И тем не менее в последнее время появились данные, свидетельствующие о том, что в геноме дрозофилы, например, синтез РНК может идти на обеих цепях ДНК. Это так называемый симметричный синтез. Любопытно, что он идет от одной точки в разные стороны: в каждой цепи от З’ к 5’-концу, так что обе образующиеся РНК начинаются с 5’-конца. Биологический смысл этого явления мы еще не знаем.
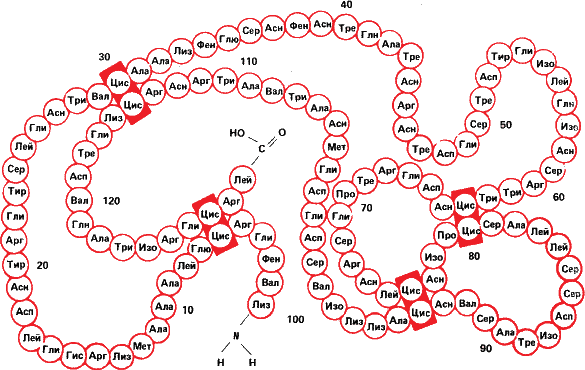
 Рис. 18. Вверху – первичная структура белка лизоцима, разрушающего оболочки бактерий. Обратите внимание на четыре сшивки цис – цис (дисульфидные связи, которые мы уже видели на рис. 3). Внизу же не абстрактная скульптура, а модель третичной скульптуры лизоцима, полученная методом рассеивания рентгеновских лучей.
Рис. 18. Вверху – первичная структура белка лизоцима, разрушающего оболочки бактерий. Обратите внимание на четыре сшивки цис – цис (дисульфидные связи, которые мы уже видели на рис. 3). Внизу же не абстрактная скульптура, а модель третичной скульптуры лизоцима, полученная методом рассеивания рентгеновских лучей.
Перекрывается ли код? Первые гипотетические модели кода были перекрывающимися Это значило, что последовательность нуклеотидов могла кодировать разные аминокислотные последовательности, в зависимости от начала считывания. Так, последовательность АТТГЦАТЦГ, если считывалась бы с А, кодировала бы Тир—Вал—Ала, если со второго нуклеотида (Т)—Вал—Ала—Мет, и т. д. Подобный код накладывал бы жесткие ограничения на последовательность аминокислот в белках. И все облегченно вздохнули, когда С. Бреннер доказал, что каждый триплет нуклеотидов в ДНК и РНК считывается только один раз (неперекрывающийся код).
И опять как гром с ясного неба. Оказалось, что у некоторых фагов гены перекрываются. У фага φX174 имеется двойное перекрытие, у фага G4 даже тройное, то есть с одной нуклеотидной последовательности считываются три аминокислотных! Это предел экономичности сигнала. Фагам хорошо, а вот каково молекулярным генетикам? Как широко распространен этот феномен в мире вирусов? Встречается ли он у высших организмов? Не знаем.
Но самое интересное из новых открытий я приберег под конец.
Конец коллинеарности гена. До последних лет все согласно считали, что ген коллинеарен тому белку, точнее, той полипептидной цепи, которую он кодирует. Иными словами, каждой тройке нуклеотидов в ДНК, с которой считывается мРНК, соответствует один аминокислотиый остаток в полипептиде.
И опять оказалось не так! В смысловой цепи ДНК, кодирующей белок, обнаружены довольно длинные вставки (интроны), никаких аминокислот не кодирующие. Они считываются при синтезе первичного транскрипта, а далее начинается непонятный процесс. Ненужные вставки вырезаются специальными ферментами и отбрасываются, остатки сшиваются. Этот процесс называется сплэйсингом (калька с английского) Я не знал этого слова, но понял значение сразу, вспомнив свою давнюю морскую практику: сплеснивать трос – значит сращивать его из кусков.
После того как все ненужное из первичного транскрипта удалено, к 5’-концу присоединяется «шапочка» – три фосфатных остатка подряд и метилированный нуклеотид. А у 3’-конца вырастает длинный полиадениловый «хвост» – последовательность из многих остатков аденина. Для чего эти добавки – недавно выяснили. Белоксинтезирующие системы клетки – рибосомы – «узнают Сеньку по шапке». 5’-конец, с которого начинается трансляция, по начальным трем фосфатам. Последовательность поли-А придает матричной РНК стабильность, она не так быстро разрушается нуклеазами. Это было показано серией изящных опытов. Так как генетический код един для всего органического мира, можно ввести в клетку чужеродную мРНК и синтезировать совсем другой белок. Этим путем удалось синтезировать в незрелых яйцеклетках шпорцевой лягушки гемоглобин кролика, белки вируса табачной мозаики и пчелиного яда. И каждый раз мРНК, лишенная поли-А последовательности, была нестабильной, распадалась быстро.
А вот для чего гену интроны? На этот счет было высказано немало соображений, вплоть до самых фантастичных: они нужны будто бы для обеспечения процесса эволюции (!). Но ни в одном организме нет ни одной структуры, специально предназначенной для эволюции. Все структуры предназначены только для выживания. Если мы признаем за интронами роль своеобразных органов эволюции, мы наделим природу способностью к прогнозированию и вернемся фактически к учению Аристотеля о будущей причине. Впору подивиться живучести телеологических заблуждений, уже третью тысячу лет воскресающих под разными именами.
А то, что интроны для чего-то нужны не в будущем, а сейчас, ясно из следующего примера. Всем хорошо известный белок инсулин – один из самых маленьких, в нем всего 50 аминокислот. Значит, его ген состоит из 150 нуклеотидов. У крысы два гена инсулина, обозначаемых, как А и В. Оказалось, что в гене А есть один интрон – нечитаемая вставка в 119 нуклеотидов, а в гене В к нему прибавляется другой – в 444 нуклеотида! Из 713 нуклеотидов в процесс трансляции вовлекается только 150 —комментарии излишни.
Не найдем ли мы какой-либо аналогии в человеческих языках? Действительно, во многих языках орфография сильно отличается от произношения. Вот примеры, заимствованные мною у Л. В. Успенского: 1) английское «дочь» пишется daughter читается – «дотэ», 2) ирландское «дочь» – пишется kathudhadh читается «кахю», 3) французское «вода» пишется читается «л’о».
