Наблюдая за хромосомами во время деления клеток, исследователи обнаружили много форм изменения наследственных программ.
Читателям должно быть известно, что при образовании половых клеток хромосомы не делятся, а расходятся в дочерние клетки, так что получаются гаметы с половинным (гаплоидным) набором хромосом. У человека, например, в нормальных клетках 46 хромосом, а в яйцеклетках и спермиях – 23. При слиянии гамет диплоидный набор восстанавливается.
Но так бывает не всегда. Порой механизм, растягивающий хромосомы по дочерним клеткам, не срабатывает. Одна гамета получается совсем без ДНК, а другая с двойным ее набором. Так возникают полиплоидные клетки и организмы; о часто это наблюдается у растений.
Иногда же в одну клетку попадает лишняя хромосома, а в другой обнаруживается нехватка. Такие явления называются анеуплоидией.
При всех этих перестройках генетическая информация, заключенная в хромосомах, не изменяется. Меняется только ее количество. Полиплоидные клетки, например, могут иметь тройной, четверной и т. д. – до тысячи и более раз! – набор генов.
Анеуплоидный геном – это многотомное, но разрозненное издание. В одной клетке не хватает «тома» инструкций (обычно такие случаи у высших организмов летальны) в другой два одинаковых. Организмы с лишней хромосомой (трисомики) также часто гибнут на ранних стадиях развития или же развиваются с серьезными дефектами. Много таких случаев описано относительно человека.
Иногда перестройка может привести к тому, что хромосома распадается на части. Судьба частей различна: они могут потеряться (делеция), снова воссоединиться в составе прежней хромосомы (иногда в перевернутом виде – инверсия) или же присоединиться к другой (транслокация). Все перестройки, как правило, для организма небезразличны.
Все упоминавшиеся изменения хромосом начинаются с разрыва нуклеотидной цепи ДНК – знаменитой двойной спирали. Поэтому мы должны от тех построек, которые видны в оптический микроскоп, перейти на молекулярный уровень.
Насколько прочны фосфодиэфирные связи, скрепляющие полимерную ДНК, и насколько устойчивы пуриновые и пиримидиновые основания в ДНК к внешним воздействиям?
Это удалось установить с достаточной точностью. Чтобы вызвать единичную мутацию – наследственное изменение генетической программы, – требуется подвести каким-то способом к ДНК энергию в 2,5—3 электронвольта (эВ). Электронвольт – единица энергии: такую энергию приобретает электрон, ускоряемый напряжением в 1 вольт.
Много это или мало? Ведь ДНК в клетке находится в окружении молекул, движущихся с весьма высокой скоростью. Оказывается, что средняя энергия теплового движения молекул при тех температурах, когда жизнь возможна, составляет примерно 1/40 эВ. Иными словами, при физиологических температурах ДНК оказывается достаточно стабильной. Но проблема эта сложнее, чем кажется на первый взгляд.
Не следует забывать, что скорости молекул при хаотическом тепловом движении неодинаковы. Убедиться в этом нетрудно. В 1827 году шотландский ботаник Р. Броун, разглядывая в микроскоп каплю воды с пыльцой растений, обнаружил, что взвешенные в жидкости пыльцевые зерна микронного размера не остаются на месте, а хаотически движутся как бы непрерывно подталкиваемые беспорядочными ударами чего-то невидимого.
Достойно удивления, что Броуново движение не привлекало внимания физиков (может быть, потому, что открыл его ботаник?) до начала нашего века, до исследований Альберта Эйнштейна, польского физика Мариана Смолуховского и французского физика Жана Перрена. А ведь из него не только вытекала непреложность существования молекул, но и возможность оценить их скорости и размеры!
Что происходит с частицей при броуновском движении? Со всех сторон она подвергается ударам молекул. Если она имеет достаточно большие размеры, то удары со всех сторон оказываются скомпенсированными – частица остается на месте. Но если размер ее, допустим, 10-5см, то весьма вероятно, что с какой-либо стороны суммарный импульс будет больше, и частица сдвинется в непредсказуемую сторону.
Иначе и быть не может: ведь скорости молекул разные и флуктуации в их распределении неизбежны. Именно от этих флуктуаций зависит голубой цвет ясного неба, так как на них сильнее рассеиваются синие лучи. Будь скорости всех молекул одинаковыми, солнечный свет не рассеивался бы и солнце светило бы в черном небе, как в космическом пространстве.
Эти же флуктуации кладут предел усилению слабых сигналов в электрических цепях. В конце концов мы слышим лишь треск, результат теплового движения электронов в цепях усилителя. Чтобы уйти за этот предел, приходится охлаждать приемник жидким азотом, водородом, а то и гелием.
Отсюда однозначно следует, что в любой достаточно большой популяции молекул неизбежно найдутся такие, которые могут нарушить структуру гена и вызвать мутацию. Ясно, что такие изменения генетических программ должны обладать следующими свойствами:
1. Они случайны в том смысле, что вероятность каждого единичного изменения не равна единице. Более стабильные части гена мутируют с меньшей частотой, более лабильные – с большей, но мы можем говорить лишь о большей или меньшей вероятности мутаций.
2. Они непредсказуемы, поскольку для предсказания какой-либо мутации мы должны знать координаты и импульсы всех молекул данной клетке.
3. Они не направлены в том смысле, что изменяют генетичеекую программу без учета содержания сохраняющейся в ней информации. Поэтому они только случайно могут оказаться адаптивными, приспособительными.
Не одна температура изменяет содержание генетических программ.
Пожалуй, большее значение имеют кванты жесткого излучения начиная с ультрафиолета, быстро движущиеся элементарные частицы, молекулы веществ, способные реагировать с ДНК (химические мутагены).
