Страница:
Касаясь совершенства баланса между процессами синтеза и деструкции, осуществляемыми в мате, уместно вспомнить еще одно высказывание Вернадского, сделанное им в 1931 году: «Первое появление жизни при создании биосферы должно было произойти не в виде появления одного какого-либо организма, а в виде их совокупности, отвечающей геохимическим функциям жизни». Похоже, что и это предсказание теперь тоже находит подтверждение.
Каковы же были условия существования этих древнейших прокариотных экосистем? Они традиционно считались подводными (морскими) до тех пор, пока И. Н. Крылов и Г. А. Заварзин (1988) не выдвинули ряд аргументов в пользу наземного образования докембрийских строматолитов. Палеонтолог А. Г. Пономаренко (1993) задался таким вопросом: что такое вообще «море» и «суша» применительно к глубокому докембрию? Раньше это просто никому не приходило в голову, хотя постановка этого вопроса абсолютно правомерна. В отсутствие высшей наземной растительности скорость водной и ветровой эрозии должна превышать современную во много раз. Незащищенные наклонные субстраты при этом будут размываться очень быстро; ландшафт окажется состоящим из практически вертикальных скал и почти горизонтальных равнин, лишь незначительно превышающих уровень моря. Так что, по всей видимости, в те времена на окраинах континентов формировались обширные мелководные бассейны с постоянно меняющимся уровнем воды и без настоящей береговой линии. Именно в таком «вымершем» ландшафте (это не было ни сушей, ни морем в современном смысле) и процветали строматолитовые экосистемы.
Помимо бентосных прокариотных экосистем, представленных матами, все это время существовали и планктонные, состоящие из сферических одноклеточных водорослей – акритарх[14] – и шарообразных колоний, напоминающих современный Volvox. Первые планктонные организмы появляются в палеонтологической летописи 3,5 млрд лет назад – одновременно со строматолитами.
Именно в планктонных экосистемах в протерозое (около 1,9–2 млрд лет назад) произошло то, что стало ключевым для дальнейшей судьбы биосферы: в составе фитопланктона появились такие акритархи, которых считают первыми на Земле эукариотами. Заключение об их эукариотности первоначально основывалось на размере клеток (они превышают 60 мкм, тогда как практически все известные прокариоты со сферическими клетками меньше 10 мкм). Однако позже на Среднесибирском плоскогорье (местонахождение Мирое-диха) было собрано множество экземпляров уникальной сохранности, на которых удалось реконструировать процес ядерного деления. А недавно в одном из австралийских нефтяных месторождений сходного возраста (1,7 млрд лет) были обнаружены стеролы – вещества, которые синтезируются только клеточными ядрами.
Гипотеза выглядит достаточно фантастично, однако имеет серьезные обоснования. Аналогичные процессы происходят и в современном мире. Например, инфузория-туфелька может содержать в качестве «домашнего животного» зеленую водоросль хлореллу. Инфузория не трогает «домашнюю» хлореллу, но немедленно переваривает любую «дикую» клетку того же вида. Хлорелла же образует внутри хозяина строго фиксированное число клеток и через несколько поколений теряет способность к самостоятельному существованию.
С другой стороны, сходство митохондрий и хлоропластов с бактериями – не только внешнее (и те и другие – овальные тела, окруженные двойной мембраной, на складках внутреннего слоя которой локализованы пигменты с дыхательными или фотосинтетическими функциями). Эти органеллы имеют собственную ДНК, причем не линейную (как хромосомная ДНК самой эукариотной клетки), а кольцеобразную – такую же, как у настоящих бактерий. В момент клеточного деления митохондрии и хлоропласты не создаются клеткой заново, а размножаются сами, как настоящие одноклеточные организмы. При этом степень их самостоятельности такова, что они вполне способны существовать и размножаться и в чужих клетках: например, хлоропласты легко размножаются в курином яйце.
Сравнив с этих позиций эукариотную клетку с матом, можно заключить, что мы имеем дело со второй (и столь же успешной) попыткой прокариот создать высокоинтегрированную колонию, на этот раз не между различными клетками, а внутри одной из них. Похоже, что, не имея возможности выработать истиную многоклеточность, прокариоты выжали все, что можно, из симбиоза: раз нельзя наладить должную дифференцировку собственных клеток, то почему бы не использовать в качестве строительного материала клетки разных организмов? Не исключено, что «конструкторские разработки» по обоим «проектам» (мат и эукариотность) первые прокариоты начали одновременно. Такое предположение может хорошо объяснить тот парадоксальный результат, к которому пришли в последние годы К. Вёзе и ряд других исследователей: сопоставив нуклеотидные последовательности различных групп организмов методом молекулярных часов[15] , они заключили, что эукариоты должны были возникнуть столь же давно, как и прокариоты.
Скорее всего, различные варианты эукариотности, т.е. внутриклеточных колоний, возникали многократно (например, есть основания полагать, что красные водоросли, резко отличающиеся от всех прочих растений по множеству ключевых признаков, являются результатом такой «независимой эукариотизации» цианобактерий), однако все эти варианты, видимо, не имели в тогдашних условиях должного преимущества перед прокариотным типом организации и «не выходили в серию». Ведь эукариотам приходится платить за свою сложность весьма дорогую цену: они потеряли присущие прокариотам «бессмертие» (видовую неизменность на протяжении миллиардов лет) и «неуязвимость» (способность жить в кипятке или в ядерном реакторе, питаться любыми ядами или чистым водородом и т.д.); какой же смысл после этого выполнять в биосфере те функции, для которых достаточно и прокариот? Если компьютер нужен вам лишь для печатания текстов и игры в «Тетрис», то вы, надо думать, не станете покупать Пентиум со всеми «наворотами», а ограничитесь чем попроще.