«Лишние», непроизносимые буквы в словах – аналоги интронов в генах. Это объясняется обычно тем, что орфография отстает от произношения и люди пишут так, как говорили несколько веков назад. А то и десятилетий: Анатоль Франс вспоминает бабушку, которая упорно выговаривала «кошемар», «булевар». Да, но почему орфография в одних языках отстает от произношения сильно, а в других за ним поспевает? Почему французское правописание консервативней русского (хотя и мы, особенно в быстрой речи, выговариваем, например, «ПалВаныч» вместо «Павел Иванович»)? Консервативность орфографии не случайна. Она достигает предела в тех языках, где много омонимов – слов с разным значением, но произносимых одинаково.
Во французском языке таких слов очень много, он как бы создан для каламбуров. Но то, что оживляет устную речь, может создать помехи при чтении письменного текста. Так что не будь французская орфография консервативной, французам пришлось бы эту консервативность выдумать.
И не случайно иероглифическая письменность упорно держится Китае. Китайский язык весь состоит из омонимов. В устной речи они распознаются по тону, а как с чтением?
А теперь вернемся к нашим интронам. Регуляторные механизма белкового синтеза, к разгадке которых мы только сейчас приступаем должны как-то «угадывать», «узнавать»[6] нужные гены, чтобы транскрибировать нужную мРНК и затем транслировать нужный белок. Немного пользы организму, если ген гемоглобина будет задействован в нервной клетке, а ген пепсина (пищеварительного фермента) в – мышцах. Генетическая программа не признает омонимов, каламбуры здесь строжайше противопоказаны. Язык генома жестко однозначен, как машинные языки ФОРТРАН или АЛГОЛ, в нем нет места недосказанности размытости значения метафоричности – короче тех особенностей человеческих языков, без которых была бы невозможной изящная словесность. Хромосома ведет себя, как та электронная вычислительная машина которая библейское изречение «Плоть немощна но дух бодр» перевела с английского (The spirit is saund, but the flesh is weak) на русский как «Водка крепкая, но мясо размякло».
Поэтому в гене должна содержаться не только информация об аминокислотных последовательностях. Там должны быть участки, по которым регуляторные элементы клетки его узнают Ясно также, что из окончательной нуклеотидной последовательности мРНК они, как сделавшие свое дело, должны быть удалены. Такими участками и могут оказаться интроны. Это только гипотеза, но на сегодняшний день она наиболее вероятна.
И в заключение рассмотрим важный вопрос: достаточно ли в клетке ДНК для кодирования всех структур сложного фенотипа?
По этому поводу еще недавно велись дискуссии. Казалось, что ДНК явно не хватает. Однако попробуем определить объем генетической информации, как это делал Джон фон Нейман, в битах. Вспомним слова великого физика лорда Кельвина: «Если Вы можете измерить то, о чем говорите, и выразить это в числах, то Вы что-то знаете об этом предмете; если же Вы не в состоянии ни измерить, ни выразить это в числах, то Ваши знания предмета скудны и неудовлетворительны».
Вот схема простенького расчета, когда-то мною проделанного. Если бы все основания в ДНК встречались в одинаковом количестве, вероятность встречи каждого из них была бы 0,25.
Отсюда информационная ценность каждого из оснований
Оценить неоднородность ДНК в геноме можно простым опытом. Если мы будем повышать температуру раствора ДНК, то на каком-то уровне средняя кинетическая энергия молекул окажется выше энергии водородных связей, которыми скреплены половинки двойной спирали. Температура, при которой распадается (денатурирует) половина молекул ДНК, называется температурой плавления. Она сильно зависит от концентрации катионов в растворе (примерно прямо пропорциональна логарифму их концентрации). В паре ГЦ три водородные связи, в паре АТ только две. Поэтому чем больше ГЦ в ДНК, тем более она «тугоплавка». Отсюда следует, что по ширине интервала температур, в котором ДНК плавится, можно судить о ее неоднородности (гетерогенности) в геноме.
Расчет дал около 1,9 бита на основание даже для гетерогенной ДНК млекопитающего (теленка). А число нуклеотидов в геноме млекопитающего около двух-трех миллиардов (у человека два миллиарда, а у буйвола на 40 процентов больше; видно, дело не в количестве). Значит, запас информации в ДНК млекопитающего 4—6 миллиардов бит, что соответствует библиотеке в полторы-две тысячи томов.
Не забудьте, что этот запас содержится в яйцеклетке или головке спермия, имеющей микронные размеры. Какова свертка информации! Сказочный джинн, вылетая из бутылки, вырастает всего-навсего выше финиковой пальмы. Насколько действительность фантастичнее волшебной сказки!
Достаточно ли такого массива информации для постройки фенотипа? Задавшись таким вопросом, ученые спохватились: ведь мы же не умеем оценить сложность фенотипа количественно. В принципе любую структуру, в том числе структуру организма, можно описать с весьма высокой точностью (предел здесь накладывает так называемая квантовомеханическая неопределенность, о которой у нас речь пойдет в следующей главе). Представим, что мы разрежем организм на серию последовательных идущих друг за другом срезов. Расположение структур на каждом из срезов можно описать в двумерной системе координат и выразить объем этой информации в битах. Детальность описания зависит от толщины среза.
Допустим, толщина среза у нас будет один ангстрем (10-10м) – это величина, близкая к пределу разрешающей способности лучших современных электронных микроскопов. Но тогда, скажем, для описания фенотипа человека ростом в 180 сантиметров придется сделать и описать 18 миллиардов таких срезов!
Ясно, что таких опытов никто не проделывал. Все подобные эксперименты оставались мысленными. Структуру поменьше и с меньшим уровнем разрешения, например бактериальную клетку или митохондрию, так описать можно. Увеличивая толщину срезов, скажем, в сто или тысячу раз, мы можем дать описания, но в сто или тысячу раз менее детальные.
А величинам, полученным в результате мысленных экспериментов и приближенных расчетов, как-то не хочется верить. Все это попросту среднепотолочные цифры. Фенотипическую информацию организма человека оценивали и в 105и в 1025бит. Та же величина для бактерии, по данным разных авторов, колеблется от 104до 1012бит!
Но в теории информации существует правило (закон Шеннона): при передаче по любому каналу информация может только теряться за счет помех, но не увеличиваться. Значит, информация, потребная для описания структуры «человек», не должна превышать четырех миллиардов бит.