Начнем с радиации. Растворы чистых нуклеиновых кислот в видимом свете прозрачны. Однако и ДНК и РНК сильно поглощают ультрафиолет. Максимум поглощения на 260 нанометрах, кванты такой энергии уже могут вызывать и вызывают мутации. Еще более эффективны рентгеновы лучи и гамма-излучение, а также быстрые частицы, возникающие при ядерных распадах. Действие жестких излучений на генетические программы изучает целая отрасль генетики радиогенетика. О ее достижениях в популярной литературе много писалось. Остановимся подробнее на другом вопросе. В случае, когда мутацию вызывает квант или элементарная частица, наиболее четко проступает принцип необходимости квантовой механики для описания мутагенеза.
Мы уже договорились с самого начала, что физические законы, физические аксиомы должны оправдываться и для тех случаев, когда мы имеем дело с живой материей.
Значит, и здесь должен действовать принцип квантовомеханической неопределенности, который впервые сформулировал В. Гейзенберг в 1927 году.
Выражается этот великий принцип на вид простой формулой
Произведение погрешности в определении координаты (Δx) на погрешность в определении импульса (Δp) не может быть меньше постоянной Планка h, равной 6,626 • 10–34Дж•с. Это ничтожная величина, поэтому, живя в макромире, мы ее не замечаем. Будь она побольше, мы бы жили в мире без ясных очертаний предметов и без четких траекторий движения. Но ведь мутации происходят в микромире. Для всего гена неопределенность пренебрежимо мала. Однако чтобы вызвать мутацию, нужно довести определенное количество энергии до участка гена размером всего лишь 10-7см (так называемый радиус эффективного обмена, впервые измеренный Н. В. Тимофеевым-Ресовским и К. Циммером). Здесь уже принципом неопределенности пренебрегать нельзя. О том, что для описания мутагенеза необходимо использовать принцип Гейзенберга, писали еще Н. В. Тимофеев-Ресовский и Паскуаль Йордан в 40-х годах. В последнее время то же утверждает и Манфред Эйген. Короче, мы можем лишь с некоторой долей вероятия предполагать, обладает ли частица нужной для мутации энергией и попадет ли она в зону эффективного объема.
Не только принцип Гейзенберга – любое другое положение квантовой механики делает процесс мутагенеза вероятностным. Вот еще пара примеров. Кроме стабильных атомов углерода и фосфора в состав ДНК могут входить их радиоактивные изотопы. Вероятность этого возрастает с увеличением радиоактивного фона нашей планеты. Местоположение атомов C14 и P32 в нитях ДНК, разумеется, случайно. А радиоактивный распад – тоже случайный процесс: знать, какой из атомов в данный момент распадется и вызовет мутацию, нам не дано, и запрет этот принципиальный.
Наконец, есть мутации, не индуцированные каким-нибудь внешним по отношению к ДНК агентом, а так называемые спонтанные. Водородные связи, скрепляющие двойную спираль ДНК, обусловлены ионами водорода (протонами). Если при раскручивании спирали ДНК, которое обязательно происходит при ее репликации, оба протона перейдут к одному из оснований, возможны другие комбинации, кроме АТ и ГЦ. А неверное спаривание оснований – мутация. Переход протона от одного основания к другому называется туннельным, описывается уравнениями квантовой механики, и все, что мы можем сказать о нем, то, что он осуществляется с частотой 1011…1012в секунду. Время и место каждой единичной спонтанной мутации непредсказуемо принципиально.
А что же вещества, изменяющие структуру гена, – мутагены? Впервые в 1932—1934 годах советские ученые В. В. Сахаров и М. Е. Лобашов независимо друг от друга получили мутации у дрозофилы, действуя на мушек йодом и рядом других веществ. С того времени список мутагенов возрос тысячекратно, теперь каждое химическое соединение, которое предполагается внедрять в практику (лекарства, красители, пластмассы и т. д.), испытывается на мутагенный эффект. Открыты – И. А. Рапопортом и Ш. Ауэрбах – так называемые супермутагены, вызывающие мутации у 100 процентов обработанных ими особей.
Может показаться, что химические мутагены не имеют особого значения в природе, во всяком случае до появления химической промышленности. Простой пример свидетельствует, что это не так. Даже такое, с позволения сказать, вещество, как ион водорода, может индуцировать мутации.
Активная реакция среды измеряется, как вы знаете, в единицах pH. Это взятый с обратным знаком логарифм концентрации водородных ионов. Например, когда реакция нейтральная, pH равно семи и концентрация ионов водорода 10-7моля. ДНК довольно устойчива к подкислению или подщелачиванию, однако при pH ниже четырех начинается отщепление пуриновых оснований – гуанина и аденина.
Содержимое клетки в общем-то сильная буферная система, поддерживающая pH в районе семи (это не относится к специализированным органам вроде желудка, где высока концентрация соляной кислоты, или же слюнных желез некоторых улиток, которые могут выделять довольно концентрированную серную кислоту). Но вспомним о размерах клеток и обязательных флуктуациях молекул. Объем бактериальной клетки всего 2•10-12см3, значит, при pH = 7 в ней всего примерно 120 водородных ионов. А это означает, что очень вероятна флуктуация: ионы водорода соберутся около, например, гуанина и отщепят его от пентознофосфатного остова ДНК. Произойдет мутация, и опять же случайным, непредсказуемым образом.
На этом можно было бы и закончить обзор изменений генетических программ. Но хотелось бы рассказать о недавно открытых хромосомных перестройках, удивительных и ранее казавшихся невозможными.
Сначала в геномах бактерий, а потом и в геномах высших организмов обнаружили удивительные «кусочки» ДНК, которые могут перемещаться с места на место в хромосоме. Их образно назвали прыгающими, генами. Различают две категории таких последовательностей. Есть короткие (500—1500 пар нуклеотидов) и длинные (более двух тысяч). Они могут вычленяться из единственной хромосомы бактерии (этот процесс называется эксцизией, выстриганием) и встраиваться в нее в другом месте (инсерция). И не только в хромосому, прыгающие гены могут включаться в плазмиды – последовательности ДНК, существующие в бактериальной клетке независимо. Так же легко прыгающие гены могут встраиваться в геномы бактериофагов. Удивительного в этом ничего нет: существует вполне убедительное мнение о том, что плазмиды – «домашние», прирученные бактерией фаги.
