Страница:
вирусы,у которых отсутствуют основные, необходимые для обмена веществ ферменты. Поэтому вирусы могут расти и размножаться, лишь проникая в К. и используя их ферментные системы.
Специальные функции клеток.В процессе эволюции многоклеточных возникло разделение функций между К., что привело к расширению возможностей приспособления животных и растений к меняющимся условиям среды. Закрепившиеся наследственно различия в форме К., их размерах и некоторых сторонах метаболизма реализуются в процессе индивидуального развития организма. Основное проявление развития — дифференцировка К., их структурная и функциональная специализация. Дифференцированные К. имеют такой же набор хромосом, как и оплодотворенная яйцеклетка. Это доказывается пересадкой ядра дифференцированной К. в предварительно лишенную ядра яйцеклетку, после чего может развиваться полноценный организм. Таким образом, различия между дифференцированными К., по-видимому, обусловливаются разными соотношениями активных и неактивных генов, каждый из которых кодирует биосинтез определённого белка. Судя по составу белков, в дифференцированных К. активна (способна к транскрипции) лишь небольшая часть (порядка 10%) генов, свойственных К. данного вида организмов. Среди них лишь немногие ответственны за специальную функцию К., а остальные обеспечивают общеклеточные функции. Так, в мышечных К. активны гены, кодирующие структуру сократимых белков, в эритроидных К. — гены, кодирующие биосинтез гемоглобина, и т.д. Однако в каждой К. должны быть активны гены, определяющие биосинтез веществ и структур, необходимых для всех К., например ферментов, участвующих в энергетических превращениях веществ. В процессе специализации К. отдельные общеклеточные функции их могут развиваться особенно сильно. Так, в железистых К. более всего выражена синтетическая активность, мышечные — наиболее сократимы, нервные — наиболее возбудимы. В узкоспециализированных К. обнаруживаются структуры, характерные лишь для этих К. (например, у животных — миофибриллы мышц, тонофибриллы и реснички некоторых покровных К., нейрофибриллы нервных К., жгутики у простейших или у сперматозоидов многоклеточных организмов). Иногда специализация сопровождается утратой некоторых свойств (например, нервные К. утрачивают способность к размножению; ядра К. кишечного эпителия млекопитающих не могут в зрелом состоянии синтезировать РНК; зрелые эритроциты млекопитающих лишены ядра). Выполнение важных для организма функций включает иногда гибель К. Так, К. эпидермиса кожи постепенно ороговевают и гибнут, но остаются некоторое время в пласте, предохраняя подлежащие ткани от повреждения и инфекции. В сальных железах К. постепенно превращаются в капли жира, который используется организмом или выделяется. Для выполнения некоторых тканевых функций К. образуют неклеточные структуры. Основные пути их образования — секреция или превращения компонентов цитоплазмы. Так, значительная по объёму часть подкожной клетчатки, хряща и кости составляет межуточное вещество — производное К. соединительной ткани. К. крови обитают в жидкой среде (плазме крови), содержащей белки, сахара и др. вещества, вырабатываемые разными К. организма. К. эпителия, образующие пласт, окружены тонкой прослойкой диффузно распределённых веществ, главным образом гликопротеидов (так называемый цемент, или надмембранный компонент). Внешние покровы членистоногих и раковины моллюсков — также продукты выделения К. Взаимодействие специализированных К. — необходимое условие жизни организма и нередко самих этих К. (см. Гистология ) .Лишённые связей друг с другом, например в культуре, К. быстро утрачивают особенности присущих им специальных функций.
Деление клеток. В основе способности К. к самовоспроизведению лежат уникальное свойство ДНК самокопироваться и строго равноценное деление репродуцированных хромосом в процессе митоза.В результате деления образуются две К., идентичные исходной по генетическим свойствам и с обновленным составом ядра и цитоплазмы. Процессы самовоспроизведения хромосом, их деления, образования двух ядер и деления цитоплазмы разделены во времени, составляя в совокупности митотический цикл К. В случае, если после деления К. начинает готовиться к следующему делению, митотический цикл совпадает с жизненным циклом К. Однако во многих случаях после деления (а иногда перед ним) К. выходят из митотического цикла, дифференцируются и выполняют в организме ту или иную специальную функцию. Состав таких К. может обновляться за счёт делений малодифференцированных К. В некоторых тканях и дифференцированные К. способны повторно входить в митотический цикл. В нервной ткани дифференцированные К. не делятся; многие из них живут так же долго, как организм в целом, то есть у человека — несколько десятков лет. При этом ядра нервных К. не утрачивают способности к делению: будучи пересажены в цитоплазму раковых К., ядра нейронов синтезируют ДНК и делятся. Опыты с клетками-гибридами показывают влияние цитоплазмы на проявление ядерных функций. Неполноценная подготовка к делению предотвращает митоз или искажает его течение. Так, в некоторых случаях не происходит деления цитоплазмы и образуется двуядерная К. Многократное деление ядер в неделящейся К. приводит к появлению многоядерных К. или сложных надклеточных структур (симпластов), например в поперечнополосатых мышцах. Иногда репродукция К. ограничивается воспроизведением хромосом, и образуется полиплоидная К., имеющая удвоенный (сравнительно с исходной К.) набор хромосом. Полиплоидизация приводит к усилению синтетической активности, увеличению размеров и массы К.