Вернее, она должна быть существенно меньше. Если в канале информации есть помехи (а они есть в любом канале), информация генотипа должна быть избыточной, многократно повторяться, иметь механизмы коррекции, устранения помех. Впервые я это очень наглядно понял, слушая переговоры по радиотелефону двух судовых радистов: «Аметист, Аметист, я 4347, я 4347, как меня слышите, прием, прием» – «4347, 4347, я Аметист, слышу вас хорошо, слышу вас хорошо, прием, прием».
Ответ не совсем точен: слышали мы хорошо, но из-за треска в динамике понимали плохо.
Примерно так обстоит дело и в канале информации «от ДНК к признакам организма»: не будь генетическая информация высокоизбыточной, новое поколение из-за случайных помех в развитии не походило бы на родителей, получался бы «не мышонок, не лягушка, а неведома зверюшка».

 Рис. 19. Пятерня однояйцевых близнецов (сестры Дионн) в день пятилетия. По особенностям симметрии удалось восстановить их историю до рождения. Оплодотворенная яйцеклетка разделилась на два бластомера, которые разошлись и стали делиться самостоятельно. Из одного получились Ивонна и Анна, из другого Цецилия и еще один бластомер, который, разделившись, дал Марию и Эмилию. В результате получилось пять зародышей с абсолютно идентичными генетическими программами. Насколько полно сходство фенотипов при тождестве генетических программ, вы можете убедиться сами, глядя на лица этих девочек.
Рис. 19. Пятерня однояйцевых близнецов (сестры Дионн) в день пятилетия. По особенностям симметрии удалось восстановить их историю до рождения. Оплодотворенная яйцеклетка разделилась на два бластомера, которые разошлись и стали делиться самостоятельно. Из одного получились Ивонна и Анна, из другого Цецилия и еще один бластомер, который, разделившись, дал Марию и Эмилию. В результате получилось пять зародышей с абсолютно идентичными генетическими программами. Насколько полно сходство фенотипов при тождестве генетических программ, вы можете убедиться сами, глядя на лица этих девочек.
Но мы-то знаем, как удивительно точно черты родителей проявляются в потомстве. Еще более разительный пример точности передачи генетической информации – однояйцевые близнецы, братья и сестры, развившиеся из одной разделившейся яйцеклетки. У них идентичные наборы генов, поэтому только у близнецов удаются пересадки тканей и органов, их путают даже хорошие знакомые и не могут различить по запаху собаки.
 Рис. 20. В мире нет двух людей с одинаковыми отпечатками пальцев. На этом и основана большая область криминалистики – дактилоскопия. Но нет правил без исключений. Я думаю, даже Шерлок Холмс не смог бы различить отпечатки пальцев однояйцевых близнецов (I и II на рисунке). Как вы видите, даже столь, казалось бы, маловажная структура, как дактилоскопический узор, весьма жестко детерминирована генетически.
Рис. 20. В мире нет двух людей с одинаковыми отпечатками пальцев. На этом и основана большая область криминалистики – дактилоскопия. Но нет правил без исключений. Я думаю, даже Шерлок Холмс не смог бы различить отпечатки пальцев однояйцевых близнецов (I и II на рисунке). Как вы видите, даже столь, казалось бы, маловажная структура, как дактилоскопический узор, весьма жестко детерминирована генетически.
Значит, информация, заключенная в генотипе, избыточна. Один из механизмов мы знаем: это двойной набор хромосом в оплодотворенной яйцеклетке. В принципе развитие нормального фенотипа может быть обеспечено половинным, гаплоидным набором хромосом. Случаи партеногенеза, развития неоплодотворенной яйцеклетки, известны у многих животных и растений.
Отсюда следует, что объем информации, закодированный в генотипе, надо уменьшить вдвое. Эти соображения и заставили многих исследователей искать другие источники генетической информации, помимо ДНК.
Эмбриолог Х. Равен, о котором мы уже упоминали, выдвинул предположение, что, помимо ядерной ДНК, хранилищем информации может быть приповерхностный, так называемый кортикальный, слой яйцеклетки. Эта гипотеза не подтвердилась. Не следует забывать, что хранители наследственной информации – гены – должны размножаться, реплицироваться. В противном случае количество их в клетке будет уменьшаться вдвое с каждым делением. Из всех известных нам соединений только нуклеиновые кислоты обладают способностью к репликации. Те клеточные структуры, которые могут размножаться, например энергетические станции клетки, митохондрии и хлорофилловые зерна растений, хлоропласты, имеют свои, автономные геномы, очень похожие на простые бактериальные. Ядру они помочь не могут, хоть бы самим воспроизвестись с минимальной помощью от ядра.
Попробуем подойти к этому вопросу иначе: а правильно ли мы оценили сложность фенотипа?
 Рис. 27. Согласно наиболее распространенной гипотезе, многоклеточные животные произошли от колоний одноклеточных. Сверхорганизмы высшего порядка возникают из колоний многоклеточных, в которых каждый организм выполняет определенную функцию. Таковы колониальные кишечнополостные – сифонофоры. На рисунке наверху: сифонофоры из группы сифонант. Верхние особи них превратились в подобие плавательных пузырей, удерживающих колонию в плавучем состоянии. Другие особи выполняют функции движения, питания и защиты колонии, а часть приспособлена для размножения. Сифонофоры-дисконанты настолько далеко зашли по этому пути, что многие ученые отказываются считать их колонией и относят к единичным гидроидным полипам. Внизу: представитель этой группы – парусник велелла (сектор тела вырезан, чтобы показать сложное строение). Здесь также действует общий принцип – совокупность одинаковых элементов, представляющая информационную систему с высокой избыточностью, превращается в более сложную за счет снижения избыточности.
Рис. 27. Согласно наиболее распространенной гипотезе, многоклеточные животные произошли от колоний одноклеточных. Сверхорганизмы высшего порядка возникают из колоний многоклеточных, в которых каждый организм выполняет определенную функцию. Таковы колониальные кишечнополостные – сифонофоры. На рисунке наверху: сифонофоры из группы сифонант. Верхние особи них превратились в подобие плавательных пузырей, удерживающих колонию в плавучем состоянии. Другие особи выполняют функции движения, питания и защиты колонии, а часть приспособлена для размножения. Сифонофоры-дисконанты настолько далеко зашли по этому пути, что многие ученые отказываются считать их колонией и относят к единичным гидроидным полипам. Внизу: представитель этой группы – парусник велелла (сектор тела вырезан, чтобы показать сложное строение). Здесь также действует общий принцип – совокупность одинаковых элементов, представляющая информационную систему с высокой избыточностью, превращается в более сложную за счет снижения избыточности.