Короткие инсерционные сегменты обладают многими удивительными свойствами. Почему-то оба конца их состоят из одинаковых последовательностей. Иногда же концевые участки перевервуты: на одном конце, например, АТГАГ, на другом – ГАГТА. Встраиваясь в ген, они часто блокируют процесс транкрипции – ген теряет активность. И наоборот, встраиваясь в перевернутом виде, они могут тот же ген активировать. Иными словами, они включают и выключают гены.
Еще более удивительны длинные прыгающие последовательности транспозоны. Именно они, встраиваясь в фаги, переносят от бактерии к бактерии гены устойчивости к антибиотикам. Так что фаги не только могут убивать бактерии или быть их «приживальщиками». Они переносят в мире микроорганизмов генетическую информацию. Так, насекомые не только вредят растениям, но и опыляют их. Ведь первые цветковые растения опылялись ветром. На их пыльниках кормилось много насекомых. Перепачканные пыльцой жуки и примитивные бабочки перелетали с пыльника на пыльник, и в результате возникли такие пары «растение – насекомое», которые не могут существовать друг без друга. Юкка, например, не может опыляться иначе как с помощью юкковой моли. И между фагом, лизирующим, «пожирающим» бактерию, фагом-«приживальщиком» и приносящей пользу плазмидой, которая переносит транспозоны, есть все переходные ступени.
Прыгающие гены описаны и у высших, ядерных организмов. У дрожжевых грибков они, например, в зависимости от положения в хромосоме, определяют пол клетки. Обнаружили их также у кукурузы и дрозофилы. Вот на последнем объекте хотелось бы остановиться подробнее.
Генетиков давно занимали случаи, когда гены в популяциях плодовой мушки-дрозофилы вдруг теряли стабильность и частота мутаций возрастала во много раз. Оказалось, что нестабильность того или иного гена обусловливается встроившимся в него инсерционным сегментом. В некоторых случаях удалось проследить, как прыгающий ген в течение нескольких лет кочует от одного участка хромосомы к другому и от одной хромосомы к другой, каждый раз вызывая новую мутацию. Шведские ученые Г. Исинг и К. Рамель проследили до сорока прыжков одной такой инсерции. Результатом была или полная инактивация гена, приводящая к смерти (летальная мутация), или же мутации «розовые глаза», «грубые глаза» (с неправильным расположением фасеток) и целый ряд других.
Такие феномены наблюдались не только в экспериментальных, рассаженных по пробиркам популяциях дрозофилы, но и в природе. Известны так называемые «модные» мутации. Название несколько неожиданное, но удачное. Например, в 30-х годах частота мутаций, вызывающих желтую окраску тела и белую окраску глаз, возросла в сотни раз, и эта «мода» длилась до 40-х годов. В 1968 году у дрозофил возникла «мода» на мутацию abnormal abdomen (брюшной отдел уродливой формы), которая продолжалась пять лет и была вытеснена лишь мутацией «опаленные щетинки».
По-видимому, причина «модных» мутаций – вирусы. Известно, что вирусы могут встраиваться в хромосомы не только бактерий, но и ядерных организмов (это доказано для аденовирусов, вируса герпеса и опухолеродных перерождающих нормальную клетку в злокачественную). Инсерция вируса в геном вызывает мутацию того гена, в который он встроился.
Примечательно, что есть явное сходство между интронами, о которых говорилось в прошлой главе, и инсерционными сегментами. Те и другие могут разделять смысловую часть гена, не находят отражения в аминокислотной последовательности кодируемого геном белка и, по-видимому, могут включать и выключать гены.
А что значит: включать и выключать?
По-видимому, это выглядит так: регуляторные элементы клетки могут узнавать и не узнавать встроенный в ген интрон (инсерционный сегмент). В первом случае идет транскрипция предшественника мРНК, во втором – нет. В конце концов, это находит отражение в изменениях признаков фенотипа. И если интрон перепрыгнет в другой ген, то он будет активировать его, а не тот, из которого он ушел.
Третья аксиома. Теперь пора подытожить все, что мы знаем о наследственных изменениях генетических программ и сформулировать аксиому биологии № 3.
Прежде всего, эти изменения случайны и ненаправленны. Их можно сравнить с шум в канале информации от родителей к потомкам. Если мутация полностью исказит смысл передаваемой по цепи поколений инструкции, она будет смертельной (летальной). Это бывает, когда в результате мутации блокируется синтез жизненно важного фермента.
Наоборот, часты случаи, когда мутация не сказывается на признаках фенотипа. Вспомним, что генетический код вырожден и одна и та же аминокислота кодируется несколькими кодонами. Если в результате мутации один кодон заменится другим, но синонимичным, в полипептидную цепь включится та же аминокислота и изменения фенотипа мы не обнаружим.
Между этими двумя полюсами лежит огромная область мутаций, так или иначе изменяющих фенотип. В каких-то конкретных условиях они могут оказаться полезными, носители их с большей вероятностью оставят потомство и передадут их потомству.
Это и есть дарвиновская неопределенная изменчивость – исходный материал для эволюции.
Аксиому биологии №3 мы можем сформулировать так:
В процессе передачи из поколения в поколение генетические программы в результате многих причин изменяются случайно и ненаправленно, и лишь случайно эти изменения оказываются приспособительными.
Третья аксиома вытекает из важнейших физических постулатов – из практической невозможности знать координаты и импульсы всех молекул в клетке, из чего следует распределение энергий молекул по Максвеллу, и из принципиальной невозможности достаточно точно определить координаты и импульсы частиц, атакующих гены (принцип Гейзенберга).
А так как и перераспределение энергий в совокупности молекул, и взаимодействие молекул в химических реакциях в конце концов можно свести к квантовомеханическим процессам, то и получается, что аксиома №3 – следствие квантовой механики.
