Страница:
Как известно, осадочные породы можно разделить по их происхождению на биогенные, т.е. образующиеся при непосредственном участии живых организмов (например, писчий мел), и абиогенные (например, песчаник). В 1922 году В. И. Вернадский, исходя из структуры и геохимических особенностей различных осадочных пород, высказал такое суждение: в геологической истории Земли не удается обнаружить периода (сколь угодно древнего), когда образование всех известных для него осадков происходило бы заведомо абиогенным путем. Следовательно, заключал он, если мы будем твердо стоять на почве эмпирических обобщений (а не всякого рода «общих соображений»), то нам придется признать, что жизнь существовала на Земле всегда – «биосфера геологически вечна». Этот несколько ошарашивающий вывод можно строго переформулировать в виде презумпции (см. дополнение к главе 1): «следует считать, что жизнь на Земле существовала изначально – до тех пор, пока не доказано обратное» (до того презумпция была противоположной). Так вот, в последние десятилетия в области изучения древнейших форм жизни произошла подлинная революция, в ходе которой точка зрения Вернадского получила весьма весомые фактические подтверждения.
Как мы помним из главы 1, древнейшие из известных минералов имеют возраст 4,2 млрд лет (оценка возраста Земли в 4,5–4,6 млрд лет основана на анализе вещества метеоритов и лунного грунта). Возраст же древнейших пород, в которых найден углерод заведомо органического происхождения (в углероде, принимавшем когда-либо участие в реакциях фотосинтеза, необратимо меняется соотношение изотопов 12C и 13C) составляет... 3,8 млрд лет. Цифра, согласитесь, неслабая и сама по себе, однако тут есть еще важное дополнительное обстоятельство. Дело в том, что формацию Исуа в Гренландии, где были обнаружены эти углеродистые прослои, составляют вообще древнейшие на Земле осадочные породы. Значит, первые достоверные следы жизни появляются на Земле одновременно с первыми достоверными следами воды. А поскольку ископаемые могут сохраняться только в осадочных породах (за редчайшими исключениями, вроде захоронений под вулканическими пеплопадами и т.п.), то можно сформулировать и так: достоверные следы жизни известны в геологической летописи Земли с того самого момента, когда возникает принципиальная возможность их фиксации. Таким образом, «презумпция Вернадского» стала теперь фактически неуязвимой.
Еще недавно биологи уверено рисовали (исходя из общих соображений) такую картину. Самый длинный отрезок в истории Земли приходился на образование первых биологических систем из неорганической материи. Несколько меньшее время потребовалось на возникновение первых клеток, и лишь после этого начался все ускоряющийся процесс собственно биологической эволюции. Первыми живыми существами были гетеротрофные микроорганизмы (гетеротрофы), питавшиеся «первичным бульоном» – той органикой, что в избытке возникала в первичной атмосфере и океане в результате процессов, частично смоделированных Миллером. Затем уже возникли и автотрофы, синтезирующие органику из углекислого газа и воды, используя для этого энергию окислительных химических реакций (хемоавтотрофы) или солнечного света (фотоавтотрофы).
Однако реальные факты вынудили отказаться от этой умозрительной схемы. В числе прочего не нашли подтверждений и представления об исходной гетеротрофности живых существ; судя по всему, автотрофность и гетеротрофность возникли одновременно. Один из ведущих специалистов по микробным сообществам Г. А. Заварзин обращает внимание на то, что «первичный бульон» даже теоретически не мог быть источником пищи для «первичной жизни», т. к. является конечным, исчерпываемым ресурсом. Поскольку живые существа размножаются в геометрической прогрессии (вспомните всякого рода занимательные задачки о потомках одной бактерии, заполняющих сперва наперсток, потом железнодорожный состав и т.д.), потомство первых же гетеротрофов должно было бы сожрать весь этот «бульон» за совершенно ничтожное время; после этого все они, естественно, мрут от голода – и извольте создавать жизнь на Земле по новой. Не меньшие неприятности, впрочем, ожидают и чисто автотрофную биосферу, которая в том же темпе (и с тем же конечным результатом!) свяжет весь углерод на планете в виде неразложимых высокомолекулярных соединений.
Первичность хемоавтотрофности (относительно фотоавтотрофности) тоже принято было считать несомненной; серьезным аргументом тут является то обстоятельство, что архебактерии (наиболее архаичные из всех прокариот) – именно автотрофы. Но и здесь, как выяснилось, все не так уж очевидно. Молекула фотосинтезирующего пигмента (например, хлорофилла) поглощает квант света; в дальнейшем энергия этого кванта используется в различных химических превращениях. Так вот, есть серьезные основания полагать, что первичной функцией этих пигментов была просто-напросто нейтрализация разрушительной для организма (да и вообще для любой высокомолекулярной системы) энергии квантов ультрафиолетового излучения, беспрепятственно проникавшего в те времена сквозь лишенную озонового слоя атмосферу. Впоследствии те, кто приспособился еще и использовать эту энергию «в мирных целях», разумеется, получили гиганские преимущества, однако сама по себе «радиационная защита» ДНК при помощи пигментов должна была сформироваться еще на стадии доорганизменных гиперциклов. Понятно, что доводить эту защиту до совершенства имело смысл лишь тем из них, кто обитал в поверхностном слое океана (10-метровый слой воды полностью защищает от ультрафиолета любой интенсивности); именно они, судя по всему, и дали начало фотоавтотрофам, тогда как глубины остались в распоряжении хемоавтотрофов.