Тот же Равен указывает, что, например, у лошади несколько миллионов печеночных клеток и все они построены одинаково. Можно конечно, определить объем информации, потребный для описания каждой из клеток, а потом умножить число бит на число клеток. Но не равносильно ли это утверждению, что для полного тиража скажем, журнала «Наука и жизнь» требуется авторов, редакторов, корректоров, художников, фотографов и т. д. в три миллиона раз больше, чем для сигнального экземпляра?
Значит, структура фенотипа также информационно избыточна, причем в весьма высокой степени. Генотип может дать подробное описание лишь одной клетки, а затем указать, что она должна повториться сотни тысяч и миллионы раз.
Вот еще хороший пример фенотипической избыточности, который нам еще пригодится в будущем.
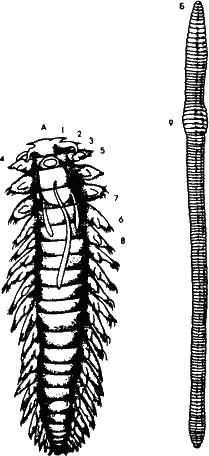 Рис. 21. Для нас чрезвычайно важен вопрос, каким путем в процессе прогрессивной эволюции происходит усложнение структуры, увеличение количества информации, потребной для описания организма. Следует помнить, что новая информация не возникает из ничего. Она возникает из избыточной информации при взаимодействии с шумом. На рисунке два кольчатых черня: многощетинковый червь полихета и малощетинковый – обычный дождевой червь. Как видим, тела их состоят из практически идентичных повторяющихся члеников, то есть информация фенотипа в высокой степени избыточна. Мутация (шум в канале передачи) изменяют форму отдельных члеников, строение конечностей на них и внутренних органов. Если эти изменения окажутся приспособительными, отбор сохранит их. Избыточность при этом снизится, но усложнится структура, и количество информации возрастет. Так идет прогрессивная эволюция.
Рис. 21. Для нас чрезвычайно важен вопрос, каким путем в процессе прогрессивной эволюции происходит усложнение структуры, увеличение количества информации, потребной для описания организма. Следует помнить, что новая информация не возникает из ничего. Она возникает из избыточной информации при взаимодействии с шумом. На рисунке два кольчатых черня: многощетинковый червь полихета и малощетинковый – обычный дождевой червь. Как видим, тела их состоят из практически идентичных повторяющихся члеников, то есть информация фенотипа в высокой степени избыточна. Мутация (шум в канале передачи) изменяют форму отдельных члеников, строение конечностей на них и внутренних органов. Если эти изменения окажутся приспособительными, отбор сохранит их. Избыточность при этом снизится, но усложнится структура, и количество информации возрастет. Так идет прогрессивная эволюция.
Есть довольно просто устроенные морские кольчатые многощетинковые черви – полихеты. Туловище наиболее примитивных из них разделяется на десятки, а то и сотни члеников. И все эти членики устроены по одному образцу, кроме первого, на котором сконцентрированы органы чувств, второго – с ротовым отверстием и последнего – с анальным отверстием. Значит, для описания фенотипа достаточно только четырех члеников, но при этом необходимо добавить: третий по счету повторяется столько-то раз.
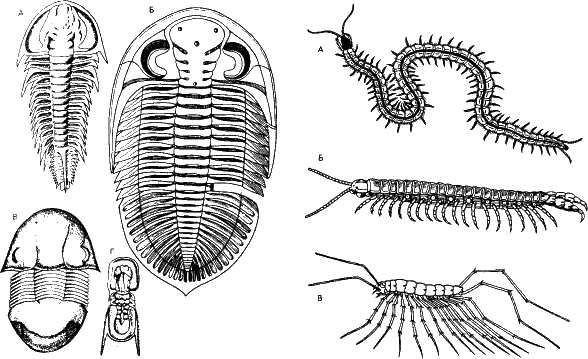 Рис. 22. Вот еще примеры высокоизбыточной фенетической информации. Вымершие членистоногие – трилобиты – имели конечности, практически одинаковые и по форме, и своим функциям: движения, захвата пищи и дыхания. Все это они делали одинаково хорошо (вернее, одинаково плохо). У многоножек (справа) ротовые части – производные конечностей – уже специализированы, однако ноги одинаковы и тело еще не делится, как у насекомых, на грудной и брюшной отделы.
Рис. 22. Вот еще примеры высокоизбыточной фенетической информации. Вымершие членистоногие – трилобиты – имели конечности, практически одинаковые и по форме, и своим функциям: движения, захвата пищи и дыхания. Все это они делали одинаково хорошо (вернее, одинаково плохо). У многоножек (справа) ротовые части – производные конечностей – уже специализированы, однако ноги одинаковы и тело еще не делится, как у насекомых, на грудной и брюшной отделы.
 Рис. 23. Различные представители отряда высших ракообразных. Здесь уже тело поделено на головной, грудной и брюшной отделы, которые несут конечности разного строения. Да и членики устроены по-разному. Структура фенотипа в процессе прогрессивной эволюции усложнилась, и требуется больше информации для ее описания.
Рис. 23. Различные представители отряда высших ракообразных. Здесь уже тело поделено на головной, грудной и брюшной отделы, которые несут конечности разного строения. Да и членики устроены по-разному. Структура фенотипа в процессе прогрессивной эволюции усложнилась, и требуется больше информации для ее описания.
Ракообразные произошли от предков, похожих на кольчатых червей. У высших ракообразных, например у речного рака, все членики устроены по-разному, особенно различаются конечности, и каждый членик поэтому придется описывать отдельно. Объем информации повышается: ведь это более сложный фенотип. И эта новая информация возникает не из ничего – из избыточной информации фенотипа (ценой снижения избыточности).