Далеко не всем это нравится. М. Эйген в книге, на которую я уже ссылался, по этому поводу вспоминает Эйнштейна. В письме от 4 декабря 1926 года Эйнштейн писал Максу Борну: «Квантовая механика внушает большое почтение. Но внутренний голос говорит мне, что это не истинный Иаков. Теория дает много, но едва ли она подводит нас ближе к тайне Старика. Во всяком случае, я убежден, что он не играет в кости…»
Старик – это бог, Не следует считать Эйнштейна религиозным. Поклонник Спинозы, он называл богом реально существующую природу.
С тех пор прошло более 50 лет: Квантовая механика многократно показала свою непротиворечивость природе, и вряд ли даже Эйнштейн сейчас выступал бы против нее. Однако в биологии до сих пор есть авторы, не признающие ненаправленной случайной изменчивости по Дарвину.
В конечном счете они склоняются к точке зрения, высказанной за полвека до Дарвина в 1809 году великим французским натуралистом Ламарком. Согласно Ламарку, организмы в процессе эволюции изменяются, но изменения эти массовые, направленные и приспособительные. Со времени Ламарка их называли по-разному: наследованием благоприобретенных свойств, ассимиляцией условий внешней среды и т. д. и т. д.
В нашей литературе наиболее основательно эту точку зрения отстаивал крупный ихтиолог и географ Л. С. Берг. Он, правда, называл свои воззрения не ламаркизмом, а номогенезом (эволюцией на основе закономерностей, то есть закономерных, а не случайных изменений). Но хотя в своей книге, вышедшей в 1922 году, Берг решительно отрекся от ламаркизма, никого это не обмануло. Уже тогда оппоненты Берга поняли, что ламаркизм и номогенез различаются лишь терминологией.
Ибо, по Бергу, наследственные изменения захватывают массы особей, идут направленно и должны быть изначально целесообразными, приспособительными. Берг, как и Ламарк, вообще считал приспособительные реакции изначальным свойством живого.
Современные номогенетики как-то стыдливо обходят последний тезис Берга, предпочитая говорить лишь о направленности наследственных изменений, а не о целесообразности их. Но направленные мутации должны быть приспособительными – ведь все организмы на Земле к каким-то условиям приспособлены.
Мне доводилось слышать выступление физика, в котором выдвигалась такая идея. Мутации сами по себе вредны или нейтральны и обеспечить эволюцию материалом не могут. Движущая сила эволюции – приспособительные изменения, которые происходят крайне редко (например, раз в тысячу лет). Из-за их редкости мы их обнаружить не можем, поэтому все опыты, опровергающие приспособительное изменение генетических программ, оказываются бездоказательными.
Но физик должен был бы понять, что номогенез, или ламаркизм, – называйте, как хотите, – основан на нарушении принципа причинности. Следствие (приспособленность) в нем определяет причину (изменение генетической программы). И опять мы возвращаемся к Аристотелю с его конечной причиной и целью одновременно. Такие взгляды на эволюцию, называются телеологическими.
Для меня представляется совершенно непостижимым, что в наши дни находятся вполне серьезные ученые, не понимающие того, что было предельно ясно еще в прошлом веке Герцену. Вспомните его «Былое и думы»:
Допустим существование неких актов приспособления, чрезвычайно редких и потому ненаблюдаемых. Нет проще приема сделать их наблюдаемыми. Надо лишь увеличить число наблюдений. Пример из той же физики: нейтрино взаимодействует с веществом с чрезвычайно низкой вероятностью, эта элементарная частица может пронизать насквозь земной шар, и вероятность того, что она вступит в реакцию с каким-либо ядром, совершенно ничтожна. Но физики уверенно детектируют нейтрино и антинейтрино. Детектор здесь – большое количество атомов хлора (обычно в составе той жидкости, которая применяется в химчистке – тетрахлорэтилена). А источник искомой частицы – ядерный реактор, испускающий мощные потоки нейтрино.
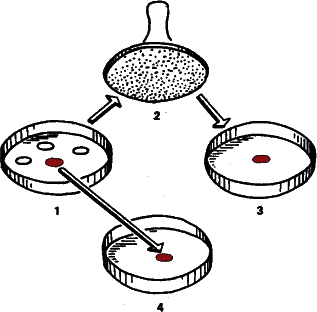 Рис. 36. Супруги Ледерберги выращивали в чашках Петри на поверхности агара тысячи колоний бактерий, а затем стерильным бархатным штампом переносили их с сохранением прежнего положения в другие чашки, где среда содержала смертельную дозу антибиотика. Если число пересеваемых колоний было достаточно велико, всегда находилась хотя бы одна, устойчивая к антибиотику. При этом нетрудно было убедиться, что материнская колония также состоит из устойчивых бактерий, хотя с антибиотиками они и не встречались.
Рис. 36. Супруги Ледерберги выращивали в чашках Петри на поверхности агара тысячи колоний бактерий, а затем стерильным бархатным штампом переносили их с сохранением прежнего положения в другие чашки, где среда содержала смертельную дозу антибиотика. Если число пересеваемых колоний было достаточно велико, всегда находилась хотя бы одна, устойчивая к антибиотику. При этом нетрудно было убедиться, что материнская колония также состоит из устойчивых бактерий, хотя с антибиотиками они и не встречались.
Американские исследователи Джошуа и Эстер Ледерберги исследовали процесс приспособления бактерий к антибиотикам. Этот феномен – проклятие современной медицины. Прославленные пенициллин, стрептомицин, олеандомицин быстро теряют свою эффективность Нужно сказать, что помогают микробам в данном случае и врачи со слабой биологической подготовкой. Они прописывают больным недостаточные дозы (из боязни «как бы чего не вышло»), то есть фактически ведут в популяциях бактерий отбор на устойчивость к антибиотику. Или же, наоборот, впадают в другую крайность и закармливают пациента антибиотиками при пустяковой простуде, а в результате тот же антибиотик окажется недейственным при двустороннем воспалении легких. Вспомните о переносах генов плазмидами и бактериофагами! Ген, определяющий, например, устойчивость бактерии к стрептомицину, легко может быть передан даже бактериям другого вида. В последнее время возникла серьезная проблема с гноеродными золотистыми стафилококками – современный врач с набором антибиотиков оказался в положении хирурга прошлого века, бессильного перед послеоперационными осложнениями. Выход в одном – все время обновлять арсенал антибиотиков, по возможности обгоняя бактерий.
