых представителей, птеранодонов, размах крыльев достигал 8 м – это самые большие летающие существа всех времен[62] ) и наверняка перешли к парящему полету, вроде современных кондоров. Исчезновение в конце юры мелких высокоманевренных рамфоринхов (хвост летающему существу нужен как противовес для разворотов с малым радиусом) с машущим полетом, после чего остались лишь крупноразмерные «живые планеры» – птеродактили, судя по всему, было напрямую связано с появлением в это время птиц. В воздушном сообществе между птерозаврами и птицами происходит такое же размежевание по размерным классам, как и в наземном – между динозаврами и маммалиями. Рамфоринхи (судя по содержимому их желудков) были частью насекомоядными, частью рыбоядными – что-то вроде современных зимородков (рис. 48). В дальнейшем рыбоядность стала для птерозавров «генеральной линией» – недаром почти все их скелеты найдены в мелководных морских отложениях.[63]Рис. 48. Охотящиеся рамфоринхи (по Norman, 1994)
Итак, в море господствуют крупные рептилии из числа низших завроморфов с «холодным» метаболизмом, в воздухе – относительно мелкие и теплокровные высшие архозавры. А что же «владыки суши» – динозавры? Вопрос об их возможной теплокровности дискутировался давно. В последнее время большинство палеонтологов сходятся на том, что интенсивность обмена у динозавров может быть и не достигала птичьей или маммальной, но заметно превышала таковую «обычных» рептилий. Во всяком случае, ранние представления о динозаврах как о массивных малоподвижных существах отошли в прошлое. Ужасные тероподы-велоцирапторы из «Парка юрского периода», напоминающие своим поведением крупных нелетающих птиц, судя по всему, вполне соответствуют реальности.У динозавров тело было приподнято над землей гораздо больше, чем у любых современных рептилий с их широко расставлеными в стороны конечностями. Об этом свидетельствуют, в числе прочего, их узкие следовые дорожки – как у маммалий и птиц. Иногда указывают на то, что динозавры просто не могли нести многотонное тело на своих не до конца выпрямленных («раскоряченных» по-рептильному) конечностях – это невозможно: надо, чтобы вес принимали на себя не мышцы, а вертикально поставленные кости (см. выше). Верно. Но все-таки это, пожалуй, не причина, а следствие. Ведь «прямохождение» выработалось уже у самых первых динозавров – небольших существ с комплекцией кенгуру, перед которыми проблема избыточного веса еще не стояла. Скорее наоборот: само происхождение динозавров было связано с общей активизацией обмена веществ. Это открыло им путь к парасагитальности (для нее, как мы помним, необходима продолжительная мышечная активность; следовательно – красная мускулатура, следовательно – аэробный метаболизм и теплокровность, и т.д.), а уже парасагитальность, в свою очередь, позволила наращивать размеры тела аж до завроподных 30 тонн.