Обновление клеток.Для длительной работы каждой К. необходимо восстановление изнашиваемых структур, как и ликвидация повреждений К., вызванных внешними воздействиями. Восстановительные процессы, характерные для всех К., связаны с изменениями проницаемости плазматической мембраны и сопровождаются усилением внутриклеточных синтезов, в первую очередь синтеза белка. Во многих тканях стимуляция восстановительных процессов приводит к репродукции генетического аппарата и делению К.; это свойственно, например покровам или кроветворной системе. Процессы внутриклеточного обновления в этих тканях выражены слабо, их К. живут сравнительно недолго (например, К. кишечного покрова млекопитающих — всего несколько суток). Максимальной выраженности внутриклеточные восстановительные процессы достигают в неделящихся или слабоделящихся клеточных популяциях, например в нервных К. Показателем совершенства процессов внутреннего обновления К. является длительность их жизни; для многих нервных К. она совпадает с продолжительностью жизни всего организма.
Мутации.Обычно процесс воспроизведения ДНК происходит без отклонений, и генетический код остаётся постоянным, что обеспечивает синтез одного и того же набора белков в огромном числе клеточных поколений. Однако в редких случаях может произойти мутация—частичное изменение структуры гена. Конечный её эффект — изменение свойств белков, кодируемых мутантными генами. Если при этом затрагиваются важные ферментные системы, свойства К., а иногда и всего организма существенно изменяются. Так, мутация одного из генов, контролирующих синтез гемоглобина, приводит к тяжелому заболеванию — анемии.Естественный отбор полезных мутаций — важный механизм эволюции.
Регуляция функций клеток.Основной механизм регуляции внутриклеточных процессов связан с различными влияниями на ферменты — высоко специфичные катализаторы биохимических реакций. Регуляция может осуществляться на генетическом уровне, когда определяется состав ферментов или количество того или иного фермента в К. В последнем случае регуляция может происходить и на уровне трансляции. Другой тип регуляции — воздействие на сам фермент, в результате чего может происходить как торможение, так и стимуляция его активности. Структурный уровень регуляции — влияние на сборку клеточных структур: мембран, рибосом и т.д. Конкретными регуляторами внутриклеточных процессов могут быть нервные влияния, гормоны, специальные вещества, вырабатываемые внутри К. либо окружающими К. (особенно белки), или же сами продукты реакций. В последнем случае воздействие осуществляется по принципу обратной связи, когда продукт реакции влияет на активность фермента — катализатора этой реакции. Регуляция может осуществляться через транспорт предшественников и ионов, влияния на матричный синтез (РНК, полисомы, ферменты синтеза), изменение формы регулируемого фермента.
Организация и регуляция функций К. на молекулярном уровне определяют такие свойства живых систем, как пространственная компактность и энергетическая экономичность. Важное свойство многоклеточных организмов — надёжность — во многом зависит от множественности (взаимозаменяемости) К. каждого функционального типа, а также от возможности их замены в результате размножения К. и обновления компонентов каждой К.
В медицине используются воздействия на К. для лечения и предупреждения заболеваний. Многие лекарственные вещества изменяют активность определенных К. Так, наркотики, транквилизаторы и болеутоляющие вещества снижают интенсивность деятельности нервных К., а стимуляторы её усиливают. Некоторые вещества стимулируют сокращение мышечных К. сосудов, другие — матки или сердца. Специальные воздействия на делящиеся К. осуществляются при использовании радиации или цитостатических веществ, блокирующих деление К. Иммунизация стимулирует деятельность лимфоидных К., вырабатывающих антитела к чужеродным белкам, предупреждая тем самым многие заболевания.
Лит.:Кольцов Н. К., Организация клетки, М. — Л., 1936; Вильсон Э., Клетка и её роль в развитии и наследственности, пер. с англ., т. 1—2, М. — Л., 1936—1940; Насонов Д. Н. и Александров В. Я., Реакция живого вещества на внешние воздействия, М. — Л., 1940; Кедровский Б. В., Цитология белковых синтезов в животной клетке, М., 1959; Мэзия Д., Митоз и физиология клеточного деления, пер. с англ., М., 1963; Руководство по цитологии, т. 1—2, М. — Л., 1965—66; Бродский В. Я., Трофика клетки, М., 1966; Живая клетка, [Сб. ст.], пер. с англ., М., 1966; Де Робертис Э., Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1967; Васильев Ю. М. и Маленков А. Г., Клеточная поверхность и реакции клеток, Л., 1968; Алов И. А., Брауде А. И., Аспиз М. Е., Основы функциональной морфологии клетки, 2 изд., М., 1969; Лёви А., Сикевиц Ф., Структура и функции клетки, пер. с англ., М., 1971; Handbook of molecular cytology, ed. A. Lima-de-Faria, Amst., 1969.
