Мы уже рассматривали пример с вирусом табачной мозаики, который при подкислении среды распадается на отдельные молекулы белка и РНК. При подщелачивании происходит обратный процесс, именуемый самосборкой. Все вирусы в клетках хозяина возникают в результате самосборки молекул нуклеиновых кислот и белков, и их структуры однозначно определяются последовательностью аминокислот в белках (и, значит, нуклеотидов в ДНК).
И не только вирусы. В результате самосборки возникают все клеточные структуры – рибосомы и клеточные мембраны. А сами клетки? Возьмем для примера простейшее животное; всем известного пресноводного полипа – гидру. Фенотип ее состоит из немногих типов клеток (около десяти). Давно уже ставят эффектные опыты, когда гидр растирают на отдельные клетки и из них в результате процесса, похожего на самосборку, возникает целая гидра. Значит, структура фенотипа гидры однозначно определяется свойствами клеток, его слагающих.
С высшими организмами такой опыт не поставишь: слишком много типов клеток и слишком сложные структуры они образуют.
 Рис. 31. Схема получения аллофенных мышей. Из яйцеводов беременных мышей извлекают яйцеклетки начавшие дробиться. Лучше всего опыт удается после трех делений (стадия восьми бластомеров). Для наглядности яйцеклетки берутся у мышей разной масти. Если обработать делящееся яйцо проназой – ферментом, расщепляющим белки, оно распадается на отдельные бластомеры. Отмытые от фермента бластомеры снова слипаются, даже если они от разных пород мышей. «Реассоциированный» зародыш можно пересадить в яйцеводы другой самки мыши. Финал – приемная мать изумленно смотрит на ни на что не похожее потомство (как та Сова из «Винни-Пуха» которая по ошибке снесла гусиное яйцо).
Рис. 31. Схема получения аллофенных мышей. Из яйцеводов беременных мышей извлекают яйцеклетки начавшие дробиться. Лучше всего опыт удается после трех делений (стадия восьми бластомеров). Для наглядности яйцеклетки берутся у мышей разной масти. Если обработать делящееся яйцо проназой – ферментом, расщепляющим белки, оно распадается на отдельные бластомеры. Отмытые от фермента бластомеры снова слипаются, даже если они от разных пород мышей. «Реассоциированный» зародыш можно пересадить в яйцеводы другой самки мыши. Финал – приемная мать изумленно смотрит на ни на что не похожее потомство (как та Сова из «Винни-Пуха» которая по ошибке снесла гусиное яйцо).
Если растереть высшее животное, скажем кролика, в кашицу, из клеток он заново не восстановится, Но на ранних стадиях развития подобные эксперименты удавались. Вы знаете из школьного курса, что оплодотворенная яйцеклетка млекопитающего уже в яйцеводах начинает дробиться, образуя зародыш. После трех дроблений зародыш соответственно состоит из восьми клеток (бластомеров).
Зародышей мыши на этой стадии извлекали из яйцеводов и обрабатывали раствором проназы. Это фермент, расщепляющий белки. Дробящаяся яйцеклетка распадалась на отдельные бластомеры. Можно смешать бластомеры разных пород мышей, например различающихся по окраске, отмыть от проназы и увидеть, как они будут слипаться друг с другом, вновь образуя зародыш. Такой зародыш можно пересадить другой мыши и дорастить до рождения и взрослого состояния. Мышей, явившихся на свет в подобных опытах, называют аллофенными.
Они – потомки трех, четырех и более родителей (в зависимости от того, сколько зародышей мы смешали). На рис. 31 показано изумление приемной матери при виде своего мозаичного, полосатого потомства.
Постепенно в умах исследователей сформулировалась идея, что в геноме яйцеклетки закодирована лишь информация о первичной структуре белков и очередности и интенсивности их синтеза. Закодирован, короче говоря, не сам фенотип, а серия инструкций по его созданию – самосборке на уровне частей клетки, самосборке на уровне клеток, тканей и органов.