Ситуация радикально изменилась около 2 млрд лет назад, когда появление одного из возможных вариантов эукариотности (аэробный фотоавтотроф) совпало с другим событием: в результате деятельности цианобактерий концентрация кислорода в атмосфере Земли достигла точки Пастера, и в этих условиях эукариотная «модель» наконец оказалась «конкурентоспособной». Именно эукариоты-аэробы со временем оттеснят прокариотные сообщества в «резервации» (пересоленные водоемы, горячие источники и т.д.) или превратят их в эфемеров (сине-зеленые водоросли, стремительно размножающиеся в луже и тут же исчезающие вместе с ней).
Это выглядит, если вдуматься, вопиющей исторической несправедливостью. Здесь мы в первый раз (но далеко не в последний) сталкиваемся с ситуацией, когда некая группа живых организмов постепенно изменяет окружающий мир «под себя», а плодами этих изменений пользуются другие, причем с достаточно печальным для «инициаторов реформ» результатом. Увы! Блестяще сформулированный Стругацкими принцип: «Будущее создается тобой, но не для тебя» – проявляется в эволюции биосферы как одна из фундаментальнейших закономерностей.
Впрочем, эукариоты начали свое триумфальное шествие далеко не сразу. Появившись почти 2 млрд лет назад, они на протяжении почти миллиарда лет не играли сколь-нибудь заметной роли в экосистемах, а все разнообразие этих организмов было ограничено фитопланктонными формами – акритархами. Создается впечатление, что сама по себе эукариотность еще не дает решающего преимущества; до тех пор, пока эукариоты остаются одноклеточными, они лишь конкурентоспособны (относительно прокариот) – но не более того. Мир продолжает оставаться прокариотным вплоть до конца протерозоя. Однако около 800 млн лет назад наступает эпоха поистине драматических перемен, о которых и пойдет речь в следующей главе.
Глава 6
Каковы же были условия существования этих древнейших прокариотных экосистем? Они традиционно считались подводными (морскими) до тех пор, пока И. Н. Крылов и Г. А. Заварзин (1988) не выдвинули ряд аргументов в пользу наземного образования докембрийских строматолитов. Палеонтолог А. Г. Пономаренко (1993) задался таким вопросом: что такое вообще «море» и «суша» применительно к глубокому докембрию? Раньше это просто никому не приходило в голову, хотя постановка этого вопроса абсолютно правомерна. В отсутствие высшей наземной растительности скорость водной и ветровой эрозии должна превышать современную во много раз. Незащищенные наклонные субстраты при этом будут размываться очень быстро; ландшафт окажется состоящим из практически вертикальных скал и почти горизонтальных равнин, лишь незначительно превышающих уровень моря. Так что, по всей видимости, в те времена на окраинах континентов формировались обширные мелководные бассейны с постоянно меняющимся уровнем воды и без настоящей береговой линии. Именно в таком «вымершем» ландшафте (это не было ни сушей, ни морем в современном смысле) и процветали строматолитовые экосистемы.
Помимо бентосных прокариотных экосистем, представленных матами, все это время существовали и планктонные, состоящие из сферических одноклеточных водорослей – акритарх[14] – и шарообразных колоний, напоминающих современный Volvox. Первые планктонные организмы появляются в палеонтологической летописи 3,5 млрд лет назад – одновременно со строматолитами.
Именно в планктонных экосистемах в протерозое (около 1,9–2 млрд лет назад) произошло то, что стало ключевым для дальнейшей судьбы биосферы: в составе фитопланктона появились такие акритархи, которых считают первыми на Земле эукариотами. Заключение об их эукариотности первоначально основывалось на размере клеток (они превышают 60 мкм, тогда как практически все известные прокариоты со сферическими клетками меньше 10 мкм). Однако позже на Среднесибирском плоскогорье (местонахождение Мирое-диха) было собрано множество экземпляров уникальной сохранности, на которых удалось реконструировать процес ядерного деления. А недавно в одном из австралийских нефтяных месторождений сходного возраста (1,7 млрд лет) были обнаружены стеролы – вещества, которые синтезируются только клеточными ядрами.