В. Я. Бродский.
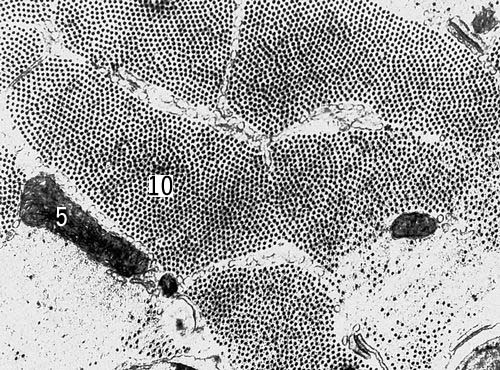 Поперечный срез мышечных клеток саранчи (увеличено в 54000 раз). Условные обозначения: 5 — митохондрии, 10 — миофибриллы.
Поперечный срез мышечных клеток саранчи (увеличено в 54000 раз). Условные обозначения: 5 — митохондрии, 10 — миофибриллы.
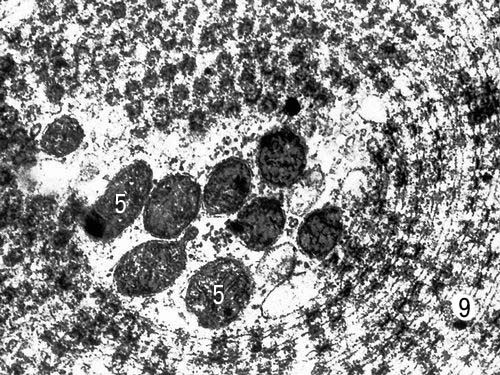 Специализированная форма мембран (пористые пластинки) в цитоплазме созревающей яйцеклетки севрюги (увеличено в 35000 раз). Условные обозначения: 5 — митохондрии, 9 — пористые пластинки.
Специализированная форма мембран (пористые пластинки) в цитоплазме созревающей яйцеклетки севрюги (увеличено в 35000 раз). Условные обозначения: 5 — митохондрии, 9 — пористые пластинки.
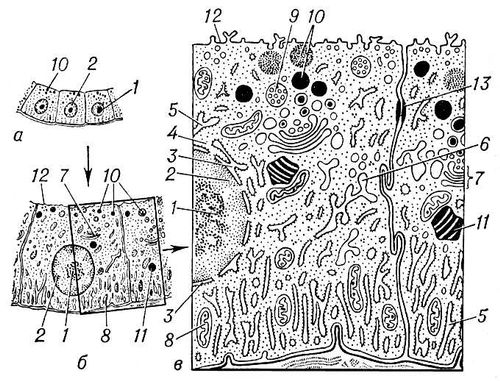 Рис. 1. Общий вид эпителиальной клетки животного при различном увеличении: а — в оптический микроскоп; б — при малом увеличении электронного микроскопа; в — при большом увеличении. Структуры ядра: 1 — ядрышко; 2 — хроматин (участки хромосом); 3 — ядерная оболочка. Структуры цитоплазмы: 4 — рибосомы; 5 — гранулярная (покрытая рибосомами) эндоплазматическая сеть; 6 — гладкоконтурная сеть; 7 — комплекс Гольджи; 8 — митохондрии; 9 — мультивезикулярные (многопузырьковые) тела; 10 — секреторные гранулы; 11 — жировые включения; 12 — плазматическая мембрана; 13 — десмосома.
Рис. 1. Общий вид эпителиальной клетки животного при различном увеличении: а — в оптический микроскоп; б — при малом увеличении электронного микроскопа; в — при большом увеличении. Структуры ядра: 1 — ядрышко; 2 — хроматин (участки хромосом); 3 — ядерная оболочка. Структуры цитоплазмы: 4 — рибосомы; 5 — гранулярная (покрытая рибосомами) эндоплазматическая сеть; 6 — гладкоконтурная сеть; 7 — комплекс Гольджи; 8 — митохондрии; 9 — мультивезикулярные (многопузырьковые) тела; 10 — секреторные гранулы; 11 — жировые включения; 12 — плазматическая мембрана; 13 — десмосома.
 Рис. 2. Схема строения клетки образовательной ткани (меристемы) растения: 1 — клеточная стенка; 2 — плазмодесмы; 3 — плазматическая мембрана; 4 — эндоплазматическая сеть; 5 — вакуоли; 6 — рибосомы; 7 — митохондрии; 8 — пластида; 9 — комплекс Гольджи; 10 — оболочка ядра; 11 — поры в ядерной оболочке; 12 — хроматин; 13 — ядрышко.