И тут оказалось, что ДНК в ядре… чересчур много. Всего мы ожидали, но только не этого! Проверим полученный ошеломляющий вывод простым расчетом. Молекулы разных белков различаются по величине и молекулярной массе, среди них есть и крошки, состоящие из немногих десятков аминокислот, и настоящие гиганты. Например, молекулярная масса молекулы синего дыхательного пигмента виноградной улитки гемоцианина около девяти миллионов дальтон. Но в расчетах удобнее оперировать средними величинами. «Средний» белок, характерный для всей живой природы, содержит примерно 300—350 аминокислотных остатков. Значит, размер «среднего» гена, кодирующего белок, без учета размера интронов, вставленных в него, около тысячи нуклеотидов.
Отсюда следует, что геном млекопитающих может содержать информацию о структуре миллиона и более белков, а геном плодовой мушки-дрозофилы – не менее ста тысяч белков. У некоторых земноводных с особо большим геномом – десятки миллионов! Эти фантастические величины совершенно не согласуются с данными, полученными другими методами. У той же дрозофилы гигантские хромосомы слюнных желез поперечно исчерчены, как бы поделены на доли – хромомеры. Генетические эксперименты как будто бы показывают, что в каждом хромомере содержится только один ген, кодирующий белок. А число хромомеров подсчитано – их около пяти тысяч. По-видимому, фенотип дрозофилы определяется синтезом лишь пяти тысяч белков. И это хорошо согласуется с данными, полученными методами биохимии. А ДНК в геноме в 20 раз больше.
Сколько белков могут синтезировать клетки млекопитающих, пока еще точно неизвестно. Но предварительные расчеты показывают: примерно 50 тысяч и уж никак не больше 100 тысяч. А ДНК хватает на 3—6 миллионов, опять получается почти стократный избыток. И если раньше исследователи ломали голову над тем, где еще может храниться генетическая информация, помимо ДНК, то теперь встает вопрос: а что же делает «лишняя» ДНК?
Намек на ответ можно найти в следующей аналогии. Мой коллега Владимир Васильевич Меншуткин, крупнейший наш специалист по моделированию биологических процессов, как-то решил смоделировать на быстродействующей ЭВМ процесс эволюции. В машинную память были вложены описания простейших организмов вроде членистого червя или же ланцетника. Подчиняясь простейшим закономерностям, вложенным в программу, они должны были эволюционировать вплоть до существ, уже похожих на предка человека – питекантропа, на худой конец, австралопитека. Но машина не приемлет человеческого многословия и описание вновь полученных форм выдает в виде краткой строчки условных символов с индексами. Ее приходилось расшифровывать, переводить в слова и полученного машиной зверя рисовать самому.
Меншуткин решил и этот последний этап – реконструкцию – возложить на машину: пусть она, например на экране дисплея, сама изобразит то животное, которое создала. Однако это оказалось неожиданно трудным. Когда мы рисуем машинное животное по выданному описанию, то не пририсовываем, например, хвост к голове, а передние ноги помещаем впереди задних, а не наоборот. К информации, выданной машиной, мы неявно прибавляем свою, из собственной головы. Но машина правильного расположения частей тела, увы, не знает. Ее приходится этому учить – закладывать в машинную память инструкцию по расположению и соотношению частей. Неожиданно получилось, что объем информации, заключенный в этих инструкциях, значительно превышает само описание! А ДНК учить некому, она должна воспроизводить сложный фенотип сама, без помощи более сложных информационных систем. Значит, помимо ДНК, потребной для кодирования белков, должна быть регуляторная, которая еще неизвестными нам путями «включает» и «выключает» нужные и ненужные данной клетке гены.
На рис. 25 дана схема становления сложного многоклеточного организма. Оплодотворенная клетка дробится на два бластомера, потом на четыре. На этих стадиях бластомеры идентичны. Недаром, случайно разъединившись, они развиваются самостоятельно и становятся идентичными копиями одного организма – однояйцевыми близнецами. Кстати, идентичными не совсем: Из одной половинки часто развивается правша, из другой – левша.