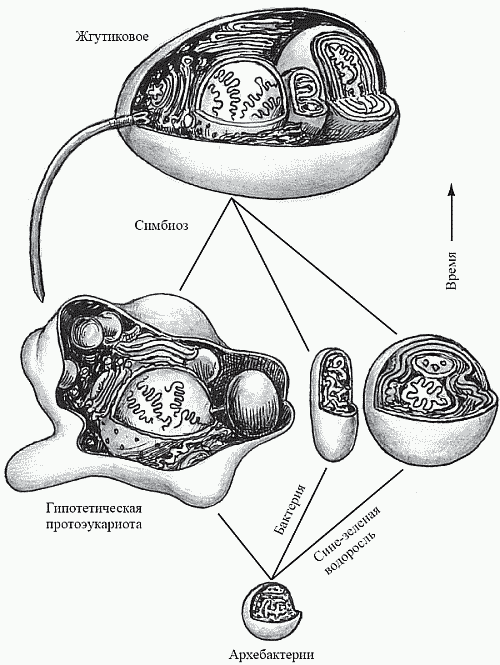
Рис. 18. Схематическое изображение симбиогенеза (по Dzik, 1997)
По поводу происхождения эукариот большинство исследователей придерживается гипотезы симбиогенеза (рис. 18). Идея о том, что растительная и животная клетки являют собой «симбиотический комплекс», была впервые высказана К. С. Мережковским (1909) и затем обоснована А. С. Фаминцыным (который ранее, в 1867 году, доказал симбиотическую природу лишайников); автором же концепции в ее современном виде считается Л. Маргулис. Концепция состоит в том, что органеллы, наличие которых отличает эукариотную клетку от прокариотной (митохондрии, хлоропласты и жгутики с базальным телом и микротрубочками), являются результатом эволюции некогда независимых прокариотных клеток, которые были захвачены клеткой-хозяином (тоже прокариотной) и не «съедены», а превращены ею в симбионтов. Предполагается, что роль клетки-хозяина выполняла крупная факультативно-анаэробная бактерия-гетеротроф. Проглотив однажды мелких аэробных бактерий-гетеротрофов, она начала использовать их в качестве «энергетических станций», перерабатывая с их помощью свою органику по более совершенной технологии (дыхание вместо брожения), что позволило ей получать из каждой молекулы глюкозы 38 молекул АТФ вместо двух. Союз был выгоден и для «рабов» – аэробов, получивших взамен гораздо более совершенный источник «топлива»: органику, добываемую крупным хищным хозяином. Далее к поверхности хозяина прикрепилась другая группа симбионтов – жгутикоподобные бактерии (Маргулис полагает, что это было нечто вроде современных спирохет), которые резко увеличили подвижность хозяина – в обмен на возможность «подсоединиться к единой энергосистеме». Поглотив затем подходящих мелких фотоавтотрофов – цианобактерий, хозяин обрел возможность получать органику не путем активного поиска ее во внешней среде, а просто, так сказать, посидев немного на солнышке. Этот переход до некоторой степени аналогичен происходившему в человеческой истории переходу от охоты и собирательства к скотоводству и земледелию.Гипотеза выглядит достаточно фантастично, однако имеет серьезные обоснования. Аналогичные процессы происходят и в современном мире. Например, инфузория-туфелька может содержать в качестве «домашнего животного» зеленую водоросль хлореллу. Инфузория не трогает «домашнюю» хлореллу, но немедленно переваривает любую «дикую» клетку того же вида. Хлорелла же образует внутри хозяина строго фиксированное число клеток и через несколько поколений теряет способность к самостоятельному существованию.
С другой стороны, сходство митохондрий и хлоропластов с бактериями – не только внешнее (и те и другие – овальные тела, окруженные двойной мембраной, на складках внутреннего слоя которой локализованы пигменты с дыхательными или фотосинтетическими функциями). Эти органеллы имеют собственную ДНК, причем не линейную (как хромосомная ДНК самой эукариотной клетки), а кольцеобразную – такую же, как у настоящих бактерий. В момент клеточного деления митохондрии и хлоропласты не создаются клеткой заново, а размножаются сами, как настоящие одноклеточные организмы. При этом степень их самостоятельности такова, что они вполне способны существовать и размножаться и в чужих клетках: например, хлоропласты легко размножаются в курином яйце.
Сравнив с этих позиций эукариотную клетку с матом, можно заключить, что мы имеем дело со второй (и столь же успешной) попыткой прокариот создать высокоинтегрированную колонию, на этот раз не между различными клетками, а внутри одной из них. Похоже, что, не имея возможности выработать истиную многоклеточность, прокариоты выжали все, что можно, из симбиоза: раз нельзя наладить должную дифференцировку собственных клеток, то почему бы не использовать в качестве строительного материала клетки разных организмов? Не исключено, что «конструкторские разработки» по обоим «проектам» (мат и эукариотность) первые прокариоты начали одновременно. Такое предположение может хорошо объяснить тот парадоксальный результат, к которому пришли в последние годы К. Вёзе и ряд других исследователей: сопоставив нуклеотидные последовательности различных групп организмов методом молекулярных часов[15] , они заключили, что эукариоты должны были возникнуть столь же давно, как и прокариоты.
Скорее всего, различные варианты эукариотности, т.е. внутриклеточных колоний, возникали многократно (например, есть основания полагать, что красные водоросли, резко отличающиеся от всех прочих растений по множеству ключевых признаков, являются результатом такой «независимой эукариотизации» цианобактерий), однако все эти варианты, видимо, не имели в тогдашних условиях должного преимущества перед прокариотным типом организации и «не выходили в серию». Ведь эукариотам приходится платить за свою сложность весьма дорогую цену: они потеряли присущие прокариотам «бессмертие» (видовую неизменность на протяжении миллиардов лет) и «неуязвимость» (способность жить в кипятке или в ядерном реакторе, питаться любыми ядами или чистым водородом и т.д.); какой же смысл после этого выполнять в биосфере те функции, для которых достаточно и прокариот? Если компьютер нужен вам лишь для печатания текстов и игры в «Тетрис», то вы, надо думать, не станете покупать Пентиум со всеми «наворотами», а ограничитесь чем попроще.
Ситуация радикально изменилась около 2 млрд лет назад, когда появление одного из возможных вариантов эукариотности (аэробный фотоавтотроф) совпало с другим событием: в результате деятельности цианобактерий концентрация кислорода в атмосфере Земли достигла точки Пастера, и в этих условиях эукариотная «модель» наконец оказалась «конкурентоспособной». Именно эукариоты-аэробы со временем оттеснят прокариотные сообщества в «резервации» (пересоленные водоемы, горячие источники и т.д.) или превратят их в эфемеров (сине-зеленые водоросли, стремительно размножающиеся в луже и тут же исчезающие вместе с ней).
Это выглядит, если вдуматься, вопиющей исторической несправедливостью. Здесь мы в первый раз (но далеко не в последний) сталкиваемся с ситуацией, когда некая группа живых организмов постепенно изменяет окружающий мир «под себя», а плодами этих изменений пользуются другие, причем с достаточно печальным для «инициаторов реформ» результатом. Увы! Блестяще сформулированный Стругацкими принцип: «Будущее создается тобой, но не для тебя» – проявляется в эволюции биосферы как одна из фундаментальнейших закономерностей.