Рис. 2. Схема строения клетки образовательной ткани (меристемы) растения: 1 — клеточная стенка; 2 — плазмодесмы; 3 — плазматическая мембрана; 4 — эндоплазматическая сеть; 5 — вакуоли; 6 — рибосомы; 7 — митохондрии; 8 — пластида; 9 — комплекс Гольджи; 10 — оболочка ядра; 11 — поры в ядерной оболочке; 12 — хроматин; 13 — ядрышко.
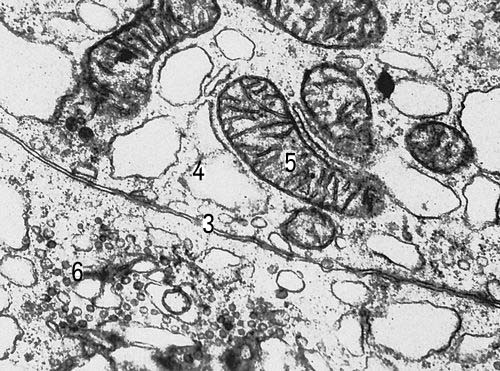 Участки двух клеток щитовидной железы крысы (увеличено в 30000 раз). Условные обозначения: 3 — клеточная оболочка, 4 — эндоплазматическая сеть, 5 — митохондрии, 6 — комплекс Гольджи.
Участки двух клеток щитовидной железы крысы (увеличено в 30000 раз). Условные обозначения: 3 — клеточная оболочка, 4 — эндоплазматическая сеть, 5 — митохондрии, 6 — комплекс Гольджи.
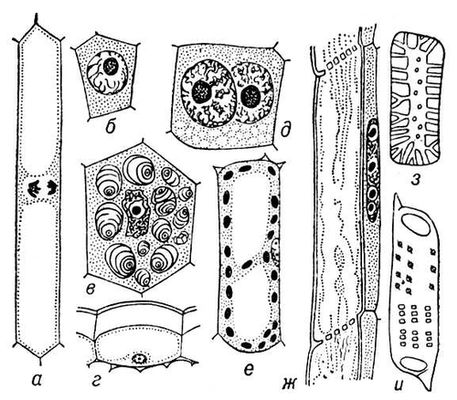 Рис. 3. Разнообразие клеток высших растений: а, б — меристематические клетки; в — крахмалоносная клетка из запасающей паренхимы; г — клетка эпидермиса; д — двуядерная клетка секреторного слоя пыльцевого гнезда; е — клетка ассимиляционной ткани листа с хлоропластами; ж — членик ситовидной трубки с клеткой-спутницей; з — каменистая клетка; и — членик сосуда.
Рис. 3. Разнообразие клеток высших растений: а, б — меристематические клетки; в — крахмалоносная клетка из запасающей паренхимы; г — клетка эпидермиса; д — двуядерная клетка секреторного слоя пыльцевого гнезда; е — клетка ассимиляционной ткани листа с хлоропластами; ж — членик ситовидной трубки с клеткой-спутницей; з — каменистая клетка; и — членик сосуда.
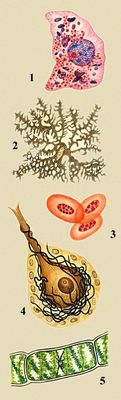 Разнообразие животных и растительных клеток: 1 — клетка печени аксолотля, в цитоплазме — красные митохондрии и фиолетовые белковые включения, в ядре — красное ядрышко и синие глыбки хроматина; 2 — хроматофор аксолотля, заполненный гранулами пигмента; 3 — эритроциты лягушки; 4 — клетка Пуркине мозжечка крысы; 5 — клетка водоросли спирогиры.
Разнообразие животных и растительных клеток: 1 — клетка печени аксолотля, в цитоплазме — красные митохондрии и фиолетовые белковые включения, в ядре — красное ядрышко и синие глыбки хроматина; 2 — хроматофор аксолотля, заполненный гранулами пигмента; 3 — эритроциты лягушки; 4 — клетка Пуркине мозжечка крысы; 5 — клетка водоросли спирогиры.
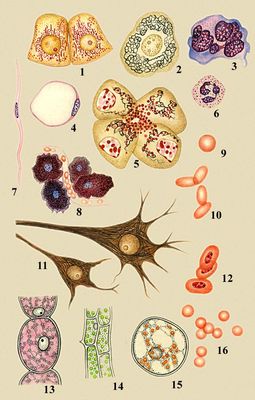 Разнообразие животных и растительных клеток: 1 — клетки почки лягушки, видны митохондрии; 2 — чувствительная клетка спинномозгового ганглия человека, виден комплекс Гольджи; 3 — мегакариоцит из костного мозга человека; 4 — жировая клетка из подкожной клетчатки крысы; 5 — клетки поджелудочной железы человека, видны комплекс Гольджи и секреторные гранулы; 6 — нейтрофильный лейкоцит человека; 7 — гладкая мышечная клетка кишечника человека; 8 — тучные клетки в рыхлой соединительной ткани крысы; 9 — эритроциты человека; 10 — эритроциты верблюда; 11 — малая и большая пирамидальные клетки коры головного мозга человека; 12 — эритроциты курицы; 13 — клетка волоска тычиночной нити традесканции; 14 — клетки листа элодеи; 15 — клетка плода ландыша; 16 — эритроциты свиньи.