Читателям должно быть известно, что при образовании половых клеток хромосомы не делятся, а расходятся в дочерние клетки, так что получаются гаметы с половинным (гаплоидным) набором хромосом. У человека, например, в нормальных клетках 46 хромосом, а в яйцеклетках и спермиях – 23. При слиянии гамет диплоидный набор восстанавливается.
Но так бывает не всегда. Порой механизм, растягивающий хромосомы по дочерним клеткам, не срабатывает. Одна гамета получается совсем без ДНК, а другая с двойным ее набором. Так возникают полиплоидные клетки и организмы; о часто это наблюдается у растений.
Иногда же в одну клетку попадает лишняя хромосома, а в другой обнаруживается нехватка. Такие явления называются анеуплоидией.
При всех этих перестройках генетическая информация, заключенная в хромосомах, не изменяется. Меняется только ее количество. Полиплоидные клетки, например, могут иметь тройной, четверной и т. д. – до тысячи и более раз! – набор генов.
Анеуплоидный геном – это многотомное, но разрозненное издание. В одной клетке не хватает «тома» инструкций (обычно такие случаи у высших организмов летальны) в другой два одинаковых. Организмы с лишней хромосомой (трисомики) также часто гибнут на ранних стадиях развития или же развиваются с серьезными дефектами. Много таких случаев описано относительно человека.
Иногда перестройка может привести к тому, что хромосома распадается на части. Судьба частей различна: они могут потеряться (делеция), снова воссоединиться в составе прежней хромосомы (иногда в перевернутом виде – инверсия) или же присоединиться к другой (транслокация). Все перестройки, как правило, для организма небезразличны.
Все упоминавшиеся изменения хромосом начинаются с разрыва нуклеотидной цепи ДНК – знаменитой двойной спирали. Поэтому мы должны от тех построек, которые видны в оптический микроскоп, перейти на молекулярный уровень.
Насколько прочны фосфодиэфирные связи, скрепляющие полимерную ДНК, и насколько устойчивы пуриновые и пиримидиновые основания в ДНК к внешним воздействиям?
Это удалось установить с достаточной точностью. Чтобы вызвать единичную мутацию – наследственное изменение генетической программы, – требуется подвести каким-то способом к ДНК энергию в 2,5—3 электронвольта (эВ). Электронвольт – единица энергии: такую энергию приобретает электрон, ускоряемый напряжением в 1 вольт.
Много это или мало? Ведь ДНК в клетке находится в окружении молекул, движущихся с весьма высокой скоростью. Оказывается, что средняя энергия теплового движения молекул при тех температурах, когда жизнь возможна, составляет примерно 1/40 эВ. Иными словами, при физиологических температурах ДНК оказывается достаточно стабильной. Но проблема эта сложнее, чем кажется на первый взгляд.
Не следует забывать, что скорости молекул при хаотическом тепловом движении неодинаковы. Убедиться в этом нетрудно. В 1827 году шотландский ботаник Р. Броун, разглядывая в микроскоп каплю воды с пыльцой растений, обнаружил, что взвешенные в жидкости пыльцевые зерна микронного размера не остаются на месте, а хаотически движутся как бы непрерывно подталкиваемые беспорядочными ударами чего-то невидимого.
Достойно удивления, что Броуново движение не привлекало внимания физиков (может быть, потому, что открыл его ботаник?) до начала нашего века, до исследований Альберта Эйнштейна, польского физика Мариана Смолуховского и французского физика Жана Перрена. А ведь из него не только вытекала непреложность существования молекул, но и возможность оценить их скорости и размеры!
Что происходит с частицей при броуновском движении? Со всех сторон она подвергается ударам молекул. Если она имеет достаточно большие размеры, то удары со всех сторон оказываются скомпенсированными – частица остается на месте. Но если размер ее, допустим, 10-5см, то весьма вероятно, что с какой-либо стороны суммарный импульс будет больше, и частица сдвинется в непредсказуемую сторону.
Иначе и быть не может: ведь скорости молекул разные и флуктуации в их распределении неизбежны. Именно от этих флуктуаций зависит голубой цвет ясного неба, так как на них сильнее рассеиваются синие лучи. Будь скорости всех молекул одинаковыми, солнечный свет не рассеивался бы и солнце светило бы в черном небе, как в космическом пространстве.
Эти же флуктуации кладут предел усилению слабых сигналов в электрических цепях. В конце концов мы слышим лишь треск, результат теплового движения электронов в цепях усилителя. Чтобы уйти за этот предел, приходится охлаждать приемник жидким азотом, водородом, а то и гелием.
Отсюда однозначно следует, что в любой достаточно большой популяции молекул неизбежно найдутся такие, которые могут нарушить структуру гена и вызвать мутацию. Ясно, что такие изменения генетических программ должны обладать следующими свойствами:
1. Они случайны в том смысле, что вероятность каждого единичного изменения не равна единице. Более стабильные части гена мутируют с меньшей частотой, более лабильные – с большей, но мы можем говорить лишь о большей или меньшей вероятности мутаций.
2. Они непредсказуемы, поскольку для предсказания какой-либо мутации мы должны знать координаты и импульсы всех молекул данной клетке.
3. Они не направлены в том смысле, что изменяют генетичеекую программу без учета содержания сохраняющейся в ней информации. Поэтому они только случайно могут оказаться адаптивными, приспособительными.
Не одна температура изменяет содержание генетических программ.
Пожалуй, большее значение имеют кванты жесткого излучения начиная с ультрафиолета, быстро движущиеся элементарные частицы, молекулы веществ, способные реагировать с ДНК (химические мутагены).