Впрочем, эукариоты начали свое триумфальное шествие далеко не сразу. Появившись почти 2 млрд лет назад, они на протяжении почти миллиарда лет не играли сколь-нибудь заметной роли в экосистемах, а все разнообразие этих организмов было ограничено фитопланктонными формами – акритархами. Создается впечатление, что сама по себе эукариотность еще не дает решающего преимущества; до тех пор, пока эукариоты остаются одноклеточными, они лишь конкурентоспособны (относительно прокариот) – но не более того. Мир продолжает оставаться прокариотным вплоть до конца протерозоя. Однако около 800 млн лет назад наступает эпоха поистине драматических перемен, о которых и пойдет речь в следующей главе.
Глава 6
Поздний докембрий: возникновение многоклеточности. Гипотеза кислородного контроля. Эдиакарский эксперимент.
Прежде чем непосредственно приступать к изучению древнейших многоклеточных организмов, давайте задумаемся: а зачем, собственно говоря, эта многоклеточность нужна? Какие преимущества она дает? На эту проблему можно посмотреть с разных сторон. Начнем с экологической точки зрения: что многоклеточность дает не самому организму, а экосистеме и (в конечном счете) биосфере.
Малоразмерные фитопланктонные организмы (прежде всего прокариотные) сталкиваются с одной проблемой, на которую впервые обратил внимание гидробиолог Б. Я. Виленкин. Снабжение этих одноклеточных биогенами и растворенными газами происходит за счет диффузии сквозь клеточную стенку. Казалось бы, это должно благоприятствовать миниатюризации клеток, максимальному увеличению отношения ее поверхности к объему. Однако очень мелкий организм неспособен выделиться из окружающей его водной массы. Пассивно паря в толще воды и оставаясь неподвижным относительно нее, он быстро создает вокруг себя «пустыню» – выедает из непосредственно окружающего его водного слоя все биогены, насыщая воду диффундирующей наружу органикой, которую не в силах удерживать внутри клетки из-за того же (высокого) соотношения «поверхность – объем». При этом безвозвратно теряется более 1/3 клеточной продукции. Но хуже то, что на окисление этой органики расходуется кислород и водный слой, окружающий организм, эвтрофицируется. Милый Пятачок прав: жизнь «очень маленького существа» полна неприятностей.
Организм избежит неприятностей, если будет перемещаться относительно вмещающей его воды. Этого он может добиться двумя способами: либо начнет сам активно двигаться в ее толще, либо прикрепится к неподвижному субстрату, чтобы вода двигалась относительно него. Второй способ прокариоты реализовали немедленно, сконструировав мат. А вот с первым способом (казалось бы, более простым и очевидным) у них возникли большие проблемы, ибо эффективных органов движения (подобных жгутикам и ресничкам эукариотных одноклеточных) у прокариот не возникает, а создание клеточных агрегаций, способных к согласованным движениям (например, волнообразным) затруднено из-за крайней слабости межклеточных взаимодействий (см. главу 5). Поэтому «генеральной линией» этот способ становится лишь при появлении эукариот, способных к образованию высокоинтегрированных клеточных агрегаций, т.е. к настоящей многоклеточности.
Многоклеточность привела, среди прочего, к резкому повышению способности организмов создавать в своем теле запас питательных веществ. Практически лишенные этих запасов одноклеточные (в особенности прокариоты) обречены реагировать на изменения содержания биогенов в окружающей среде единственным способом – изменением плотности популяций. (Примером такого их поведения может служить «цветение» водоемов, когда стремительное размножение одноклеточных и нитчатых водорослей быстро приводит к исчерпанию ресурса экосистемы; в результате безмерно разросшаяся популяция в одночасье вымирает, а на окисление этой мертвой органики расходуется затем почти весь кислород водоема – «замор».) Запасные вещества и резервная биомасса крупных организмов делают их популяции более независимыми от колебаний ресурса и стабилизируют их плотность. В свою очередь, существующие в экосистеме геохимические круговороты обретают в лице этих организмов крупное резервное депо[16] , оказывающее на все эти циклы мощное стабилизирующее воздействие. Такая «взаимная стабилизация» кажется одним из главных экологических следствий возникновения многоклеточных (хотя правильнее сказать – макроскопических, т.е. видимых невооруженным глазом) организмов.
Последняя оговорка неслучайна. Если мы от вопроса «для чего возникла многоклеточность?» перейдем к вопросу «как и когда она возникла?», то нам прежде всего придется определить, какой организм следует считать многоклеточным. Если даже не брать в расчет чисто терминологические проблемы (следует ли называть «колонией» Volvox, у которого есть дифференциация между клетками переднего и заднего полушарий, зародыши, возникающие путем палинтомии – деления без последующего увеличения размеров клеток, и который к тому же смертен ), здесь существует и проблема вполне объективная. А именно: соответствует ли строгим критериям многоклеточности строение тела макроскопических водорослей и грибов? Большинство биологов ныне отвечают на этот вопрос отрицательно, оставляя термин «многоклеточные» лишь за зелеными растениями (Metaphyta) и животными (Metazoa); Дж. Корлисс (1983) ввел для этих двух групп удачный термин – многотканевые организмы.