Разнообразие животных и растительных клеток: 1 — клетки почки лягушки, видны митохондрии; 2 — чувствительная клетка спинномозгового ганглия человека, виден комплекс Гольджи; 3 — мегакариоцит из костного мозга человека; 4 — жировая клетка из подкожной клетчатки крысы; 5 — клетки поджелудочной железы человека, видны комплекс Гольджи и секреторные гранулы; 6 — нейтрофильный лейкоцит человека; 7 — гладкая мышечная клетка кишечника человека; 8 — тучные клетки в рыхлой соединительной ткани крысы; 9 — эритроциты человека; 10 — эритроциты верблюда; 11 — малая и большая пирамидальные клетки коры головного мозга человека; 12 — эритроциты курицы; 13 — клетка волоска тычиночной нити традесканции; 14 — клетки листа элодеи; 15 — клетка плода ландыша; 16 — эритроциты свиньи.
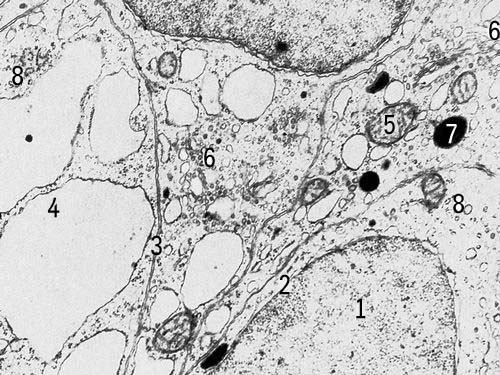 Клетки щитовидной железы крысы с включениями (увеличено в 18000 раз). Условные обозначения: 1 — ядро, 2 — ядерная оболочка, 3 — клеточная оболочка, 4 — эндоплазматическая сеть, 5 — митохондрии, 6 — комплекс Гольджи, 7 — плотные тела, 8 — рибосомы.
Клетки щитовидной железы крысы с включениями (увеличено в 18000 раз). Условные обозначения: 1 — ядро, 2 — ядерная оболочка, 3 — клеточная оболочка, 4 — эндоплазматическая сеть, 5 — митохондрии, 6 — комплекс Гольджи, 7 — плотные тела, 8 — рибосомы.
клетка.
К. т. утверждает представление о единстве всего живого и его эволюционном развитии. Ф. Энгельс назвал К. т. одним из трёх величайших открытий, обеспечивших прогресс естествознания в 19 в. (см. «Диалектика природы», 1969, с. 168).
Специальные функции клеток.В процессе эволюции многоклеточных возникло разделение функций между К., что привело к расширению возможностей приспособления животных и растений к меняющимся условиям среды. Закрепившиеся наследственно различия в форме К., их размерах и некоторых сторонах метаболизма реализуются в процессе индивидуального развития организма. Основное проявление развития — дифференцировка К., их структурная и функциональная специализация. Дифференцированные К. имеют такой же набор хромосом, как и оплодотворенная яйцеклетка. Это доказывается пересадкой ядра дифференцированной К. в предварительно лишенную ядра яйцеклетку, после чего может развиваться полноценный организм. Таким образом, различия между дифференцированными К., по-видимому, обусловливаются разными соотношениями активных и неактивных генов, каждый из которых кодирует биосинтез определённого белка. Судя по составу белков, в дифференцированных К. активна (способна к транскрипции) лишь небольшая часть (порядка 10%) генов, свойственных К. данного вида организмов. Среди них лишь немногие ответственны за специальную функцию К., а остальные обеспечивают общеклеточные функции. Так, в мышечных К. активны гены, кодирующие структуру сократимых белков, в эритроидных К. — гены, кодирующие биосинтез гемоглобина, и т.д. Однако в каждой К. должны быть активны гены, определяющие биосинтез веществ и структур, необходимых для всех К., например ферментов, участвующих в энергетических превращениях веществ. В процессе специализации К. отдельные общеклеточные функции их могут развиваться особенно сильно. Так, в железистых К. более всего выражена синтетическая активность, мышечные — наиболее сократимы, нервные — наиболее возбудимы. В узкоспециализированных К. обнаруживаются структуры, характерные лишь для этих К. (например, у животных — миофибриллы мышц, тонофибриллы и реснички некоторых покровных К., нейрофибриллы нервных К., жгутики у простейших или у сперматозоидов многоклеточных организмов). Иногда специализация сопровождается утратой некоторых свойств (например, нервные К. утрачивают способность к размножению; ядра К. кишечного эпителия млекопитающих не могут в зрелом состоянии синтезировать РНК; зрелые эритроциты млекопитающих лишены ядра). Выполнение важных для организма функций включает иногда гибель К. Так, К. эпидермиса кожи постепенно ороговевают и гибнут, но остаются некоторое время в пласте, предохраняя подлежащие ткани от повреждения и инфекции. В сальных железах К. постепенно превращаются в капли жира, который используется организмом или выделяется. Для выполнения некоторых тканевых функций К. образуют неклеточные структуры. Основные пути их образования — секреция или превращения компонентов цитоплазмы. Так, значительная по объёму часть подкожной клетчатки, хряща и кости составляет межуточное вещество — производное К. соединительной ткани. К. крови обитают в жидкой среде (плазме крови), содержащей белки, сахара и др. вещества, вырабатываемые разными К. организма. К. эпителия, образующие пласт, окружены тонкой прослойкой диффузно распределённых веществ, главным образом гликопротеидов (так называемый цемент, или надмембранный компонент). Внешние покровы членистоногих и раковины моллюсков — также продукты выделения К. Взаимодействие специализированных К. — необходимое условие жизни организма и нередко самих этих К. (см. Гистология ) .Лишённые связей друг с другом, например в культуре, К. быстро утрачивают особенности присущих им специальных функций.