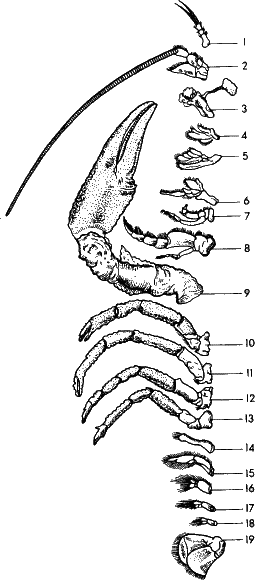 Рис. 24. На предыдущих рисунках вы видели примеры избыточности информации а строении тела членистых животных – кольчатых червей, трилобитов, многоножек. У высших членистых монотонность сменяется разнообразием, новая информация возникает за счет избыточной. У речного рака практически каждую конечность надо описывать отдельно, ибо они выполняют разные функции и имеют разную форму: первая и вторая пара – органы чувств, с третьей по пятую – челюсти, с шестой по восьмую – ногочелюсти (передают пищу челюстям), с девятой по тринадцатую – ходильные, с четырнадцатой по восемнадцатую – брюшные. Последняя пара образует «хвостовой» веер: хлопая им, рак плывет задом наперед (ползает он, как и все прочие животные, головой вперед, а не пятится).
Рис. 24. На предыдущих рисунках вы видели примеры избыточности информации а строении тела членистых животных – кольчатых червей, трилобитов, многоножек. У высших членистых монотонность сменяется разнообразием, новая информация возникает за счет избыточной. У речного рака практически каждую конечность надо описывать отдельно, ибо они выполняют разные функции и имеют разную форму: первая и вторая пара – органы чувств, с третьей по пятую – челюсти, с шестой по восьмую – ногочелюсти (передают пищу челюстям), с девятой по тринадцатую – ходильные, с четырнадцатой по восемнадцатую – брюшные. Последняя пара образует «хвостовой» веер: хлопая им, рак плывет задом наперед (ползает он, как и все прочие животные, головой вперед, а не пятится).
Определяя объем информации потребной для описания фенетической структуры, не следует забывать о связях между признакам и организма. Допустим, нам известно о некоем животном только то, что у него одна левая дуга аорты. Кажется, это очень мало. Ошибаетесь: отсюда однозначно следует, что сердце у него четырехкамерное, эритроциты без ядер, хорошо развитый мозг, постоянная температура тела. Это значит, что оно относится к классу млекопитающих. Такие сцепления признаков называются корреляциями. Впервые их широко применил основатель палеонтологии Жорж Кювье. Известно, что он по отдельной кости уверенно описывал облик животного. Существует анекдот про Кювье: один из его учеников решил над ним подшутить, надел на себя шкуру с рогами и копытами, подошел к учителю ночью и прорычал страшным голосом: «Я съем тебя». Кювье спросонья твердо сказал: «Рога и копыта – значит, ты травоядное и не можешь съесть меня». Случай, конечно, вымышленный, но логику Кювье демонстрирует хорошо.
Впрочем, бывали случаи, когда логика Кювье подводила. Большой изогнутый коготь, найденный отдельно от прочих костей, он приписал муравьеду. На деле коготь принадлежал халикотерию: жили на земле и дожили почти до появления на ней человека странные звери, по всем признакам копытные, но имевшие вместо копыт мощные ногти. Так что корреляция отнюдь не всегда бывает полной.
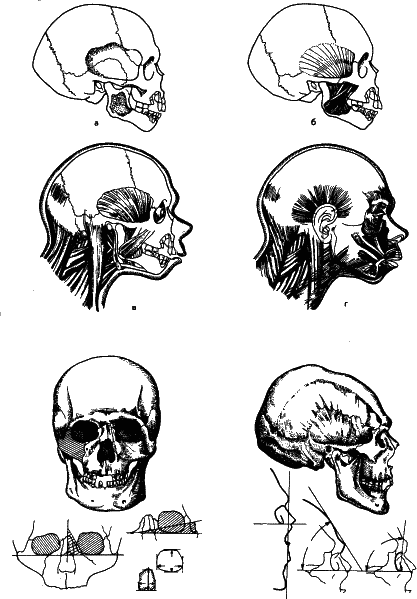
 Рис. 28. В организме одна структура может с большей или меньшей жесткостью определять детали строения другой. Именно это позволило нашему замечательному анатому, антропологу, археологу и скульптору М. М. Герасаимову разработать метод восстановления лица по черепу. Здесь приведены две схемы: реконструкция лица неандертальского мальчика из грота Тешик-Таш и головы скифского воина из богатого погребения. Любопытно, что, когда был изготовлен его скульптурный портрет, историки опознали его по барельефам и монетам. Это оказался скифский царь Скилур.
Рис. 28. В организме одна структура может с большей или меньшей жесткостью определять детали строения другой. Именно это позволило нашему замечательному анатому, антропологу, археологу и скульптору М. М. Герасаимову разработать метод восстановления лица по черепу. Здесь приведены две схемы: реконструкция лица неандертальского мальчика из грота Тешик-Таш и головы скифского воина из богатого погребения. Любопытно, что, когда был изготовлен его скульптурный портрет, историки опознали его по барельефам и монетам. Это оказался скифский царь Скилур.
Наш замечательный антрополог, анатом и скульптор М. М. Герасимов, Руководствуясь теми же принципами что и Кювье разработал методику восстановления лица по черепу, причем с точностью, удовлетворяющей не только археологов, но и работников уголовного розыска. А это возможно осуществить только в одном случае: когда структура одной части организма определяет структуру другой.
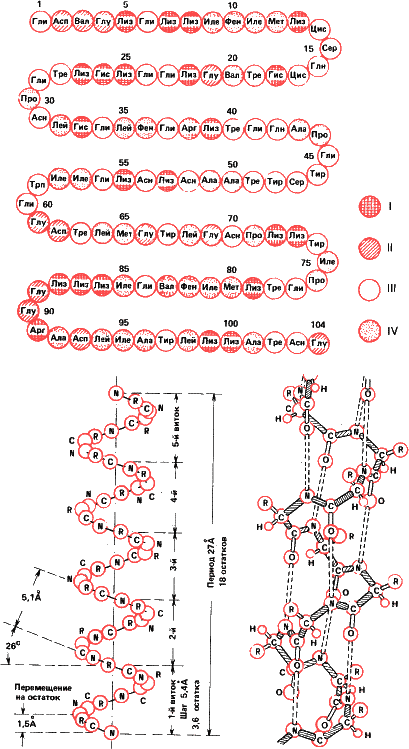 Рис.29. На молекулярном уровне структура высшего уровня также определяется структурой низшего. На рисунке сверху первичная структура белка – цитохрома С, переносчика электронов в дыхательных цепях (обозначения те же, что и на рис. 16). Внизу: схема вторичной структуры – спирали Полинга – Кори. Ее поддерживают водородные связи между группами —CO – и —NH —. Что образуется из полипептида со вторичной структурой, показано на следующем рисунке.