Есть и прямые доказательства существования фотоавтотрофов с самых давних времен (древнейшие организмы из формации Исуа, как мы помним, были именно фотосинтезирующими, а в породах возраста 3,1 млрд лет содержатся остатки хлорофилла – фитан и пристан – и даже неразложившийся цианобактериальный пигмент фикобилин), но гораздо проще отслеживать деятельность фотоавтотрофов по одному из ее побочных продуктов – кислороду. Впервые такой анализ был проведен Л. Беркнером и Л. Маршаллом (1962); они пришли к выводу, что количество кислорода на Земле росло на протяжении всей ее истории – постоянно, но крайне медленно, и вплоть до начала кембрия атмосфера оставалась практически бескислородной (не более 1% от его нынешнего количества). Однако данные, накопленные к настоящему моменту, заставляют предполагать, что насыщение атмосферы кислородом произошло гораздо раньше, в более отдаленные времена.
В отличие от других атмосферных газов кислород не может быть получен путем дегазации магмы, поэтому он отсутствовал в первичной атмосфере Земли, которая была восстановительной. Об этом свидетельствует наличие в древнейших, главным образом катархейских, породах легко окисляемых, но не окисленных (несмотря на пребывание на земной поверхности) веществ – таких, как графит (углерод), лазурит и т.д. Небольшие количества кислорода, образовывавшегося в результате фотодиссоциации молекул воды под действием жесткого ультрафиолетового излучения, полностью расходовались на окисление постоянно выделяемых вулканами аммиака, закиси углерода, метана и сероводорода. Однако с определенного момента в океанах начали образовываться так называемые полосчатые железные руды (джеспиллиты), представляющие собой чередование бедных и обогащенных окислами железа прослоев. Они формируются, когда двухвалентное (закисное) железо, выработавшееся в результате вулканической активности и растворенное в морской воде, реагирует в поверхностных слоях океана с молекулярным кислородом, окисляется до трехвалентного (окисного) и выпадает в виде нерастворимого осадка на дно. Таким образом, начало образования руд этого типа (состоящих в основном из гематита Fe2O3 и магнетита FeO · Fe2O3) означает появление на Земле достаточно мощного источника молекулярного кислорода, каковым могут быть только фотосинтезирующие организмы. Впервые джеспиллиты появляются все в той же формации Исуа; это может служить независимым подтверждением того, что содержащееся в ней углеродистое вещество действительно принадлежало фотосинтетикам (рис. 16).
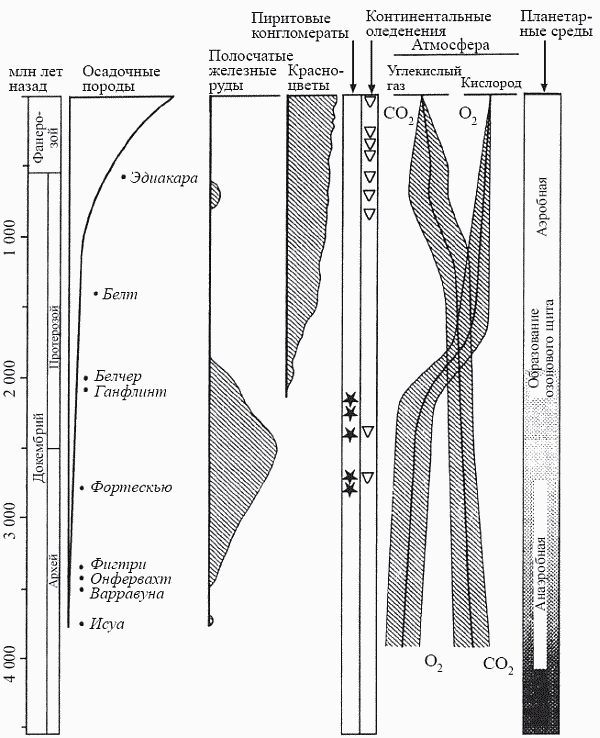
Ситуация радикально изменилась около 2 млрд лет назад, когда процесс гравитационной дифференциации недр (см. главу 3) привел к тому, что бо?льшая часть железа перешла в ядро планеты и растворенное в морской воде закисное железо не могло уже возобновляться по мере выбывания из цикла. Фотоавтотрофы получили возможность завершить свою титаническую работу по окислению закисного железа и осаждению его в виде джеспиллитов; именно в это время возникли все крупнейшие месторождения железа, такие, как Курская магнитная аномалия. В дальнейшем руды этого типа уже не образовывались (за исключением очень краткого эпизода в самом конце докембрия – о нем речь пойдет в главе 6). Кислород, ранее полностью расходовавшийся на этот процесс, стал теперь мало-помалу насыщать атмосферу; с этого времени пиритовые конгломераты исчезли, а на смену донно-морским железным рудам пришли терригенные (имеющие наземное происхождение) красноцветы, т.е. процесс окисления железа начался и на суше (рис. 16). Яркий образ для описания этих событий придумал Г. А. Заварзин: он назвал их временем, когда «биосфера „вывернулась наизнанку“ за счет появления кислородной атмосферы как продукта обмена цианобактерий, и вместо кислородных «карманов» появились анаэробные «карманы» в местах разложения органики».
Именно в это время (1,9 млрд лет назад) в канадской формации Ганфлинт впервые появляются звездчатые образования, полностью идентичные тем, что образует ныне облигатно-аэробная марганцевоосаждающая бактерия Metallogenium. Без кислорода окисление железа и марганца не идет, и образуемые этой бактерией металлические кристаллы в виде характерных «паучков» возникают только в сильно окислительной обстановке. Это должно означать, что в тот момент содержание кислорода в атмосфере уже достигло величины как минимум в 1% от современного (точка Пастера). Именно с этой пороговой концентрации становится «экономически оправданным» налаживание процесса кислородного дыхания, в ходе которого из каждой молекулы глюкозы можно будет получать 38 энергетических единиц (молекул АТФ) вместо двух, образующихся при бескислородном брожении. С другой стороны, в атмосфере начинает возникать озоновый слой, преграждающий путь смертоносному ультрафиолету, что ведет к колоссальному расширению спектра пригодных для жизни местообитаний.