Деление клеток. В основе способности К. к самовоспроизведению лежат уникальное свойство ДНК самокопироваться и строго равноценное деление репродуцированных хромосом в процессе митоза.В результате деления образуются две К., идентичные исходной по генетическим свойствам и с обновленным составом ядра и цитоплазмы. Процессы самовоспроизведения хромосом, их деления, образования двух ядер и деления цитоплазмы разделены во времени, составляя в совокупности митотический цикл К. В случае, если после деления К. начинает готовиться к следующему делению, митотический цикл совпадает с жизненным циклом К. Однако во многих случаях после деления (а иногда перед ним) К. выходят из митотического цикла, дифференцируются и выполняют в организме ту или иную специальную функцию. Состав таких К. может обновляться за счёт делений малодифференцированных К. В некоторых тканях и дифференцированные К. способны повторно входить в митотический цикл. В нервной ткани дифференцированные К. не делятся; многие из них живут так же долго, как организм в целом, то есть у человека — несколько десятков лет. При этом ядра нервных К. не утрачивают способности к делению: будучи пересажены в цитоплазму раковых К., ядра нейронов синтезируют ДНК и делятся. Опыты с клетками-гибридами показывают влияние цитоплазмы на проявление ядерных функций. Неполноценная подготовка к делению предотвращает митоз или искажает его течение. Так, в некоторых случаях не происходит деления цитоплазмы и образуется двуядерная К. Многократное деление ядер в неделящейся К. приводит к появлению многоядерных К. или сложных надклеточных структур (симпластов), например в поперечнополосатых мышцах. Иногда репродукция К. ограничивается воспроизведением хромосом, и образуется полиплоидная К., имеющая удвоенный (сравнительно с исходной К.) набор хромосом. Полиплоидизация приводит к усилению синтетической активности, увеличению размеров и массы К.
Обновление клеток.Для длительной работы каждой К. необходимо восстановление изнашиваемых структур, как и ликвидация повреждений К., вызванных внешними воздействиями. Восстановительные процессы, характерные для всех К., связаны с изменениями проницаемости плазматической мембраны и сопровождаются усилением внутриклеточных синтезов, в первую очередь синтеза белка. Во многих тканях стимуляция восстановительных процессов приводит к репродукции генетического аппарата и делению К.; это свойственно, например покровам или кроветворной системе. Процессы внутриклеточного обновления в этих тканях выражены слабо, их К. живут сравнительно недолго (например, К. кишечного покрова млекопитающих — всего несколько суток). Максимальной выраженности внутриклеточные восстановительные процессы достигают в неделящихся или слабоделящихся клеточных популяциях, например в нервных К. Показателем совершенства процессов внутреннего обновления К. является длительность их жизни; для многих нервных К. она совпадает с продолжительностью жизни всего организма.
Мутации.Обычно процесс воспроизведения ДНК происходит без отклонений, и генетический код остаётся постоянным, что обеспечивает синтез одного и того же набора белков в огромном числе клеточных поколений. Однако в редких случаях может произойти мутация—частичное изменение структуры гена. Конечный её эффект — изменение свойств белков, кодируемых мутантными генами. Если при этом затрагиваются важные ферментные системы, свойства К., а иногда и всего организма существенно изменяются. Так, мутация одного из генов, контролирующих синтез гемоглобина, приводит к тяжелому заболеванию — анемии.Естественный отбор полезных мутаций — важный механизм эволюции.