Рис.29. На молекулярном уровне структура высшего уровня также определяется структурой низшего. На рисунке сверху первичная структура белка – цитохрома С, переносчика электронов в дыхательных цепях (обозначения те же, что и на рис. 16). Внизу: схема вторичной структуры – спирали Полинга – Кори. Ее поддерживают водородные связи между группами —CO – и —NH —. Что образуется из полипептида со вторичной структурой, показано на следующем рисунке.
Этот принцип соблюдается в природе начиная с молекулярного уровня. Первичная структура белковой молекулы – это последовательность аминокислотных остатков в полипептиде. Соседние звенья в пептидной цепочке соединяются водородными связями, образуя спиралеобразную фигуру (так называемая спираль Полинга-Кори). Это вторичная структура. Но спираль Полинга также образует трехмерную третичную структуру, специфичную для каждого белка. Наконец, отдельные белковые глобулы могут объединяться попарно и по четыре, а то и больше, образуя четвертичную структуру. Таков, например, гемоглобин.
И все эти структуры определяются одной – первичной. Значит, в генотипе нужно кодировать только последовательность аминокислот, все остальное возникает при соответствующих условиях само.
Перекрывается ли код? Первые гипотетические модели кода были перекрывающимися Это значило, что последовательность нуклеотидов могла кодировать разные аминокислотные последовательности, в зависимости от начала считывания. Так, последовательность АТТГЦАТЦГ, если считывалась бы с А, кодировала бы Тир—Вал—Ала, если со второго нуклеотида (Т)—Вал—Ала—Мет, и т. д. Подобный код накладывал бы жесткие ограничения на последовательность аминокислот в белках. И все облегченно вздохнули, когда С. Бреннер доказал, что каждый триплет нуклеотидов в ДНК и РНК считывается только один раз (неперекрывающийся код).
И опять как гром с ясного неба. Оказалось, что у некоторых фагов гены перекрываются. У фага φX174 имеется двойное перекрытие, у фага G4 даже тройное, то есть с одной нуклеотидной последовательности считываются три аминокислотных! Это предел экономичности сигнала. Фагам хорошо, а вот каково молекулярным генетикам? Как широко распространен этот феномен в мире вирусов? Встречается ли он у высших организмов? Не знаем.
Но самое интересное из новых открытий я приберег под конец.
Конец коллинеарности гена. До последних лет все согласно считали, что ген коллинеарен тому белку, точнее, той полипептидной цепи, которую он кодирует. Иными словами, каждой тройке нуклеотидов в ДНК, с которой считывается мРНК, соответствует один аминокислотиый остаток в полипептиде.
И опять оказалось не так! В смысловой цепи ДНК, кодирующей белок, обнаружены довольно длинные вставки (интроны), никаких аминокислот не кодирующие. Они считываются при синтезе первичного транскрипта, а далее начинается непонятный процесс. Ненужные вставки вырезаются специальными ферментами и отбрасываются, остатки сшиваются. Этот процесс называется сплэйсингом (калька с английского) Я не знал этого слова, но понял значение сразу, вспомнив свою давнюю морскую практику: сплеснивать трос – значит сращивать его из кусков.
После того как все ненужное из первичного транскрипта удалено, к 5’-концу присоединяется «шапочка» – три фосфатных остатка подряд и метилированный нуклеотид. А у 3’-конца вырастает длинный полиадениловый «хвост» – последовательность из многих остатков аденина. Для чего эти добавки – недавно выяснили. Белоксинтезирующие системы клетки – рибосомы – «узнают Сеньку по шапке». 5’-конец, с которого начинается трансляция, по начальным трем фосфатам. Последовательность поли-А придает матричной РНК стабильность, она не так быстро разрушается нуклеазами. Это было показано серией изящных опытов. Так как генетический код един для всего органического мира, можно ввести в клетку чужеродную мРНК и синтезировать совсем другой белок. Этим путем удалось синтезировать в незрелых яйцеклетках шпорцевой лягушки гемоглобин кролика, белки вируса табачной мозаики и пчелиного яда. И каждый раз мРНК, лишенная поли-А последовательности, была нестабильной, распадалась быстро.
А вот для чего гену интроны? На этот счет было высказано немало соображений, вплоть до самых фантастичных: они нужны будто бы для обеспечения процесса эволюции (!). Но ни в одном организме нет ни одной структуры, специально предназначенной для эволюции. Все структуры предназначены только для выживания. Если мы признаем за интронами роль своеобразных органов эволюции, мы наделим природу способностью к прогнозированию и вернемся фактически к учению Аристотеля о будущей причине. Впору подивиться живучести телеологических заблуждений, уже третью тысячу лет воскресающих под разными именами.
А то, что интроны для чего-то нужны не в будущем, а сейчас, ясно из следующего примера. Всем хорошо известный белок инсулин – один из самых маленьких, в нем всего 50 аминокислот. Значит, его ген состоит из 150 нуклеотидов. У крысы два гена инсулина, обозначаемых, как А и В. Оказалось, что в гене А есть один интрон – нечитаемая вставка в 119 нуклеотидов, а в гене В к нему прибавляется другой – в 444 нуклеотида! Из 713 нуклеотидов в процесс трансляции вовлекается только 150 —комментарии излишни.
Не найдем ли мы какой-либо аналогии в человеческих языках? Действительно, во многих языках орфография сильно отличается от произношения. Вот примеры, заимствованные мною у Л. В. Успенского: 1) английское «дочь» пишется daughter читается – «дотэ», 2) ирландское «дочь» – пишется kathudhadh читается «кахю», 3) французское «вода» пишется читается «л’о».
«Лишние», непроизносимые буквы в словах – аналоги интронов в генах. Это объясняется обычно тем, что орфография отстает от произношения и люди пишут так, как говорили несколько веков назад. А то и десятилетий: Анатоль Франс вспоминает бабушку, которая упорно выговаривала «кошемар», «булевар». Да, но почему орфография в одних языках отстает от произношения сильно, а в других за ним поспевает? Почему французское правописание консервативней русского (хотя и мы, особенно в быстрой речи, выговариваем, например, «ПалВаныч» вместо «Павел Иванович»)? Консервативность орфографии не случайна. Она достигает предела в тех языках, где много омонимов – слов с разным значением, но произносимых одинаково.