Таким образом, достижение живыми организмами макроскопических размеров может происходить различными способами, а истиная многоклеточность (многотканевость) – лишь один из них. В любом случае, многоклеточность (в широком смысле) почти наверняка возникала в процессе эволюции многократно и независимо: во многих типах водорослей – красных (Rhodophyta), зеленых (Chlorophyta), золотистых (Chrysophyta) – или высших грибов-аскомицетов (Ascomyceta) можно выявить преемственные ряды от одноклеточных форм через колонии (например, нитчатые формы) к макроскопическим организмам с аналогами тканевой дифференцировки.
Эта гипотетическая эволюционная картина вполне соответствует современным палеонтологическим даным (рис. 19). Первые эукариоты появились (как мы знаем из главы 5) около 2 млрд лет назад среди фитопланктонных акритарх; вскоре к ним добавились и нитчатые формы с эукариотными параметрами клеток. Замечательно, что они никогда не встречаются в прокариотных бентосных сообществах (цианобактериальных матах), а с самого начала формируют свой собственный тип растительности, названный У. Шенборном (1987) «водорослевыми лугами». Ныне сообщества такого типа известны лишь в некоторых антарктических внутренних водоемах; в докембрии же они, судя по характеру осадков, были широко распространены в морях за пределами мелководий (которые были заняты матами).
В рифее (1,4–1,2 млрд лет назад) эти нитчатые эукариотные формы достигли значительного разнообразия. Именно тогда в составе водорослевых лугов появились и первые макроскопические водоросли с пластинчатым, корковым и кожистым типами слоевища, а к венду (650 млн лет) основным компонентом этих сообществ становятся вендотении – лентовидные водоросли длиной до 15 см. Есть даже сообщение о находке каких-то пластинчатых водорослей в китайской формации Чанчен с возрастом 1,8 млрд лет; эта датировка нуждается в подтверждении, но не кажется нереальной. Дело в том, что таксономическая принадлежность всех этих форм недостаточно ясна, однако некоторые из них очень сходны с низшими красными водорослями – бангиевыми. А красные водоросли, как полагают многие исследователи, являются самой архаичной ветвью эукариот; возможно, они произошли в результате «независимой эукариотизации» цианобактерий.
Однопроцентное содержание кислорода (имеется в виду 1% от его современного количества) – это тот критический минимум, ниже которого аэробный метаболизм принципиально невозможен; однако для жизнедеятельности макроскопических животных кислорода необходимо существенно больше. Б. Раннегар недавно провел специальные расчеты, из которых следует, что животным, составлявшим первую фауну многоклеточных – эдиакарскую (о ней речь впереди), кислорода требовалось не менее 6–10% от нынешнего уровня; это в том случае, если они имели развитую систему циркуляции, доставлявшую кислород к тканям. Если же такая система у них еще не развилась и они дышали за счет прямой диффузии (скорее всего, так и было), то требовалось для их жизнедеятельности гораздо больше кислорода (может быть, его содержание сопоставимо с нынешним). Итак, гипотеза «кислородного контроля» кажется вполне логичным объяснением появления макроскопических животных лишь в конце протерозоя – если принять более высокий критический порог, чем однопроцентный, предлагавшийся Беркнером и Маршаллом. Однако возможна ли проверка этой гипотезы? Мы помним (из главы 5), что можно по составу осадков отличить аэробные обстановки от анаэробных, но как определить количественные различия в содержании кислорода в былые эпохи?
Известно, что количество кислорода, создаваемого небиологическими процессами (фотолиз воды и т.д.), совершенно ничтожно; почти весь свободный кислород планеты создан фотосинтезирующими организмами. Однако живые существа не только производят кислород, но и потребляют его при дыхании. В биосфере осуществляется достаточно простая химическая реакция: nСО2+nH2O?(CH2O)n + nО2. «Читая» ее слева направо, мы получаем фотосинтез, а справа налево – дыхание (а также горение и гниение). Уровень содержания кислорода на планете стабилен потому, что прямая и обратная реакции взаимно уравновешиваются; так что если мы попытаемся увеличить содержание свободного кислорода в атмосфере путем простого наращивания объема фотосинтезирующего вещества, то из этой затеи ничего не выйдет. Сместить химическое равновесие, как вам должно быть известно из курса химии, можно, лишь выводя из сферы реакции один из ее продуктов. В нашем случае добиться увеличения выхода О2 можно, лишь необратимо изымая из нее восстановленный углерод в форме (CH2O)n или его производных.
Таким образом, производство кислорода биосферой начинает превалировать над потреблением этого газа (ею же), только если происходит захоронение в осадках неокисленного органического вещества. Этот вывод кажется достаточно парадоксальным и противоречащим расхожим представлениям. Так, например, природоохранная пропаганда вбила в голову широкой публики магическую фразу «Леса – это легкие планеты»; и мало кто дает себе труд задуматься над тем, что в действительности любое сбалансированное сообщество (в том числе тропический лес) потребляет ровно столько кислорода, сколько и производит. Если что и можно назвать «легкими планеты», так это болота, где как раз и идет процесс захоронения неокисленной органики. Поэтому, если мы установим, что в некий период геологической истории происходило интенсивное захоронение органического углерода, то мы вправе будем заключить, что в это время столь же интенсивно накапливался и кислород. А вот оценить темпы захоронения органического углерода в прошлые эпохи вполне возможно. Для этого существует специальный метод, основанный на изучении изотопного отношения 12C/13C в соответствующих осадках (органический углерод, участвовавший когда-либо в реакциях фотосинтеза, обогащен «легким» изотопом 12C).