Регуляция функций клеток.Основной механизм регуляции внутриклеточных процессов связан с различными влияниями на ферменты — высоко специфичные катализаторы биохимических реакций. Регуляция может осуществляться на генетическом уровне, когда определяется состав ферментов или количество того или иного фермента в К. В последнем случае регуляция может происходить и на уровне трансляции. Другой тип регуляции — воздействие на сам фермент, в результате чего может происходить как торможение, так и стимуляция его активности. Структурный уровень регуляции — влияние на сборку клеточных структур: мембран, рибосом и т.д. Конкретными регуляторами внутриклеточных процессов могут быть нервные влияния, гормоны, специальные вещества, вырабатываемые внутри К. либо окружающими К. (особенно белки), или же сами продукты реакций. В последнем случае воздействие осуществляется по принципу обратной связи, когда продукт реакции влияет на активность фермента — катализатора этой реакции. Регуляция может осуществляться через транспорт предшественников и ионов, влияния на матричный синтез (РНК, полисомы, ферменты синтеза), изменение формы регулируемого фермента.
Организация и регуляция функций К. на молекулярном уровне определяют такие свойства живых систем, как пространственная компактность и энергетическая экономичность. Важное свойство многоклеточных организмов — надёжность — во многом зависит от множественности (взаимозаменяемости) К. каждого функционального типа, а также от возможности их замены в результате размножения К. и обновления компонентов каждой К.
В медицине используются воздействия на К. для лечения и предупреждения заболеваний. Многие лекарственные вещества изменяют активность определенных К. Так, наркотики, транквилизаторы и болеутоляющие вещества снижают интенсивность деятельности нервных К., а стимуляторы её усиливают. Некоторые вещества стимулируют сокращение мышечных К. сосудов, другие — матки или сердца. Специальные воздействия на делящиеся К. осуществляются при использовании радиации или цитостатических веществ, блокирующих деление К. Иммунизация стимулирует деятельность лимфоидных К., вырабатывающих антитела к чужеродным белкам, предупреждая тем самым многие заболевания.
Лит.:Кольцов Н. К., Организация клетки, М. — Л., 1936; Вильсон Э., Клетка и её роль в развитии и наследственности, пер. с англ., т. 1—2, М. — Л., 1936—1940; Насонов Д. Н. и Александров В. Я., Реакция живого вещества на внешние воздействия, М. — Л., 1940; Кедровский Б. В., Цитология белковых синтезов в животной клетке, М., 1959; Мэзия Д., Митоз и физиология клеточного деления, пер. с англ., М., 1963; Руководство по цитологии, т. 1—2, М. — Л., 1965—66; Бродский В. Я., Трофика клетки, М., 1966; Живая клетка, [Сб. ст.], пер. с англ., М., 1966; Де Робертис Э., Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1967; Васильев Ю. М. и Маленков А. Г., Клеточная поверхность и реакции клеток, Л., 1968; Алов И. А., Брауде А. И., Аспиз М. Е., Основы функциональной морфологии клетки, 2 изд., М., 1969; Лёви А., Сикевиц Ф., Структура и функции клетки, пер. с англ., М., 1971; Handbook of molecular cytology, ed. A. Lima-de-Faria, Amst., 1969.
В. Я. Бродский.
клетка.
К. т. утверждает представление о единстве всего живого и его эволюционном развитии. Ф. Энгельс назвал К. т. одним из трёх величайших открытий, обеспечивших прогресс естествознания в 19 в. (см. «Диалектика природы», 1969, с. 168).
Исторически открытие клеток и создание К. т. не совпадают. Впервые наблюдал под микроскопом клеточное строение у растений на срезах пробки и стеблей различных живых растений английский микроскопист Р. Гук, описавший свои наблюдения в сочинении «Микрография» (1665). Английский ботаник Н. Грю полагал, что стенки клеток образованы переплётом волокон, наподобие текстиля, откуда и возник термин «ткани» (1682). В 18 в. под влиянием философских идей в науку начинает проникать мысль о единстве живой природы. Попытку найти нечто общее в строении растений и животных сделал К. Ф.
Вольф,но его представления об общности процессов развития «пузырьков», «зёрнышек» и «клеток» были лишь провозвестниками будущей К. т., как и идеи немецкого учёного Л. Окена о построении организмов из «пузырьков» или «инфузорий». В начале 19 в., в связи с успехами в микроскопическом изучении растений, стало ясно, что клетки — не пустоты в общей массе раститительного, вещества, а структуры, имеющие собственную оболочку; их можно изолировать друг от друга. К концу 3-го десятилетия 19 в. выяснилось, что почти все органы растений имеют клеточное строение, и в учебнике немецкого ботаника Ф. Мейена (1830) клетка уже фигурирует как общий структурный элемент тканей растений. Но клетку ещё понимали как камеру, главная часть которой составляет её оболочка, а содержимое имеет второстепенное значение. Ядро в растительной клетке описал Р.
Броун
(1831), но внимание к ядру привлек М.