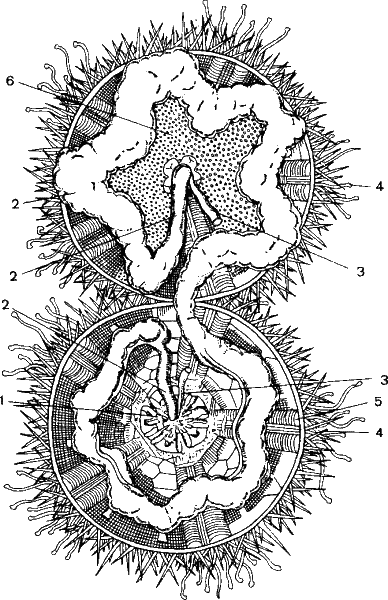
 Рис. 25. Индивидуальное развитие организма (онтогенез) как бы повторяет эволюционный процесс усложнения организации. Яйцеклетка, например, морского ежа дробится на бластомеры, первоначально одинаковые. Потом клетки зародыша изменяются, каждая по-своему, подчиняясь генетической программе. Из избыточной информации одинаковых бластомеров получается новая информация специализированных клеток, а затем тканей и органов. В результате образуется личинка – эхиноплутеус, а из нее – взрослый морской еж. Слева вверху тот же еж, в разрезе по экватору, чтобы было видно его сложное внутреннее строение – жевательный аппарат (Аристотелев фонарь – 1), кишечник (2), части воднососудистой системы (З, 4, 5), половые железы (6).
Рис. 25. Индивидуальное развитие организма (онтогенез) как бы повторяет эволюционный процесс усложнения организации. Яйцеклетка, например, морского ежа дробится на бластомеры, первоначально одинаковые. Потом клетки зародыша изменяются, каждая по-своему, подчиняясь генетической программе. Из избыточной информации одинаковых бластомеров получается новая информация специализированных клеток, а затем тканей и органов. В результате образуется личинка – эхиноплутеус, а из нее – взрослый морской еж. Слева вверху тот же еж, в разрезе по экватору, чтобы было видно его сложное внутреннее строение – жевательный аппарат (Аристотелев фонарь – 1), кишечник (2), части воднососудистой системы (З, 4, 5), половые железы (6).
Но уже после третьего дробления клетки зародыша становятся неидентичными – по размеру, набору синтезируемых белковых молекул и будущей судьбе в онтогенезе. В результате получается у млекопитающих около сотни различных типов клеток. И фактически все клетки, различные по форме и функциям с генетической точки зрения, отличаются только тем, что в них синтезируются разные белки, то есть транскрипция РНК идет с разных генов.
 Рис. 26. Из предыдущего рисунка вытекает любопытное следствие. Нетрудно сообразить, что чем раньше генетическая программа изменит свойства бластомера, тем резче скажется это на зародыше. Ведь бластомер потом делится десятки, а то и сотни раз, и все ткани и органы, получившиеся из него, также наследуют в десятки и сотни раз размноженное, усиленное изменение. Поэтому ранние изменения бластомеров невыгодны: организм может сойти с нормального пути развития. Изменения на более поздних стадиях не так сильно изменяют развитие. Так, при запуске космической ракеты ошибка в траектории на одну угловую секунду уведет ее за сотни тысяч километров от цели, но такая же ошибка в конце полета уже несущественна, ею можно пренебречь. Поэтому ранние стадии развития организмов должны быть консервативными, у самых далеких видов весьма схожими. Вот мы и пришли путем чисто логических рассуждений к одной из теорем биологии – так называемому биогенетическому закону Мюллера – Геккеля. Ранние стадии развития ехидны и кенгуру, оленя и кошки, мартышки и человека весьма сходны (верхний ряд). А как различаются взрослые организмы!..