Начнем с радиации. Растворы чистых нуклеиновых кислот в видимом свете прозрачны. Однако и ДНК и РНК сильно поглощают ультрафиолет. Максимум поглощения на 260 нанометрах, кванты такой энергии уже могут вызывать и вызывают мутации. Еще более эффективны рентгеновы лучи и гамма-излучение, а также быстрые частицы, возникающие при ядерных распадах. Действие жестких излучений на генетические программы изучает целая отрасль генетики радиогенетика. О ее достижениях в популярной литературе много писалось. Остановимся подробнее на другом вопросе. В случае, когда мутацию вызывает квант или элементарная частица, наиболее четко проступает принцип необходимости квантовой механики для описания мутагенеза.
Мы уже договорились с самого начала, что физические законы, физические аксиомы должны оправдываться и для тех случаев, когда мы имеем дело с живой материей.
Значит, и здесь должен действовать принцип квантовомеханической неопределенности, который впервые сформулировал В. Гейзенберг в 1927 году.
Выражается этот великий принцип на вид простой формулой
и утверждает, что нельзя определить одновременно координату и импульс (то есть энергию) элементарной частицы точнее, чем следует из этого неравенства.Δp•Δx≥h
Произведение погрешности в определении координаты (Δx) на погрешность в определении импульса (Δp) не может быть меньше постоянной Планка h, равной 6,626 • 10–34Дж•с. Это ничтожная величина, поэтому, живя в макромире, мы ее не замечаем. Будь она побольше, мы бы жили в мире без ясных очертаний предметов и без четких траекторий движения. Но ведь мутации происходят в микромире. Для всего гена неопределенность пренебрежимо мала. Однако чтобы вызвать мутацию, нужно довести определенное количество энергии до участка гена размером всего лишь 10-7см (так называемый радиус эффективного обмена, впервые измеренный Н. В. Тимофеевым-Ресовским и К. Циммером). Здесь уже принципом неопределенности пренебрегать нельзя. О том, что для описания мутагенеза необходимо использовать принцип Гейзенберга, писали еще Н. В. Тимофеев-Ресовский и Паскуаль Йордан в 40-х годах. В последнее время то же утверждает и Манфред Эйген. Короче, мы можем лишь с некоторой долей вероятия предполагать, обладает ли частица нужной для мутации энергией и попадет ли она в зону эффективного объема.
Не только принцип Гейзенберга – любое другое положение квантовой механики делает процесс мутагенеза вероятностным. Вот еще пара примеров. Кроме стабильных атомов углерода и фосфора в состав ДНК могут входить их радиоактивные изотопы. Вероятность этого возрастает с увеличением радиоактивного фона нашей планеты. Местоположение атомов C14 и P32 в нитях ДНК, разумеется, случайно. А радиоактивный распад – тоже случайный процесс: знать, какой из атомов в данный момент распадется и вызовет мутацию, нам не дано, и запрет этот принципиальный.
Наконец, есть мутации, не индуцированные каким-нибудь внешним по отношению к ДНК агентом, а так называемые спонтанные. Водородные связи, скрепляющие двойную спираль ДНК, обусловлены ионами водорода (протонами). Если при раскручивании спирали ДНК, которое обязательно происходит при ее репликации, оба протона перейдут к одному из оснований, возможны другие комбинации, кроме АТ и ГЦ. А неверное спаривание оснований – мутация. Переход протона от одного основания к другому называется туннельным, описывается уравнениями квантовой механики, и все, что мы можем сказать о нем, то, что он осуществляется с частотой 1011…1012в секунду. Время и место каждой единичной спонтанной мутации непредсказуемо принципиально.
А что же вещества, изменяющие структуру гена, – мутагены? Впервые в 1932—1934 годах советские ученые В. В. Сахаров и М. Е. Лобашов независимо друг от друга получили мутации у дрозофилы, действуя на мушек йодом и рядом других веществ. С того времени список мутагенов возрос тысячекратно, теперь каждое химическое соединение, которое предполагается внедрять в практику (лекарства, красители, пластмассы и т. д.), испытывается на мутагенный эффект. Открыты – И. А. Рапопортом и Ш. Ауэрбах – так называемые супермутагены, вызывающие мутации у 100 процентов обработанных ими особей.
Может показаться, что химические мутагены не имеют особого значения в природе, во всяком случае до появления химической промышленности. Простой пример свидетельствует, что это не так. Даже такое, с позволения сказать, вещество, как ион водорода, может индуцировать мутации.
Активная реакция среды измеряется, как вы знаете, в единицах pH. Это взятый с обратным знаком логарифм концентрации водородных ионов. Например, когда реакция нейтральная, pH равно семи и концентрация ионов водорода 10-7моля. ДНК довольно устойчива к подкислению или подщелачиванию, однако при pH ниже четырех начинается отщепление пуриновых оснований – гуанина и аденина.
Содержимое клетки в общем-то сильная буферная система, поддерживающая pH в районе семи (это не относится к специализированным органам вроде желудка, где высока концентрация соляной кислоты, или же слюнных желез некоторых улиток, которые могут выделять довольно концентрированную серную кислоту). Но вспомним о размерах клеток и обязательных флуктуациях молекул. Объем бактериальной клетки всего 2•10-12см3, значит, при pH = 7 в ней всего примерно 120 водородных ионов. А это означает, что очень вероятна флуктуация: ионы водорода соберутся около, например, гуанина и отщепят его от пентознофосфатного остова ДНК. Произойдет мутация, и опять же случайным, непредсказуемым образом.
На этом можно было бы и закончить обзор изменений генетических программ. Но хотелось бы рассказать о недавно открытых хромосомных перестройках, удивительных и ранее казавшихся невозможными.