Шлейден,считавший его цитобластом — образователем клетки. По Шлейдену, из зернистой субстанции конденсируется ядрышко, вокруг которого формируется ядро, а вокруг ядра — клетка, причём ядро в процессе образования клетки исчезает. В начале 2-й четверти 19 в. работы школы чешского биолога Я. Пуркине дали большой материал по микроскопическому строению тканей животных, но в своей «теории зернышек» Пуркине не смог провести границу между различными «зернышками» (так он называл клетки, ядра, а иногда и секреторные включения).
Заслуга оформления К. т. принадлежит ученику немецкого биолога И. Мюллера — Т.
Шванну,который, ознакомившись с исследованиями Шлейдена, увидел в ядре критерий для сопоставления тканевых структур животных и клеток растений. В 1839 вышло сочинение Шванна «Микроскопические исследования о соответствии в структуре и росте животных и растений» (рус. пер. 1939), в самом заглавии которого
была выражена сущность К. т. Однако Шванн продолжал считать главным компонентом клетки ее оболочку и воспринял ложное представление Шлейдена о новообразовании клеток из бесструктурного вещества (цитобластемы). Затем К. т. была распространена на одноклеточные организмы —
простейших,которые были признаны свободно живущими клетками (К. Зибольд, 1848).
Дальнейшее развитие К. т. связано с открытием протоплазмы и клеточного деления. К середине 19 в. выяснилось, что главным в клетке является её «содержимое» — протоплазма. В 1858 немецкий патолог Р.
Вирхов
опубликовал «Целлюлярную патологию», в которой распространил К. т. на явления патологии и обратил внимание на ведущее значение ядра в клетке, провозгласив принцип образования клеток путём деления («каждая клетка из клетки»). Деление вначале трактовалось как перешнуровка ядра и клеточного тела. В 70—80-х гг. был открыт
митоз
как универсальный способ клеточного деления, типичный для всех клеточных организмов. В конце 19 в. были открыты клеточные органоиды, и клетку перестали рассматривать как простой комочек протоплазмы. Вместе с тем во 2-й половине 19 в. наметилась механистическая трактовка организма как суммы клеток.
Современная К. т. исходит из единства расчленённости многоклеточных организмов на клетки и целостности организма, основанной на взаимодействии клеток. Чем сложнее организм, тем более выступает его целостность, которая у животных осуществляется нервной и гуморальной системами, а у растений — непосредственной цитоплазматической связью клеток (плазмодесмами и фитогормонами). Электронномикроскопические исследования укрепили основные положения К. т. Доказана универсальность клеточных органоидов в растительных и животных клетках. Показано, что есть организмы (Procariota), не имеющие оформленного ядра (например, бактериофаги, вирусы, отчасти бактерии, синезелёные водоросли); некоторые из них (бактерии, водоросли) часто называют клетками, исходя из наличия у них ДНК, но правильнее оставить понятие клетки за организмами, у которых ДНК оформлена в виде хромосом и находится в ядрах (Eucariota).
Лит.:Кацнельсон З. С., Клеточная теория в её историческом развитии, Л., 1963; Вермель Е. М., История учения о клетке, М., 1970.
З. С. Кацнельсон.
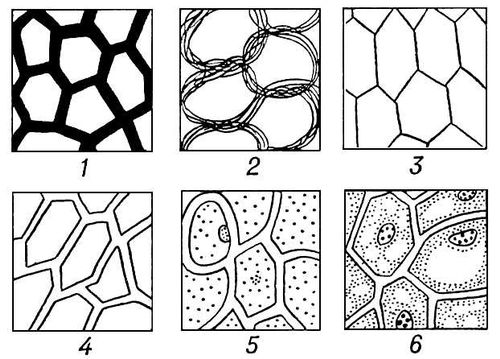 Развитие представлений о клеточном строении растений: 1 — клетки-пустоты в непрерывном растительном веществе (Р. Гук, 1665): 2 — стенки клеток или пузырьков построены из переплетённых волокон, образующих ткань (Н. Грю, 1682); 3 — клетки-камеры, имеющие общую стенку (начало 19 в.); 4 — каждая клетка имеет собственную оболочку (Г. Линк, И. Мольденхавер, 1812); 5 — образователь клетки — ядро («цитобласт»), исчезающее в процессе клеткообразования (М. Шлейден, 1838): 6 — клетки, состоящие из протоплазмы и ядра (Х. Моль, 1844).
Развитие представлений о клеточном строении растений: 1 — клетки-пустоты в непрерывном растительном веществе (Р. Гук, 1665): 2 — стенки клеток или пузырьков построены из переплетённых волокон, образующих ткань (Н. Грю, 1682); 3 — клетки-камеры, имеющие общую стенку (начало 19 в.); 4 — каждая клетка имеет собственную оболочку (Г. Линк, И. Мольденхавер, 1812); 5 — образователь клетки — ядро («цитобласт»), исчезающее в процессе клеткообразования (М. Шлейден, 1838): 6 — клетки, состоящие из протоплазмы и ядра (Х. Моль, 1844).