Одним из симптомов произошедшей «кислородной революции» стало то, что у автотрофов возникли серьезные проблемы с азот-фиксацией, т.е. переводом атмосферного азота в усвояемую, ионную форму. Азотфиксация возможна только в бескислородных условиях или при низком содержании кислорода, поэтому у автотрофов (таких, как цианобактерии), выделяющих кислород в процессе собственной жизнедеятельности, азотфиксация идет в специальных клетках с «герметической» утолщенной оболочкой – гетероцистах и акинетах. Акинеты впервые появляются в палеонтологической летописи как раз после «кислородной революции» – около 1,4 млрд лет назад.
Примерно к середине протерозоя (1,7–1,8 млрд лет назад) «кислородная революция» в целом завершается и мир становится аэробным (рис. 16). Впрочем, с точки зрения существ, составлявших тогдашнюю биосферу, этот процесс следовало бы назвать иначе: необратимое отравление кислородом атмосферы планеты. Смена анаэробных условий на аэробные не могла не вызвать катастрофических перемен в структуре тогдашних экосистем. В действительности «кислородная революция» есть не что иное, как первый в истории Земли глобальный экологический кризис.
Во всех предыдущих построениях мы имели дело лишь с геохимическими следствиями тех процессов, что происходили на Земле при предполагаемом нами участии живых существ. Но располагаем ли мы хоть какой-то информацией о самих этих существах? Можно ли их «пощупать пальцами»? Оказывается, можно. В последние десятилетия были разработаны специальные методы обработки осадочных горных пород, позволяющие выделять содержащиеся в них клеточные оболочки, а в некоторых случаях даже получать косвенную информацию о внутреннем строении этих клеток. К настоящему моменту в докембрийских породах обнаружено множество одноклеточных организмов, древнейшие из них – в местонахождениях Варравуна в Австралии (3,5 млрд лет) и Онфервахт в Южной Африке (3,4 млрд лет). Это оказались несколько видов цианобактерий (сине-зеленых водорослей), ничем особенно не отличающихся от современных; мы уже упоминали о том, что из пород возраста 3,1 млрд лет был выделен пигмент фикобилин, который используется при фотосинтезе современными цианобактериями (никем, кроме них).
Итак, в раннем докембрии существовал совершенно особый мир, формируемый прокариотными организмами – бактериями и циано-бактериями. Разделение живых существ на прокариоты и эукариоты (эти термины были введены в 1925 году Э. Шаттоном), основанное на наличии или отсутствии в их клетках оформленного ядра, теперь считают существенно более фундаментальным, чем, например, разделение на «животные» и «растения». Из многих их различий для нас сейчас важнее всего то, что среди прокариот неизвестны многоклеточные организмы. Есть нитчатые и пальмеллоидные[12] формы цианобактерий, однако уровень интеграции клеток в этих структурах – это все-таки уровень колонии, а не организма. А поскольку времени на всякого рода эксперименты у прокариот, как мы видели, было более чем достаточно (3,5 млрд лет), то приходится предположить, что по каким-то причинам на прокариотной основе многоклеточность не возникает в принципе. Может быть, дело в отсутствии у них центриолей и митотического веретена, без которого невозможно точно ориентировать в пространстве делящиеся клетки и формировать из них сколь-нибудь сложные ансамбли. Может быть – в свойствах их клеточной оболочки, препятствующих межклеточным взаимодействиям; это делает невозможным обмен веществами, так что клетки, находящиеся внутри организма и не имеющие прямого контакта с окружающей средой (что неизбежно при многоклеточности), не могли бы питаться и выводить продукты распада. Может быть – в чем-то еще. Как бы то ни было, такой путь повышения уровня организации, как многоклеточность, для прокариот оказался закрытым. Выход из этого тупика они нашли весьма оригинальный.
Для докембрийских осадочных толщ чрезвычайно характерны строматолиты (с греческого – «каменный ковер») – тонкослойчатые колонны или холмики, состоящие главным образом из карбоната кальция (рис. 17, а и б). Эти структуры известны еще с середины XIX века, однако их происхождение оставалось совершенно неясным до 30-х годов XX века, когда было сделано замечательнейшее открытие в области классической (т.е. немолекулярной) биологии. На литорали залива Шарк-Бэй в западной Австралии и на атлантическом побережье Багамских островов были найдены небольшие рифовые постройки неизвестного ранее типа – плосковершинные известковые «бочки» диаметром 1–1,5 м (рис. 17, в) и располагающиеся ниже уровня отлива слоистые корки; все это при внимательном рассмотрении оказалось... современными строматолитами. Выяснилось, что строматолит образуется в результате жизнедеятельности совершенно ни на что не похожего прокариотного сообщества, называемого цианобактериальным матом. Маты существуют во многих районах мира, главным образом в таких пересоленных лагунах, как Сиваш, однако настоящие строматолиты (как в Шарк-Бэй и на Багамской отмели) они образуют довольно редко.
1. Плотный верхний слой – поверхность роста (1–1,5 мм), в котором есть и автотрофы (продуцирующие кислород цианобактерии-фотосинтетики), и гетеротрофы (облигатно-аэробные, т.е. неспособные жить в отсутствие кислорода, бактерии).