Несколько лет назад на архипелаге Шпицберген была открыта уникальная по полноте последовательность позднепротерозойских осадков, отлагавшихся в период с 850 до 600 млн лет назад. Исследовав эти осадки на предмет изотопного отношения 12C/13C, Э. Нолль (1996) установил, что на протяжении всего этого времени темпы захоронения органического углерода оставались самыми высокими за всю историю Земли. Изучение позднепротерозойских осадков в других районах мира – в Канаде, Австралии и Южной Африке – подтвердило выводы Нолля. Итак, появлению макроскопической фауны предшествовало резкое увеличение количества свободного кислорода; вряд ли можно счесть это случайным совпадением (рис. 20).
Все это, вероятно, стало следствием событий, произошедших в биосфере около 1,2 млрд лет назад, когда начался постоянный рост численности и видового разнообразия эукариотного фитопланктона (закончился 600 млн лет назад). Известно, что в планктонных экосистемах детритная пищевая цепь развита очень слабо, а оборот вещества и энергии идет почти исключительно по пастбищной цепи; практически вся мертвая органика «проваливается» из экосистемы вниз, на дно водоема, где и утилизуется в тамошних детритных цепях. Однако донные детритные цепи протерозоя существуют в условиях сильного недостатка кислорода и потому обладают ограниченной «пропускной способностью». Начиная с некоторого момента они перестают справляться с тем нарастающим потоком органики, что поставляется сверху наращивающими свою продуктивность фитопланктонными сообществами. Другим источником органики могут быть лишенные в ту пору сколь-нибудь эффективных консументов («травоядных») водорослевые луга. В результате органический углерод начинает захораниваться в осадках на дне океана; оборотной стороной этого процесса является, как мы теперь знаем, накопление свободного кислорода, приводящее – через обратный парниковый эффект – к оледенению.
Малоразмерные фитопланктонные организмы (прежде всего прокариотные) сталкиваются с одной проблемой, на которую впервые обратил внимание гидробиолог Б. Я. Виленкин. Снабжение этих одноклеточных биогенами и растворенными газами происходит за счет диффузии сквозь клеточную стенку. Казалось бы, это должно благоприятствовать миниатюризации клеток, максимальному увеличению отношения ее поверхности к объему. Однако очень мелкий организм неспособен выделиться из окружающей его водной массы. Пассивно паря в толще воды и оставаясь неподвижным относительно нее, он быстро создает вокруг себя «пустыню» – выедает из непосредственно окружающего его водного слоя все биогены, насыщая воду диффундирующей наружу органикой, которую не в силах удерживать внутри клетки из-за того же (высокого) соотношения «поверхность – объем». При этом безвозвратно теряется более 1/3 клеточной продукции. Но хуже то, что на окисление этой органики расходуется кислород и водный слой, окружающий организм, эвтрофицируется. Милый Пятачок прав: жизнь «очень маленького существа» полна неприятностей.
Организм избежит неприятностей, если будет перемещаться относительно вмещающей его воды. Этого он может добиться двумя способами: либо начнет сам активно двигаться в ее толще, либо прикрепится к неподвижному субстрату, чтобы вода двигалась относительно него. Второй способ прокариоты реализовали немедленно, сконструировав мат. А вот с первым способом (казалось бы, более простым и очевидным) у них возникли большие проблемы, ибо эффективных органов движения (подобных жгутикам и ресничкам эукариотных одноклеточных) у прокариот не возникает, а создание клеточных агрегаций, способных к согласованным движениям (например, волнообразным) затруднено из-за крайней слабости межклеточных взаимодействий (см. главу 5). Поэтому «генеральной линией» этот способ становится лишь при появлении эукариот, способных к образованию высокоинтегрированных клеточных агрегаций, т.е. к настоящей многоклеточности.
Многоклеточность привела, среди прочего, к резкому повышению способности организмов создавать в своем теле запас питательных веществ. Практически лишенные этих запасов одноклеточные (в особенности прокариоты) обречены реагировать на изменения содержания биогенов в окружающей среде единственным способом – изменением плотности популяций. (Примером такого их поведения может служить «цветение» водоемов, когда стремительное размножение одноклеточных и нитчатых водорослей быстро приводит к исчерпанию ресурса экосистемы; в результате безмерно разросшаяся популяция в одночасье вымирает, а на окисление этой мертвой органики расходуется затем почти весь кислород водоема – «замор».) Запасные вещества и резервная биомасса крупных организмов делают их популяции более независимыми от колебаний ресурса и стабилизируют их плотность. В свою очередь, существующие в экосистеме геохимические круговороты обретают в лице этих организмов крупное резервное депо[16] , оказывающее на все эти циклы мощное стабилизирующее воздействие. Такая «взаимная стабилизация» кажется одним из главных экологических следствий возникновения многоклеточных (хотя правильнее сказать – макроскопических, т.е. видимых невооруженным глазом) организмов.
Последняя оговорка неслучайна. Если мы от вопроса «для чего возникла многоклеточность?» перейдем к вопросу «как и когда она возникла?», то нам прежде всего придется определить, какой организм следует считать многоклеточным. Если даже не брать в расчет чисто терминологические проблемы (следует ли называть «колонией» Volvox, у которого есть дифференциация между клетками переднего и заднего полушарий, зародыши, возникающие путем палинтомии – деления без последующего увеличения размеров клеток, и который к тому же смертен ), здесь существует и проблема вполне объективная. А именно: соответствует ли строгим критериям многоклеточности строение тела макроскопических водорослей и грибов? Большинство биологов ныне отвечают на этот вопрос отрицательно, оставляя термин «многоклеточные» лишь за зелеными растениями (Metaphyta) и животными (Metazoa); Дж. Корлисс (1983) ввел для этих двух групп удачный термин – многотканевые организмы.