Во французском языке таких слов очень много, он как бы создан для каламбуров. Но то, что оживляет устную речь, может создать помехи при чтении письменного текста. Так что не будь французская орфография консервативной, французам пришлось бы эту консервативность выдумать.
И не случайно иероглифическая письменность упорно держится Китае. Китайский язык весь состоит из омонимов. В устной речи они распознаются по тону, а как с чтением?
А теперь вернемся к нашим интронам. Регуляторные механизма белкового синтеза, к разгадке которых мы только сейчас приступаем должны как-то «угадывать», «узнавать»[6] нужные гены, чтобы транскрибировать нужную мРНК и затем транслировать нужный белок. Немного пользы организму, если ген гемоглобина будет задействован в нервной клетке, а ген пепсина (пищеварительного фермента) в – мышцах. Генетическая программа не признает омонимов, каламбуры здесь строжайше противопоказаны. Язык генома жестко однозначен, как машинные языки ФОРТРАН или АЛГОЛ, в нем нет места недосказанности размытости значения метафоричности – короче тех особенностей человеческих языков, без которых была бы невозможной изящная словесность. Хромосома ведет себя, как та электронная вычислительная машина которая библейское изречение «Плоть немощна но дух бодр» перевела с английского (The spirit is saund, but the flesh is weak) на русский как «Водка крепкая, но мясо размякло».
Поэтому в гене должна содержаться не только информация об аминокислотных последовательностях. Там должны быть участки, по которым регуляторные элементы клетки его узнают Ясно также, что из окончательной нуклеотидной последовательности мРНК они, как сделавшие свое дело, должны быть удалены. Такими участками и могут оказаться интроны. Это только гипотеза, но на сегодняшний день она наиболее вероятна.
И в заключение рассмотрим важный вопрос: достаточно ли в клетке ДНК для кодирования всех структур сложного фенотипа?
По этому поводу еще недавно велись дискуссии. Казалось, что ДНК явно не хватает. Однако попробуем определить объем генетической информации, как это делал Джон фон Нейман, в битах. Вспомним слова великого физика лорда Кельвина: «Если Вы можете измерить то, о чем говорите, и выразить это в числах, то Вы что-то знаете об этом предмете; если же Вы не в состоянии ни измерить, ни выразить это в числах, то Ваши знания предмета скудны и неудовлетворительны».
Вот схема простенького расчета, когда-то мною проделанного. Если бы все основания в ДНК встречались в одинаковом количестве, вероятность встречи каждого из них была бы 0,25.
Отсюда информационная ценность каждого из оснований
H = – (4 • 0,25 log2 0.25) = 2 бита.Однако ДНК в геноме неоднородна по составу. Для высших организмов, например позвоночных животных, доля пары гуанин – цитозин составляет всего около 40 процентов. Кроме того, в ней имеются фракции, обогащенные парами АТ и ГЦ. У некоторых крабов в хромосомах выявлены последовательности состоящие только из двух оснований А и Т. Информационная ценность нуклеотидного звена в них снижается вдвое:
H = – (2 • 0,25 log2 0.25) = 1 бит.то есть основание может быть только или аденином или же тимином.
Оценить неоднородность ДНК в геноме можно простым опытом. Если мы будем повышать температуру раствора ДНК, то на каком-то уровне средняя кинетическая энергия молекул окажется выше энергии водородных связей, которыми скреплены половинки двойной спирали. Температура, при которой распадается (денатурирует) половина молекул ДНК, называется температурой плавления. Она сильно зависит от концентрации катионов в растворе (примерно прямо пропорциональна логарифму их концентрации). В паре ГЦ три водородные связи, в паре АТ только две. Поэтому чем больше ГЦ в ДНК, тем более она «тугоплавка». Отсюда следует, что по ширине интервала температур, в котором ДНК плавится, можно судить о ее неоднородности (гетерогенности) в геноме.
Расчет дал около 1,9 бита на основание даже для гетерогенной ДНК млекопитающего (теленка). А число нуклеотидов в геноме млекопитающего около двух-трех миллиардов (у человека два миллиарда, а у буйвола на 40 процентов больше; видно, дело не в количестве). Значит, запас информации в ДНК млекопитающего 4—6 миллиардов бит, что соответствует библиотеке в полторы-две тысячи томов.
Не забудьте, что этот запас содержится в яйцеклетке или головке спермия, имеющей микронные размеры. Какова свертка информации! Сказочный джинн, вылетая из бутылки, вырастает всего-навсего выше финиковой пальмы. Насколько действительность фантастичнее волшебной сказки!
Достаточно ли такого массива информации для постройки фенотипа? Задавшись таким вопросом, ученые спохватились: ведь мы же не умеем оценить сложность фенотипа количественно. В принципе любую структуру, в том числе структуру организма, можно описать с весьма высокой точностью (предел здесь накладывает так называемая квантовомеханическая неопределенность, о которой у нас речь пойдет в следующей главе). Представим, что мы разрежем организм на серию последовательных идущих друг за другом срезов. Расположение структур на каждом из срезов можно описать в двумерной системе координат и выразить объем этой информации в битах. Детальность описания зависит от толщины среза.
Допустим, толщина среза у нас будет один ангстрем (10-10м) – это величина, близкая к пределу разрешающей способности лучших современных электронных микроскопов. Но тогда, скажем, для описания фенотипа человека ростом в 180 сантиметров придется сделать и описать 18 миллиардов таких срезов!
Ясно, что таких опытов никто не проделывал. Все подобные эксперименты оставались мысленными. Структуру поменьше и с меньшим уровнем разрешения, например бактериальную клетку или митохондрию, так описать можно. Увеличивая толщину срезов, скажем, в сто или тысячу раз, мы можем дать описания, но в сто или тысячу раз менее детальные.
А величинам, полученным в результате мысленных экспериментов и приближенных расчетов, как-то не хочется верить. Все это попросту среднепотолочные цифры. Фенотипическую информацию организма человека оценивали и в 105и в 1025бит. Та же величина для бактерии, по данным разных авторов, колеблется от 104до 1012бит!
Но в теории информации существует правило (закон Шеннона): при передаче по любому каналу информация может только теряться за счет помех, но не увеличиваться. Значит, информация, потребная для описания структуры «человек», не должна превышать четырех миллиардов бит.