2. Тонкая (менее 1 мм) подкладка; в ней автотрофами служат бактерии, осуществляющие бескислородный фотосинтез (при котором источником водорода является не вода, а другие вещества, например, сероводород – тогда выделяется не молекулярный кислород, а сера). Гетеротрофами же служат аэробы (факультативные), которые могут использовать кислород, но в его отсутствие способны удовлетворять свои потребности в энергии не дыханием, а брожением.
3. Ниже двух верхних слоев, в которых идут процессы фотосинтеза и которые в совокупности называют фотической зоной мата, лежит толстая бескислородная зона, где процветают разнообразные анаэробы: для них молекулярный кислород не нужен или даже является ядом.
На поверхность обитающего на мелководье мата постоянно выпадают частички осадка (обычно кристаллы карбоната кальция), которые затемняют соответствующе участки фотической зоны и должны постепенно ухудшать условия идущего в ней фотосинтеза – вплоть до неизбежной гибели фотосинтетиков. От катастрофы эти микроорганизмы спасает положительный фототаксис: по мере того, как интенсивность света в фотической зоне падает, они мигрируют вверх сквозь накапливающийся слой осадка, образуя выше него новую поверхность роста с подкладкой. Миграция эта идет разными способами: нитчатые формы обладают способностью к скользящему движению сквозь осадок, пальмеллоидные – к ускоренному нарастанию верхних частей колонии; еще один путь – временный переход в латентное состояние с последующей реколонизацией поверхности осадка.
Оказавшиеся таким образом внутри бескислородной зоны мата осадки структурируются и слой за слоем наращивают верхнюю поверхность строматолита; измерения показали, что современные строматолиты прирастают со скоростью около 0,3 мм/год. Каждый слой состоит из пары слойков – светлого и темного, состоящих, соответственно, из более мелких и более крупных кристаллов карбоната кальция; крупные кристаллы образуются в тот сезон года, когда процесс осаждения идет быстрее (рис. 17, д). Следует подчеркнуть, что вещество, из которого построен стоматолит, не создается матом – последний лишь структурирует естественное осадконакопление. Строматолиты чаще всего состоят из карбоната кальция лишь потому, что карбонатный тип осадконакопления в море наиболее обычен, однако в иных гидрохимических условиях формируются строматолиты фосфатные, кремнезёмные, железистые и др.[13]
Не все маты представляли собой донные сообщества. Помимо уже известных нам строматолитов, следы жизнедеятельности микробных сообществ представлены еще и онколитами – более или менее сферическими образованиями, в которых известковые слои располагаются не линейно (как в строматолите); они имеют концентрическую структуру. Предполагается, что онколиты создавались особыми прокариотными сообществами, похожими на плавающие в толще воды шары, оболочка которых являлась матом; осадки отлагались кнутри от оболочки этого «батискафа», а плавучесть ему придавали образуемые при метаболизме газы, не могущие покинуть замкнутую полость внутри колонии. Позже мы столкнемся с особым типом водорослевого сообщества – плавающим матом, характерным для некоторых внутренних водоемов мезозоя (см. главу 9).
Итак, мат – это высокоинтегрированное сообщество с чрезвычайно сложно организованной трофической структурой: два верхних слоя составлены продуцентами и консументами первого порядка («растениями» и «травоядными»), соединенными в пищевую цепь пастбищного типа, а нижний слой сформирован редуцентами («падальщиками»), получающими органику из верхних слоев по детритной цепи (рис. 17). Более того, измерения показали, что мат является вообще одной из самых сбалансированных экосистем: он производит ровно столько органики и кислорода, сколько тут же расходует в процессе своей жизнедеятельности (нулевой баланс); а ведь замкнутость геохимических циклов была ранее определена нами как главное направление в эволюции биосферы. Именно такая структура «производства и потребления» в прошлом позволяла матам образовывать кислородные оазисы и постепенно увеличивать их – вместо того, чтобы пытаться сходу изменить всю эту бескислородную среду.
Можно сказать, что в мате «ничто не пропадает», даже энергия солнечного света (разумеется, в рамках суммарного КПД фотосинтеза, составляющего около 10%). Фотосинтезирующие бактерии подкладки способны (благодаря своим пигментам) поглощать свет из иной, чем цианобактерии вышележащего слоя, части спектра. Поверхность роста становится для жителей подкладки как бы прозрачной, что позволяет утилизовать всю поступающую в фотическую зону световую энергию. Более того, поскольку солнечный спектр меняется в течение дня («покраснение» Солнца на восходе и закате), для улучшения использования энергии в мате происходят упорядоченные вертикальные миграции – смена горизонтов, занимаемых бактериями с разными типами пигментов.
Все это позволяет сделать следующий вывод: уровень интеграции, достигнутый составляющими мат микроорганизмами, превосходит уровень, наблюдаемый в обычных экосистемах, и как минимум не уступает тому, что наблюдается у лишайников. И уж коль скоро мы считаем «организмами» лишайники, то такое определение с полным основанием можно отнести и к мату. А поскольку следы жизнедеятельности матов – строматолиты – достоверно появляются в геологической летописи все в тех же древнейших осадочных формациях Варравуна и Онфервахт (3,5–3,4 млрд лет), то приходится признать следующее. Жизнь, похоже, появляется на Земле сразу в виде экосистемы, целостность которой вполне сопоставима с целостностью многоклеточного организма. Существование в природе такой замечательной формы организации, как мат (существует без особых изменений как минимум 3,5 млрд лет), позволяет по-иному взглянуть и на многоклеточность, которая так и не возникла у прокариот, – а нужна ли она им?