Сначала в геномах бактерий, а потом и в геномах высших организмов обнаружили удивительные «кусочки» ДНК, которые могут перемещаться с места на место в хромосоме. Их образно назвали прыгающими, генами. Различают две категории таких последовательностей. Есть короткие (500—1500 пар нуклеотидов) и длинные (более двух тысяч). Они могут вычленяться из единственной хромосомы бактерии (этот процесс называется эксцизией, выстриганием) и встраиваться в нее в другом месте (инсерция). И не только в хромосому, прыгающие гены могут включаться в плазмиды – последовательности ДНК, существующие в бактериальной клетке независимо. Так же легко прыгающие гены могут встраиваться в геномы бактериофагов. Удивительного в этом ничего нет: существует вполне убедительное мнение о том, что плазмиды – «домашние», прирученные бактерией фаги.
Короткие инсерционные сегменты обладают многими удивительными свойствами. Почему-то оба конца их состоят из одинаковых последовательностей. Иногда же концевые участки перевервуты: на одном конце, например, АТГАГ, на другом – ГАГТА. Встраиваясь в ген, они часто блокируют процесс транкрипции – ген теряет активность. И наоборот, встраиваясь в перевернутом виде, они могут тот же ген активировать. Иными словами, они включают и выключают гены.
Еще более удивительны длинные прыгающие последовательности транспозоны. Именно они, встраиваясь в фаги, переносят от бактерии к бактерии гены устойчивости к антибиотикам. Так что фаги не только могут убивать бактерии или быть их «приживальщиками». Они переносят в мире микроорганизмов генетическую информацию. Так, насекомые не только вредят растениям, но и опыляют их. Ведь первые цветковые растения опылялись ветром. На их пыльниках кормилось много насекомых. Перепачканные пыльцой жуки и примитивные бабочки перелетали с пыльника на пыльник, и в результате возникли такие пары «растение – насекомое», которые не могут существовать друг без друга. Юкка, например, не может опыляться иначе как с помощью юкковой моли. И между фагом, лизирующим, «пожирающим» бактерию, фагом-«приживальщиком» и приносящей пользу плазмидой, которая переносит транспозоны, есть все переходные ступени.
Прыгающие гены описаны и у высших, ядерных организмов. У дрожжевых грибков они, например, в зависимости от положения в хромосоме, определяют пол клетки. Обнаружили их также у кукурузы и дрозофилы. Вот на последнем объекте хотелось бы остановиться подробнее.
Генетиков давно занимали случаи, когда гены в популяциях плодовой мушки-дрозофилы вдруг теряли стабильность и частота мутаций возрастала во много раз. Оказалось, что нестабильность того или иного гена обусловливается встроившимся в него инсерционным сегментом. В некоторых случаях удалось проследить, как прыгающий ген в течение нескольких лет кочует от одного участка хромосомы к другому и от одной хромосомы к другой, каждый раз вызывая новую мутацию. Шведские ученые Г. Исинг и К. Рамель проследили до сорока прыжков одной такой инсерции. Результатом была или полная инактивация гена, приводящая к смерти (летальная мутация), или же мутации «розовые глаза», «грубые глаза» (с неправильным расположением фасеток) и целый ряд других.
Такие феномены наблюдались не только в экспериментальных, рассаженных по пробиркам популяциях дрозофилы, но и в природе. Известны так называемые «модные» мутации. Название несколько неожиданное, но удачное. Например, в 30-х годах частота мутаций, вызывающих желтую окраску тела и белую окраску глаз, возросла в сотни раз, и эта «мода» длилась до 40-х годов. В 1968 году у дрозофил возникла «мода» на мутацию abnormal abdomen (брюшной отдел уродливой формы), которая продолжалась пять лет и была вытеснена лишь мутацией «опаленные щетинки».
По-видимому, причина «модных» мутаций – вирусы. Известно, что вирусы могут встраиваться в хромосомы не только бактерий, но и ядерных организмов (это доказано для аденовирусов, вируса герпеса и опухолеродных перерождающих нормальную клетку в злокачественную). Инсерция вируса в геном вызывает мутацию того гена, в который он встроился.
Примечательно, что есть явное сходство между интронами, о которых говорилось в прошлой главе, и инсерционными сегментами. Те и другие могут разделять смысловую часть гена, не находят отражения в аминокислотной последовательности кодируемого геном белка и, по-видимому, могут включать и выключать гены.
А что значит: включать и выключать?
По-видимому, это выглядит так: регуляторные элементы клетки могут узнавать и не узнавать встроенный в ген интрон (инсерционный сегмент). В первом случае идет транскрипция предшественника мРНК, во втором – нет. В конце концов, это находит отражение в изменениях признаков фенотипа. И если интрон перепрыгнет в другой ген, то он будет активировать его, а не тот, из которого он ушел.
Третья аксиома. Теперь пора подытожить все, что мы знаем о наследственных изменениях генетических программ и сформулировать аксиому биологии № 3.
Прежде всего, эти изменения случайны и ненаправленны. Их можно сравнить с шум в канале информации от родителей к потомкам. Если мутация полностью исказит смысл передаваемой по цепи поколений инструкции, она будет смертельной (летальной). Это бывает, когда в результате мутации блокируется синтез жизненно важного фермента.
Наоборот, часты случаи, когда мутация не сказывается на признаках фенотипа. Вспомним, что генетический код вырожден и одна и та же аминокислота кодируется несколькими кодонами. Если в результате мутации один кодон заменится другим, но синонимичным, в полипептидную цепь включится та же аминокислота и изменения фенотипа мы не обнаружим.
Между этими двумя полюсами лежит огромная область мутаций, так или иначе изменяющих фенотип. В каких-то конкретных условиях они могут оказаться полезными, носители их с большей вероятностью оставят потомство и передадут их потомству.
Это и есть дарвиновская неопределенная изменчивость – исходный материал для эволюции.
Аксиому биологии №3 мы можем сформулировать так:
В процессе передачи из поколения в поколение генетические программы в результате многих причин изменяются случайно и ненаправленно, и лишь случайно эти изменения оказываются приспособительными.
Третья аксиома вытекает из важнейших физических постулатов – из практической невозможности знать координаты и импульсы всех молекул в клетке, из чего следует распределение энергий молекул по Максвеллу, и из принципиальной невозможности достаточно точно определить координаты и импульсы частиц, атакующих гены (принцип Гейзенберга).