Вернее, она должна быть существенно меньше. Если в канале информации есть помехи (а они есть в любом канале), информация генотипа должна быть избыточной, многократно повторяться, иметь механизмы коррекции, устранения помех. Впервые я это очень наглядно понял, слушая переговоры по радиотелефону двух судовых радистов: «Аметист, Аметист, я 4347, я 4347, как меня слышите, прием, прием» – «4347, 4347, я Аметист, слышу вас хорошо, слышу вас хорошо, прием, прием».
Ответ не совсем точен: слышали мы хорошо, но из-за треска в динамике понимали плохо.
Примерно так обстоит дело и в канале информации «от ДНК к признакам организма»: не будь генетическая информация высокоизбыточной, новое поколение из-за случайных помех в развитии не походило бы на родителей, получался бы «не мышонок, не лягушка, а неведома зверюшка».
Но мы-то знаем, как удивительно точно черты родителей проявляются в потомстве. Еще более разительный пример точности передачи генетической информации – однояйцевые близнецы, братья и сестры, развившиеся из одной разделившейся яйцеклетки. У них идентичные наборы генов, поэтому только у близнецов удаются пересадки тканей и органов, их путают даже хорошие знакомые и не могут различить по запаху собаки.
Значит, информация, заключенная в генотипе, избыточна. Один из механизмов мы знаем: это двойной набор хромосом в оплодотворенной яйцеклетке. В принципе развитие нормального фенотипа может быть обеспечено половинным, гаплоидным набором хромосом. Случаи партеногенеза, развития неоплодотворенной яйцеклетки, известны у многих животных и растений.
Отсюда следует, что объем информации, закодированный в генотипе, надо уменьшить вдвое. Эти соображения и заставили многих исследователей искать другие источники генетической информации, помимо ДНК.
Эмбриолог Х. Равен, о котором мы уже упоминали, выдвинул предположение, что, помимо ядерной ДНК, хранилищем информации может быть приповерхностный, так называемый кортикальный, слой яйцеклетки. Эта гипотеза не подтвердилась. Не следует забывать, что хранители наследственной информации – гены – должны размножаться, реплицироваться. В противном случае количество их в клетке будет уменьшаться вдвое с каждым делением. Из всех известных нам соединений только нуклеиновые кислоты обладают способностью к репликации. Те клеточные структуры, которые могут размножаться, например энергетические станции клетки, митохондрии и хлорофилловые зерна растений, хлоропласты, имеют свои, автономные геномы, очень похожие на простые бактериальные. Ядру они помочь не могут, хоть бы самим воспроизвестись с минимальной помощью от ядра.
Попробуем подойти к этому вопросу иначе: а правильно ли мы оценили сложность фенотипа?
Тот же Равен указывает, что, например, у лошади несколько миллионов печеночных клеток и все они построены одинаково. Можно конечно, определить объем информации, потребный для описания каждой из клеток, а потом умножить число бит на число клеток. Но не равносильно ли это утверждению, что для полного тиража скажем, журнала «Наука и жизнь» требуется авторов, редакторов, корректоров, художников, фотографов и т. д. в три миллиона раз больше, чем для сигнального экземпляра?
Значит, структура фенотипа также информационно избыточна, причем в весьма высокой степени. Генотип может дать подробное описание лишь одной клетки, а затем указать, что она должна повториться сотни тысяч и миллионы раз.
Вот еще хороший пример фенотипической избыточности, который нам еще пригодится в будущем.
Есть довольно просто устроенные морские кольчатые многощетинковые черви – полихеты. Туловище наиболее примитивных из них разделяется на десятки, а то и сотни члеников. И все эти членики устроены по одному образцу, кроме первого, на котором сконцентрированы органы чувств, второго – с ротовым отверстием и последнего – с анальным отверстием. Значит, для описания фенотипа достаточно только четырех члеников, но при этом необходимо добавить: третий по счету повторяется столько-то раз.
Ракообразные произошли от предков, похожих на кольчатых червей. У высших ракообразных, например у речного рака, все членики устроены по-разному, особенно различаются конечности, и каждый членик поэтому придется описывать отдельно. Объем информации повышается: ведь это более сложный фенотип. И эта новая информация возникает не из ничего – из избыточной информации фенотипа (ценой снижения избыточности).
Определяя объем информации потребной для описания фенетической структуры, не следует забывать о связях между признакам и организма. Допустим, нам известно о некоем животном только то, что у него одна левая дуга аорты. Кажется, это очень мало. Ошибаетесь: отсюда однозначно следует, что сердце у него четырехкамерное, эритроциты без ядер, хорошо развитый мозг, постоянная температура тела. Это значит, что оно относится к классу млекопитающих. Такие сцепления признаков называются корреляциями. Впервые их широко применил основатель палеонтологии Жорж Кювье. Известно, что он по отдельной кости уверенно описывал облик животного. Существует анекдот про Кювье: один из его учеников решил над ним подшутить, надел на себя шкуру с рогами и копытами, подошел к учителю ночью и прорычал страшным голосом: «Я съем тебя». Кювье спросонья твердо сказал: «Рога и копыта – значит, ты травоядное и не можешь съесть меня». Случай, конечно, вымышленный, но логику Кювье демонстрирует хорошо.
Впрочем, бывали случаи, когда логика Кювье подводила. Большой изогнутый коготь, найденный отдельно от прочих костей, он приписал муравьеду. На деле коготь принадлежал халикотерию: жили на земле и дожили почти до появления на ней человека странные звери, по всем признакам копытные, но имевшие вместо копыт мощные ногти. Так что корреляция отнюдь не всегда бывает полной.
Наш замечательный антрополог, анатом и скульптор М. М. Герасимов, Руководствуясь теми же принципами что и Кювье разработал методику восстановления лица по черепу, причем с точностью, удовлетворяющей не только археологов, но и работников уголовного розыска. А это возможно осуществить только в одном случае: когда структура одной части организма определяет структуру другой.
Этот принцип соблюдается в природе начиная с молекулярного уровня. Первичная структура белковой молекулы – это последовательность аминокислотных остатков в полипептиде. Соседние звенья в пептидной цепочке соединяются водородными связями, образуя спиралеобразную фигуру (так называемая спираль Полинга-Кори). Это вторичная структура. Но спираль Полинга также образует трехмерную третичную структуру, специфичную для каждого белка. Наконец, отдельные белковые глобулы могут объединяться попарно и по четыре, а то и больше, образуя четвертичную структуру. Таков, например, гемоглобин.
И все эти структуры определяются одной – первичной. Значит, в генотипе нужно кодировать только последовательность аминокислот, все остальное возникает при соответствующих условиях само.