Таким образом, достижение живыми организмами макроскопических размеров может происходить различными способами, а истиная многоклеточность (многотканевость) – лишь один из них. В любом случае, многоклеточность (в широком смысле) почти наверняка возникала в процессе эволюции многократно и независимо: во многих типах водорослей – красных (Rhodophyta), зеленых (Chlorophyta), золотистых (Chrysophyta) – или высших грибов-аскомицетов (Ascomyceta) можно выявить преемственные ряды от одноклеточных форм через колонии (например, нитчатые формы) к макроскопическим организмам с аналогами тканевой дифференцировки.
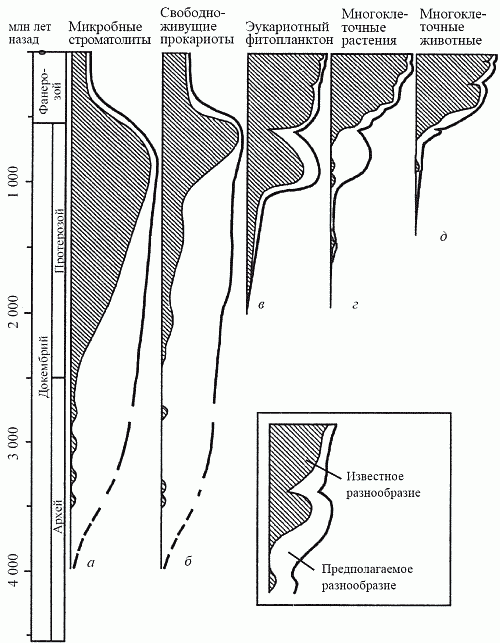
Эта гипотетическая эволюционная картина вполне соответствует современным палеонтологическим даным (рис. 19). Первые эукариоты появились (как мы знаем из главы 5) около 2 млрд лет назад среди фитопланктонных акритарх; вскоре к ним добавились и нитчатые формы с эукариотными параметрами клеток. Замечательно, что они никогда не встречаются в прокариотных бентосных сообществах (цианобактериальных матах), а с самого начала формируют свой собственный тип растительности, названный У. Шенборном (1987) «водорослевыми лугами». Ныне сообщества такого типа известны лишь в некоторых антарктических внутренних водоемах; в докембрии же они, судя по характеру осадков, были широко распространены в морях за пределами мелководий (которые были заняты матами).
В рифее (1,4–1,2 млрд лет назад) эти нитчатые эукариотные формы достигли значительного разнообразия. Именно тогда в составе водорослевых лугов появились и первые макроскопические водоросли с пластинчатым, корковым и кожистым типами слоевища, а к венду (650 млн лет) основным компонентом этих сообществ становятся вендотении – лентовидные водоросли длиной до 15 см. Есть даже сообщение о находке каких-то пластинчатых водорослей в китайской формации Чанчен с возрастом 1,8 млрд лет; эта датировка нуждается в подтверждении, но не кажется нереальной. Дело в том, что таксономическая принадлежность всех этих форм недостаточно ясна, однако некоторые из них очень сходны с низшими красными водорослями – бангиевыми. А красные водоросли, как полагают многие исследователи, являются самой архаичной ветвью эукариот; возможно, они произошли в результате «независимой эукариотизации» цианобактерий.
Рис. 19. Изменения во времени относительного обилия основных биотических компонентов: а – прокариоты-строматолитообразователи; б – свободноживущие прокариоты; в – эукариотный фитопланктон; г – многоклеточные растения; д – многоклеточные животные (по Schopf, 1992)
Животные и следы их жизнедеятельности (норки и следовые дорожки на поверхности осадка) достоверно появились в палеонтологической летописи лишь в конце протерозоя – около 800 млн лет назад. (Интересно, что водоросли с минерализованными слоевищами, для которых можно предполагать тот же уровень организации, что и у высших красных и бурых водорослей, появились еще позднее – в венде.) В чем же причина того, что истиная многоклеточность возникла так поздно? В 60-е годы прошлого века (вспомним главу 5) существовала гипотеза «кислородного контроля» Беркнера и Маршалла, согласно которой содержание кислорода в земной атмосфере вплоть до начала фанерозоя (540 млн лет назад) было ниже точки Пастера и не допускало существования более высоко организованных форм жизни, чем водоросли. Поскольку со временем установили, что точка Пастера в действительности была пройдена гораздо раньше – более чем за миллиард лет до времени появления первых многоклеточных, причинная связь между этими явлениями была отвергнута и о гипотезе «кислородного контроля» забыли. Как позже выяснилось, напрасно.Однопроцентное содержание кислорода (имеется в виду 1% от его современного количества) – это тот критический минимум, ниже которого аэробный метаболизм принципиально невозможен; однако для жизнедеятельности макроскопических животных кислорода необходимо существенно больше. Б. Раннегар недавно провел специальные расчеты, из которых следует, что животным, составлявшим первую фауну многоклеточных – эдиакарскую (о ней речь впереди), кислорода требовалось не менее 6–10% от нынешнего уровня; это в том случае, если они имели развитую систему циркуляции, доставлявшую кислород к тканям. Если же такая система у них еще не развилась и они дышали за счет прямой диффузии (скорее всего, так и было), то требовалось для их жизнедеятельности гораздо больше кислорода (может быть, его содержание сопоставимо с нынешним). Итак, гипотеза «кислородного контроля» кажется вполне логичным объяснением появления макроскопических животных лишь в конце протерозоя – если принять более высокий критический порог, чем однопроцентный, предлагавшийся Беркнером и Маршаллом. Однако возможна ли проверка этой гипотезы? Мы помним (из главы 5), что можно по составу осадков отличить аэробные обстановки от анаэробных, но как определить количественные различия в содержании кислорода в былые эпохи?