Как мы помним из главы 1, древнейшие из известных минералов имеют возраст 4,2 млрд лет (оценка возраста Земли в 4,5–4,6 млрд лет основана на анализе вещества метеоритов и лунного грунта). Возраст же древнейших пород, в которых найден углерод заведомо органического происхождения (в углероде, принимавшем когда-либо участие в реакциях фотосинтеза, необратимо меняется соотношение изотопов 12C и 13C) составляет... 3,8 млрд лет. Цифра, согласитесь, неслабая и сама по себе, однако тут есть еще важное дополнительное обстоятельство. Дело в том, что формацию Исуа в Гренландии, где были обнаружены эти углеродистые прослои, составляют вообще древнейшие на Земле осадочные породы. Значит, первые достоверные следы жизни появляются на Земле одновременно с первыми достоверными следами воды. А поскольку ископаемые могут сохраняться только в осадочных породах (за редчайшими исключениями, вроде захоронений под вулканическими пеплопадами и т.п.), то можно сформулировать и так: достоверные следы жизни известны в геологической летописи Земли с того самого момента, когда возникает принципиальная возможность их фиксации. Таким образом, «презумпция Вернадского» стала теперь фактически неуязвимой.
Еще недавно биологи уверено рисовали (исходя из общих соображений) такую картину. Самый длинный отрезок в истории Земли приходился на образование первых биологических систем из неорганической материи. Несколько меньшее время потребовалось на возникновение первых клеток, и лишь после этого начался все ускоряющийся процесс собственно биологической эволюции. Первыми живыми существами были гетеротрофные микроорганизмы (гетеротрофы), питавшиеся «первичным бульоном» – той органикой, что в избытке возникала в первичной атмосфере и океане в результате процессов, частично смоделированных Миллером. Затем уже возникли и автотрофы, синтезирующие органику из углекислого газа и воды, используя для этого энергию окислительных химических реакций (хемоавтотрофы) или солнечного света (фотоавтотрофы).
Однако реальные факты вынудили отказаться от этой умозрительной схемы. В числе прочего не нашли подтверждений и представления об исходной гетеротрофности живых существ; судя по всему, автотрофность и гетеротрофность возникли одновременно. Один из ведущих специалистов по микробным сообществам Г. А. Заварзин обращает внимание на то, что «первичный бульон» даже теоретически не мог быть источником пищи для «первичной жизни», т. к. является конечным, исчерпываемым ресурсом. Поскольку живые существа размножаются в геометрической прогрессии (вспомните всякого рода занимательные задачки о потомках одной бактерии, заполняющих сперва наперсток, потом железнодорожный состав и т.д.), потомство первых же гетеротрофов должно было бы сожрать весь этот «бульон» за совершенно ничтожное время; после этого все они, естественно, мрут от голода – и извольте создавать жизнь на Земле по новой. Не меньшие неприятности, впрочем, ожидают и чисто автотрофную биосферу, которая в том же темпе (и с тем же конечным результатом!) свяжет весь углерод на планете в виде неразложимых высокомолекулярных соединений.
Первичность хемоавтотрофности (относительно фотоавтотрофности) тоже принято было считать несомненной; серьезным аргументом тут является то обстоятельство, что архебактерии (наиболее архаичные из всех прокариот) – именно автотрофы. Но и здесь, как выяснилось, все не так уж очевидно. Молекула фотосинтезирующего пигмента (например, хлорофилла) поглощает квант света; в дальнейшем энергия этого кванта используется в различных химических превращениях. Так вот, есть серьезные основания полагать, что первичной функцией этих пигментов была просто-напросто нейтрализация разрушительной для организма (да и вообще для любой высокомолекулярной системы) энергии квантов ультрафиолетового излучения, беспрепятственно проникавшего в те времена сквозь лишенную озонового слоя атмосферу. Впоследствии те, кто приспособился еще и использовать эту энергию «в мирных целях», разумеется, получили гиганские преимущества, однако сама по себе «радиационная защита» ДНК при помощи пигментов должна была сформироваться еще на стадии доорганизменных гиперциклов. Понятно, что доводить эту защиту до совершенства имело смысл лишь тем из них, кто обитал в поверхностном слое океана (10-метровый слой воды полностью защищает от ультрафиолета любой интенсивности); именно они, судя по всему, и дали начало фотоавтотрофам, тогда как глубины остались в распоряжении хемоавтотрофов.
Есть и прямые доказательства существования фотоавтотрофов с самых давних времен (древнейшие организмы из формации Исуа, как мы помним, были именно фотосинтезирующими, а в породах возраста 3,1 млрд лет содержатся остатки хлорофилла – фитан и пристан – и даже неразложившийся цианобактериальный пигмент фикобилин), но гораздо проще отслеживать деятельность фотоавтотрофов по одному из ее побочных продуктов – кислороду. Впервые такой анализ был проведен Л. Беркнером и Л. Маршаллом (1962); они пришли к выводу, что количество кислорода на Земле росло на протяжении всей ее истории – постоянно, но крайне медленно, и вплоть до начала кембрия атмосфера оставалась практически бескислородной (не более 1% от его нынешнего количества). Однако данные, накопленные к настоящему моменту, заставляют предполагать, что насыщение атмосферы кислородом произошло гораздо раньше, в более отдаленные времена.