А так как и перераспределение энергий в совокупности молекул, и взаимодействие молекул в химических реакциях в конце концов можно свести к квантовомеханическим процессам, то и получается, что аксиома №3 – следствие квантовой механики.
Далеко не всем это нравится. М. Эйген в книге, на которую я уже ссылался, по этому поводу вспоминает Эйнштейна. В письме от 4 декабря 1926 года Эйнштейн писал Максу Борну: «Квантовая механика внушает большое почтение. Но внутренний голос говорит мне, что это не истинный Иаков. Теория дает много, но едва ли она подводит нас ближе к тайне Старика. Во всяком случае, я убежден, что он не играет в кости…»
Старик – это бог, Не следует считать Эйнштейна религиозным. Поклонник Спинозы, он называл богом реально существующую природу.
С тех пор прошло более 50 лет: Квантовая механика многократно показала свою непротиворечивость природе, и вряд ли даже Эйнштейн сейчас выступал бы против нее. Однако в биологии до сих пор есть авторы, не признающие ненаправленной случайной изменчивости по Дарвину.
В конечном счете они склоняются к точке зрения, высказанной за полвека до Дарвина в 1809 году великим французским натуралистом Ламарком. Согласно Ламарку, организмы в процессе эволюции изменяются, но изменения эти массовые, направленные и приспособительные. Со времени Ламарка их называли по-разному: наследованием благоприобретенных свойств, ассимиляцией условий внешней среды и т. д. и т. д.
В нашей литературе наиболее основательно эту точку зрения отстаивал крупный ихтиолог и географ Л. С. Берг. Он, правда, называл свои воззрения не ламаркизмом, а номогенезом (эволюцией на основе закономерностей, то есть закономерных, а не случайных изменений). Но хотя в своей книге, вышедшей в 1922 году, Берг решительно отрекся от ламаркизма, никого это не обмануло. Уже тогда оппоненты Берга поняли, что ламаркизм и номогенез различаются лишь терминологией.
Ибо, по Бергу, наследственные изменения захватывают массы особей, идут направленно и должны быть изначально целесообразными, приспособительными. Берг, как и Ламарк, вообще считал приспособительные реакции изначальным свойством живого.
Современные номогенетики как-то стыдливо обходят последний тезис Берга, предпочитая говорить лишь о направленности наследственных изменений, а не о целесообразности их. Но направленные мутации должны быть приспособительными – ведь все организмы на Земле к каким-то условиям приспособлены.
Мне доводилось слышать выступление физика, в котором выдвигалась такая идея. Мутации сами по себе вредны или нейтральны и обеспечить эволюцию материалом не могут. Движущая сила эволюции – приспособительные изменения, которые происходят крайне редко (например, раз в тысячу лет). Из-за их редкости мы их обнаружить не можем, поэтому все опыты, опровергающие приспособительное изменение генетических программ, оказываются бездоказательными.
Но физик должен был бы понять, что номогенез, или ламаркизм, – называйте, как хотите, – основан на нарушении принципа причинности. Следствие (приспособленность) в нем определяет причину (изменение генетической программы). И опять мы возвращаемся к Аристотелю с его конечной причиной и целью одновременно. Такие взгляды на эволюцию, называются телеологическими.
Для меня представляется совершенно непостижимым, что в наши дни находятся вполне серьезные ученые, не понимающие того, что было предельно ясно еще в прошлом веке Герцену. Вспомните его «Былое и думы»:
«Естествоиспытатели, хвастающие своим материализмом, толкуют о каких-то вперед задуманных планах природы, о ее целях и ловком избрании средств… Это – фатализм в третьей степени, в кубе; на первой кипит кровь Януария, на второй орошаются поля дождем по молитве, на третьей открываются тайные замыслы химического процесса, хвалятся экономические способности жизненной силы, заготовляющей желтки для зародышей и т. п.».Сказано как будто про наших современных номогенетиков. Я уже писал об этом подробнее в другой своей книге[7]. Здесь, пожалуй, будет довольно нескольких примеров того, как направленная приспособительная изменчивость вступает в противоречие с принципом причинности.
Допустим существование неких актов приспособления, чрезвычайно редких и потому ненаблюдаемых. Нет проще приема сделать их наблюдаемыми. Надо лишь увеличить число наблюдений. Пример из той же физики: нейтрино взаимодействует с веществом с чрезвычайно низкой вероятностью, эта элементарная частица может пронизать насквозь земной шар, и вероятность того, что она вступит в реакцию с каким-либо ядром, совершенно ничтожна. Но физики уверенно детектируют нейтрино и антинейтрино. Детектор здесь – большое количество атомов хлора (обычно в составе той жидкости, которая применяется в химчистке – тетрахлорэтилена). А источник искомой частицы – ядерный реактор, испускающий мощные потоки нейтрино.
Американские исследователи Джошуа и Эстер Ледерберги исследовали процесс приспособления бактерий к антибиотикам. Этот феномен – проклятие современной медицины. Прославленные пенициллин, стрептомицин, олеандомицин быстро теряют свою эффективность Нужно сказать, что помогают микробам в данном случае и врачи со слабой биологической подготовкой. Они прописывают больным недостаточные дозы (из боязни «как бы чего не вышло»), то есть фактически ведут в популяциях бактерий отбор на устойчивость к антибиотику. Или же, наоборот, впадают в другую крайность и закармливают пациента антибиотиками при пустяковой простуде, а в результате тот же антибиотик окажется недейственным при двустороннем воспалении легких. Вспомните о переносах генов плазмидами и бактериофагами! Ген, определяющий, например, устойчивость бактерии к стрептомицину, легко может быть передан даже бактериям другого вида. В последнее время возникла серьезная проблема с гноеродными золотистыми стафилококками – современный врач с набором антибиотиков оказался в положении хирурга прошлого века, бессильного перед послеоперационными осложнениями. Выход в одном – все время обновлять арсенал антибиотиков, по возможности обгоняя бактерий.