Известно, что количество кислорода, создаваемого небиологическими процессами (фотолиз воды и т.д.), совершенно ничтожно; почти весь свободный кислород планеты создан фотосинтезирующими организмами. Однако живые существа не только производят кислород, но и потребляют его при дыхании. В биосфере осуществляется достаточно простая химическая реакция: nСО2+nH2O?(CH2O)n + nО2. «Читая» ее слева направо, мы получаем фотосинтез, а справа налево – дыхание (а также горение и гниение). Уровень содержания кислорода на планете стабилен потому, что прямая и обратная реакции взаимно уравновешиваются; так что если мы попытаемся увеличить содержание свободного кислорода в атмосфере путем простого наращивания объема фотосинтезирующего вещества, то из этой затеи ничего не выйдет. Сместить химическое равновесие, как вам должно быть известно из курса химии, можно, лишь выводя из сферы реакции один из ее продуктов. В нашем случае добиться увеличения выхода О2 можно, лишь необратимо изымая из нее восстановленный углерод в форме (CH2O)n или его производных.
Таким образом, производство кислорода биосферой начинает превалировать над потреблением этого газа (ею же), только если происходит захоронение в осадках неокисленного органического вещества. Этот вывод кажется достаточно парадоксальным и противоречащим расхожим представлениям. Так, например, природоохранная пропаганда вбила в голову широкой публики магическую фразу «Леса – это легкие планеты»; и мало кто дает себе труд задуматься над тем, что в действительности любое сбалансированное сообщество (в том числе тропический лес) потребляет ровно столько кислорода, сколько и производит. Если что и можно назвать «легкими планеты», так это болота, где как раз и идет процесс захоронения неокисленной органики. Поэтому, если мы установим, что в некий период геологической истории происходило интенсивное захоронение органического углерода, то мы вправе будем заключить, что в это время столь же интенсивно накапливался и кислород. А вот оценить темпы захоронения органического углерода в прошлые эпохи вполне возможно. Для этого существует специальный метод, основанный на изучении изотопного отношения 12C/13C в соответствующих осадках (органический углерод, участвовавший когда-либо в реакциях фотосинтеза, обогащен «легким» изотопом 12C).
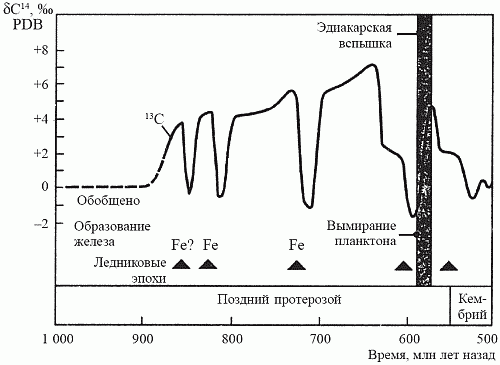
Несколько лет назад на архипелаге Шпицберген была открыта уникальная по полноте последовательность позднепротерозойских осадков, отлагавшихся в период с 850 до 600 млн лет назад. Исследовав эти осадки на предмет изотопного отношения 12C/13C, Э. Нолль (1996) установил, что на протяжении всего этого времени темпы захоронения органического углерода оставались самыми высокими за всю историю Земли. Изучение позднепротерозойских осадков в других районах мира – в Канаде, Австралии и Южной Африке – подтвердило выводы Нолля. Итак, появлению макроскопической фауны предшествовало резкое увеличение количества свободного кислорода; вряд ли можно счесть это случайным совпадением (рис. 20).
Рис. 20. Ключевые характеристики истории позднего протерозоя. Минимумы на кривой захоронения органического углерода соответствуют ледниковым эпохам и интенсивному образованию железорудных формаций (по Ноллю, 1995)
О том, что на Земле в это время действительно существенно возросло содержание кислорода, могут свидетельствовать и другие факты. Во-первых, в океанах вновь происходит осаждение джеспеллитов, прекратившееся около 1,8 млрд лет назад (см. рис. 16). Это может означать, что кислород впервые насытил глубоководные части океана, располагающиеся ниже фотической зоны, и при этом произошло осаждение еще сохранявшихся в тех глубинах запасов закисного железа. Во-вторых, это было время как минимум трех покровных оледенений, последнее из которых – Лапландское оледенение (600 млн лет назад) – было, по всей видимости, самым крупным за всю историю Земли. Одной из причин начала оледенений считают так называемый обратный парниковый эффект: когда в атмосфере планеты отношение О2/СО2 смещается в сторону кислорода, планета начинает гораздо хуже удерживать тепло, доставляемое центральным светилом.Все это, вероятно, стало следствием событий, произошедших в биосфере около 1,2 млрд лет назад, когда начался постоянный рост численности и видового разнообразия эукариотного фитопланктона (закончился 600 млн лет назад). Известно, что в планктонных экосистемах детритная пищевая цепь развита очень слабо, а оборот вещества и энергии идет почти исключительно по пастбищной цепи; практически вся мертвая органика «проваливается» из экосистемы вниз, на дно водоема, где и утилизуется в тамошних детритных цепях. Однако донные детритные цепи протерозоя существуют в условиях сильного недостатка кислорода и потому обладают ограниченной «пропускной способностью». Начиная с некоторого момента они перестают справляться с тем нарастающим потоком органики, что поставляется сверху наращивающими свою продуктивность фитопланктонными сообществами. Другим источником органики могут быть лишенные в ту пору сколь-нибудь эффективных консументов («травоядных») водорослевые луга. В результате органический углерод начинает захораниваться в осадках на дне океана; оборотной стороной этого процесса является, как мы теперь знаем, накопление свободного кислорода, приводящее – через обратный парниковый эффект – к оледенению.