В отличие от других атмосферных газов кислород не может быть получен путем дегазации магмы, поэтому он отсутствовал в первичной атмосфере Земли, которая была восстановительной. Об этом свидетельствует наличие в древнейших, главным образом катархейских, породах легко окисляемых, но не окисленных (несмотря на пребывание на земной поверхности) веществ – таких, как графит (углерод), лазурит и т.д. Небольшие количества кислорода, образовывавшегося в результате фотодиссоциации молекул воды под действием жесткого ультрафиолетового излучения, полностью расходовались на окисление постоянно выделяемых вулканами аммиака, закиси углерода, метана и сероводорода. Однако с определенного момента в океанах начали образовываться так называемые полосчатые железные руды (джеспиллиты), представляющие собой чередование бедных и обогащенных окислами железа прослоев. Они формируются, когда двухвалентное (закисное) железо, выработавшееся в результате вулканической активности и растворенное в морской воде, реагирует в поверхностных слоях океана с молекулярным кислородом, окисляется до трехвалентного (окисного) и выпадает в виде нерастворимого осадка на дно. Таким образом, начало образования руд этого типа (состоящих в основном из гематита Fe2O3 и магнетита FeO · Fe2O3) означает появление на Земле достаточно мощного источника молекулярного кислорода, каковым могут быть только фотосинтезирующие организмы. Впервые джеспиллиты появляются все в той же формации Исуа; это может служить независимым подтверждением того, что содержащееся в ней углеродистое вещество действительно принадлежало фотосинтетикам (рис. 16).
Рис. 16. Геологические свидетельства изменений состава земной атмосферы и условий среды на протяжении докембрия и фанерозоя (по Schopf, 1992)
Источник-то кислорода возник, но мир еще на протяжении полутора миллиардов лет оставался анаэробным; об этом свидетельствует наличие в соответствующих отложениях конгломератов из пирита (FeS2). Зачастую они представляют собой гальку с отчетливыми следами обработки ее течением – это, кстати, первое свидетельство существования на Земле пресных вод. Но текучие воды не могут не быть хорошо аэрироваными, и то, что столь легко окисляемое вещество, как пирит, осталось неокисленным, свидетельствует о практически бескислородной атмосфере. Сообщества фотоавтотрофов формируют в это время своеобразные кислородные оазисы (термин П. Клауда) в бескислородной пустыне; их возможностей хватает на создание окислительных обстановок (и осаждение железа в окисной форме) лишь в своем непосредственном окружении.Ситуация радикально изменилась около 2 млрд лет назад, когда процесс гравитационной дифференциации недр (см. главу 3) привел к тому, что бо?льшая часть железа перешла в ядро планеты и растворенное в морской воде закисное железо не могло уже возобновляться по мере выбывания из цикла. Фотоавтотрофы получили возможность завершить свою титаническую работу по окислению закисного железа и осаждению его в виде джеспиллитов; именно в это время возникли все крупнейшие месторождения железа, такие, как Курская магнитная аномалия. В дальнейшем руды этого типа уже не образовывались (за исключением очень краткого эпизода в самом конце докембрия – о нем речь пойдет в главе 6). Кислород, ранее полностью расходовавшийся на этот процесс, стал теперь мало-помалу насыщать атмосферу; с этого времени пиритовые конгломераты исчезли, а на смену донно-морским железным рудам пришли терригенные (имеющие наземное происхождение) красноцветы, т.е. процесс окисления железа начался и на суше (рис. 16). Яркий образ для описания этих событий придумал Г. А. Заварзин: он назвал их временем, когда «биосфера „вывернулась наизнанку“ за счет появления кислородной атмосферы как продукта обмена цианобактерий, и вместо кислородных «карманов» появились анаэробные «карманы» в местах разложения органики».
Именно в это время (1,9 млрд лет назад) в канадской формации Ганфлинт впервые появляются звездчатые образования, полностью идентичные тем, что образует ныне облигатно-аэробная марганцевоосаждающая бактерия Metallogenium. Без кислорода окисление железа и марганца не идет, и образуемые этой бактерией металлические кристаллы в виде характерных «паучков» возникают только в сильно окислительной обстановке. Это должно означать, что в тот момент содержание кислорода в атмосфере уже достигло величины как минимум в 1% от современного (точка Пастера). Именно с этой пороговой концентрации становится «экономически оправданным» налаживание процесса кислородного дыхания, в ходе которого из каждой молекулы глюкозы можно будет получать 38 энергетических единиц (молекул АТФ) вместо двух, образующихся при бескислородном брожении. С другой стороны, в атмосфере начинает возникать озоновый слой, преграждающий путь смертоносному ультрафиолету, что ведет к колоссальному расширению спектра пригодных для жизни местообитаний.
Одним из симптомов произошедшей «кислородной революции» стало то, что у автотрофов возникли серьезные проблемы с азот-фиксацией, т.е. переводом атмосферного азота в усвояемую, ионную форму. Азотфиксация возможна только в бескислородных условиях или при низком содержании кислорода, поэтому у автотрофов (таких, как цианобактерии), выделяющих кислород в процессе собственной жизнедеятельности, азотфиксация идет в специальных клетках с «герметической» утолщенной оболочкой – гетероцистах и акинетах. Акинеты впервые появляются в палеонтологической летописи как раз после «кислородной революции» – около 1,4 млрд лет назад.
Примерно к середине протерозоя (1,7–1,8 млрд лет назад) «кислородная революция» в целом завершается и мир становится аэробным (рис. 16). Впрочем, с точки зрения существ, составлявших тогдашнюю биосферу, этот процесс следовало бы назвать иначе: необратимое отравление кислородом атмосферы планеты. Смена анаэробных условий на аэробные не могла не вызвать катастрофических перемен в структуре тогдашних экосистем. В действительности «кислородная революция» есть не что иное, как первый в истории Земли глобальный экологический кризис.
Во всех предыдущих построениях мы имели дело лишь с геохимическими следствиями тех процессов, что происходили на Земле при предполагаемом нами участии живых существ. Но располагаем ли мы хоть какой-то информацией о самих этих существах? Можно ли их «пощупать пальцами»? Оказывается, можно. В последние десятилетия были разработаны специальные методы обработки осадочных горных пород, позволяющие выделять содержащиеся в них клеточные оболочки, а в некоторых случаях даже получать косвенную информацию о внутреннем строении этих клеток. К настоящему моменту в докембрийских породах обнаружено множество одноклеточных организмов, древнейшие из них – в местонахождениях Варравуна в Австралии (3,5 млрд лет) и Онфервахт в Южной Африке (3,4 млрд лет). Это оказались несколько видов цианобактерий (сине-зеленых водорослей), ничем особенно не отличающихся от современных; мы уже упоминали о том, что из пород возраста 3,1 млрд лет был выделен пигмент фикобилин, который используется при фотосинтезе современными цианобактериями (никем, кроме них).
Итак, в раннем докембрии существовал совершенно особый мир, формируемый прокариотными организмами – бактериями и циано-бактериями. Разделение живых существ на прокариоты и эукариоты (эти термины были введены в 1925 году Э. Шаттоном), основанное на наличии или отсутствии в их клетках оформленного ядра, теперь считают существенно более фундаментальным, чем, например, разделение на «животные» и «растения». Из многих их различий для нас сейчас важнее всего то, что среди прокариот неизвестны многоклеточные организмы. Есть нитчатые и пальмеллоидные[12] формы цианобактерий, однако уровень интеграции клеток в этих структурах – это все-таки уровень колонии, а не организма. А поскольку времени на всякого рода эксперименты у прокариот, как мы видели, было более чем достаточно (3,5 млрд лет), то приходится предположить, что по каким-то причинам на прокариотной основе многоклеточность не возникает в принципе. Может быть, дело в отсутствии у них центриолей и митотического веретена, без которого невозможно точно ориентировать в пространстве делящиеся клетки и формировать из них сколь-нибудь сложные ансамбли. Может быть – в свойствах их клеточной оболочки, препятствующих межклеточным взаимодействиям; это делает невозможным обмен веществами, так что клетки, находящиеся внутри организма и не имеющие прямого контакта с окружающей средой (что неизбежно при многоклеточности), не могли бы питаться и выводить продукты распада. Может быть – в чем-то еще. Как бы то ни было, такой путь повышения уровня организации, как многоклеточность, для прокариот оказался закрытым. Выход из этого тупика они нашли весьма оригинальный.
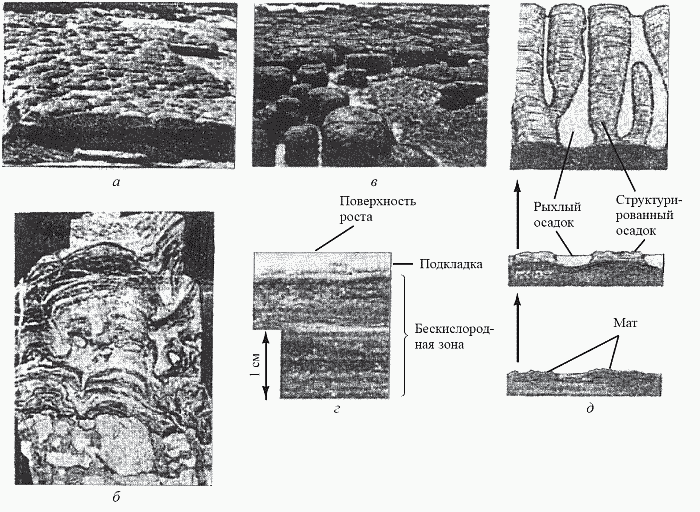
Для докембрийских осадочных толщ чрезвычайно характерны строматолиты (с греческого – «каменный ковер») – тонкослойчатые колонны или холмики, состоящие главным образом из карбоната кальция (рис. 17, а и б). Эти структуры известны еще с середины XIX века, однако их происхождение оставалось совершенно неясным до 30-х годов XX века, когда было сделано замечательнейшее открытие в области классической (т.е. немолекулярной) биологии. На литорали залива Шарк-Бэй в западной Австралии и на атлантическом побережье Багамских островов были найдены небольшие рифовые постройки неизвестного ранее типа – плосковершинные известковые «бочки» диаметром 1–1,5 м (рис. 17, в) и располагающиеся ниже уровня отлива слоистые корки; все это при внимательном рассмотрении оказалось... современными строматолитами. Выяснилось, что строматолит образуется в результате жизнедеятельности совершенно ни на что не похожего прокариотного сообщества, называемого цианобактериальным матом. Маты существуют во многих районах мира, главным образом в таких пересоленных лагунах, как Сиваш, однако настоящие строматолиты (как в Шарк-Бэй и на Багамской отмели) они образуют довольно редко.
Рис. 17. Маты и строматолиты, современные и ископаемые:
а – ископаемые строматолиты, общий вид; б – они же, в разрезе; в – современные строматолитовые постройки в Шарк-Бэй, Австралия; г – поперечный разрез мата; д – образование строматолита матом (схема) (по Schopf, 1992, и Dzik, 1997)
Мат, располагающийся на верхней поверхности создаваемого им строматолита, представляет собой плотный многослойный «ковер» общей толщиной до 2 см; основу его составляют нитчатые либо пальмеллоидные цианобактерии, но помимо них в формировании сообщества участвует множество самых различных микроорганизмов. Внутри мата легко различимы несколько функционально дискретных слоев (рис. 17, г):1. Плотный верхний слой – поверхность роста (1–1,5 мм), в котором есть и автотрофы (продуцирующие кислород цианобактерии-фотосинтетики), и гетеротрофы (облигатно-аэробные, т.е. неспособные жить в отсутствие кислорода, бактерии).
2. Тонкая (менее 1 мм) подкладка; в ней автотрофами служат бактерии, осуществляющие бескислородный фотосинтез (при котором источником водорода является не вода, а другие вещества, например, сероводород – тогда выделяется не молекулярный кислород, а сера). Гетеротрофами же служат аэробы (факультативные), которые могут использовать кислород, но в его отсутствие способны удовлетворять свои потребности в энергии не дыханием, а брожением.
3. Ниже двух верхних слоев, в которых идут процессы фотосинтеза и которые в совокупности называют фотической зоной мата, лежит толстая бескислородная зона, где процветают разнообразные анаэробы: для них молекулярный кислород не нужен или даже является ядом.
На поверхность обитающего на мелководье мата постоянно выпадают частички осадка (обычно кристаллы карбоната кальция), которые затемняют соответствующе участки фотической зоны и должны постепенно ухудшать условия идущего в ней фотосинтеза – вплоть до неизбежной гибели фотосинтетиков. От катастрофы эти микроорганизмы спасает положительный фототаксис: по мере того, как интенсивность света в фотической зоне падает, они мигрируют вверх сквозь накапливающийся слой осадка, образуя выше него новую поверхность роста с подкладкой. Миграция эта идет разными способами: нитчатые формы обладают способностью к скользящему движению сквозь осадок, пальмеллоидные – к ускоренному нарастанию верхних частей колонии; еще один путь – временный переход в латентное состояние с последующей реколонизацией поверхности осадка.
Оказавшиеся таким образом внутри бескислородной зоны мата осадки структурируются и слой за слоем наращивают верхнюю поверхность строматолита; измерения показали, что современные строматолиты прирастают со скоростью около 0,3 мм/год. Каждый слой состоит из пары слойков – светлого и темного, состоящих, соответственно, из более мелких и более крупных кристаллов карбоната кальция; крупные кристаллы образуются в тот сезон года, когда процесс осаждения идет быстрее (рис. 17, д). Следует подчеркнуть, что вещество, из которого построен стоматолит, не создается матом – последний лишь структурирует естественное осадконакопление. Строматолиты чаще всего состоят из карбоната кальция лишь потому, что карбонатный тип осадконакопления в море наиболее обычен, однако в иных гидрохимических условиях формируются строматолиты фосфатные, кремнезёмные, железистые и др.[13]
Не все маты представляли собой донные сообщества. Помимо уже известных нам строматолитов, следы жизнедеятельности микробных сообществ представлены еще и онколитами – более или менее сферическими образованиями, в которых известковые слои располагаются не линейно (как в строматолите); они имеют концентрическую структуру. Предполагается, что онколиты создавались особыми прокариотными сообществами, похожими на плавающие в толще воды шары, оболочка которых являлась матом; осадки отлагались кнутри от оболочки этого «батискафа», а плавучесть ему придавали образуемые при метаболизме газы, не могущие покинуть замкнутую полость внутри колонии. Позже мы столкнемся с особым типом водорослевого сообщества – плавающим матом, характерным для некоторых внутренних водоемов мезозоя (см. главу 9).
Итак, мат – это высокоинтегрированное сообщество с чрезвычайно сложно организованной трофической структурой: два верхних слоя составлены продуцентами и консументами первого порядка («растениями» и «травоядными»), соединенными в пищевую цепь пастбищного типа, а нижний слой сформирован редуцентами («падальщиками»), получающими органику из верхних слоев по детритной цепи (рис. 17). Более того, измерения показали, что мат является вообще одной из самых сбалансированных экосистем: он производит ровно столько органики и кислорода, сколько тут же расходует в процессе своей жизнедеятельности (нулевой баланс); а ведь замкнутость геохимических циклов была ранее определена нами как главное направление в эволюции биосферы. Именно такая структура «производства и потребления» в прошлом позволяла матам образовывать кислородные оазисы и постепенно увеличивать их – вместо того, чтобы пытаться сходу изменить всю эту бескислородную среду.
Можно сказать, что в мате «ничто не пропадает», даже энергия солнечного света (разумеется, в рамках суммарного КПД фотосинтеза, составляющего около 10%). Фотосинтезирующие бактерии подкладки способны (благодаря своим пигментам) поглощать свет из иной, чем цианобактерии вышележащего слоя, части спектра. Поверхность роста становится для жителей подкладки как бы прозрачной, что позволяет утилизовать всю поступающую в фотическую зону световую энергию. Более того, поскольку солнечный спектр меняется в течение дня («покраснение» Солнца на восходе и закате), для улучшения использования энергии в мате происходят упорядоченные вертикальные миграции – смена горизонтов, занимаемых бактериями с разными типами пигментов.
Все это позволяет сделать следующий вывод: уровень интеграции, достигнутый составляющими мат микроорганизмами, превосходит уровень, наблюдаемый в обычных экосистемах, и как минимум не уступает тому, что наблюдается у лишайников. И уж коль скоро мы считаем «организмами» лишайники, то такое определение с полным основанием можно отнести и к мату. А поскольку следы жизнедеятельности матов – строматолиты – достоверно появляются в геологической летописи все в тех же древнейших осадочных формациях Варравуна и Онфервахт (3,5–3,4 млрд лет), то приходится признать следующее. Жизнь, похоже, появляется на Земле сразу в виде экосистемы, целостность которой вполне сопоставима с целостностью многоклеточного организма. Существование в природе такой замечательной формы организации, как мат (существует без особых изменений как минимум 3,5 млрд лет), позволяет по-иному взглянуть и на многоклеточность, которая так и не возникла у прокариот, – а нужна ли она им?