Рис. 26. Из предыдущего рисунка вытекает любопытное следствие. Нетрудно сообразить, что чем раньше генетическая программа изменит свойства бластомера, тем резче скажется это на зародыше. Ведь бластомер потом делится десятки, а то и сотни раз, и все ткани и органы, получившиеся из него, также наследуют в десятки и сотни раз размноженное, усиленное изменение. Поэтому ранние изменения бластомеров невыгодны: организм может сойти с нормального пути развития. Изменения на более поздних стадиях не так сильно изменяют развитие. Так, при запуске космической ракеты ошибка в траектории на одну угловую секунду уведет ее за сотни тысяч километров от цели, но такая же ошибка в конце полета уже несущественна, ею можно пренебречь. Поэтому ранние стадии развития организмов должны быть консервативными, у самых далеких видов весьма схожими. Вот мы и пришли путем чисто логических рассуждений к одной из теорем биологии – так называемому биогенетическому закону Мюллера – Геккеля. Ранние стадии развития ехидны и кенгуру, оленя и кошки, мартышки и человека весьма сходны (верхний ряд). А как различаются взрослые организмы!..
Ведь геномы у них, как правило, остаются одинаковыми. Из этого правила, конечно, есть исключения. Как уже упоминалось, эритроциты млекопитающих вообще теряют ядра. У организмов со строго фиксированным числом клеток фенотипа часть материала хромосом выбрасывается из ядра: та ДНК, которая не нужна для жизнедеятельности специализированным клеткам. Особенно это наглядно у аскариды: ее хромосомы в процессе развития как бы обтрепываются, теряя значительные куски, но клетки, из которых развиваются половые, сохраняют геном в целости.
Но это исключение. А в подавляющем большинстве случаев, как бы ни отличалась клетка от исходной яйцеклетки, геном ее стабилен. Можно предположить, что, если мы активируем спящие, заблокированные гены ядра, мы заставляем клетку развиваться по несвойственному ей пути.
 Рис. 32. Профессор Кембриджского университета Дж. Гёрдон проделал замечательный опыт. Он облучил ультрафиолетом икринки шпорцевой лягушки (это земноводное, строго говоря, не лягушка, она близка к знаменитой жабе-пипе, хорошо разводится в лабораториях и стала излюбленным объектом эмбриологов). Тем самым ядра икринок были инактивированы, попросту убиты. Затем в каждую икринку микроманипулятором ввели ядра клеток кишечного эпителия другой лягушки. Для наглядности икринки брались у лягушки дикой буро-зеленой окраски, а ядра – от бело-розового альбиноса. Икринки после пересадки ядер развивались нормально, и все полученные таким способом лягушата оказались альбиносами. На рисунке вверху, слева направо: отец и мать лягушки, у которой брались ядра, и буро-зеленый донор икринок. Внизу: потомство (уместно задать вопрос: чье оно, кто их отец и мать?)
Рис. 32. Профессор Кембриджского университета Дж. Гёрдон проделал замечательный опыт. Он облучил ультрафиолетом икринки шпорцевой лягушки (это земноводное, строго говоря, не лягушка, она близка к знаменитой жабе-пипе, хорошо разводится в лабораториях и стала излюбленным объектом эмбриологов). Тем самым ядра икринок были инактивированы, попросту убиты. Затем в каждую икринку микроманипулятором ввели ядра клеток кишечного эпителия другой лягушки. Для наглядности икринки брались у лягушки дикой буро-зеленой окраски, а ядра – от бело-розового альбиноса. Икринки после пересадки ядер развивались нормально, и все полученные таким способом лягушата оказались альбиносами. На рисунке вверху, слева направо: отец и мать лягушки, у которой брались ядра, и буро-зеленый донор икринок. Внизу: потомство (уместно задать вопрос: чье оно, кто их отец и мать?)
В этом нас убеждают многочисленные опыты. Вот один из них, быть может, самый красивый. Английский ученый Дж. Гёрдон при помощи микроманипулятора – прибора, позволяющего разрезать бактерию на две равные половинки, вынул из клетки кишечного эпителия шпорцевой лягушки ядро и ввел его в яйцеклетку другой лягушки (ядро яйцеклетки было убито ультрафиолетовым излучением). Чужое ядро прижилось, яйцеклетка стала дробиться, и получился нормальный головастик. Лягушка, получившаяся из него, обладала всеми признаками той особи, из которой бралось ядро. Кстати, любопытный вопрос: кто мать и кто отец полученной в эксперименте лягушки? Для наглядности ядро извлекалось из клетки бело-розовой лягушки, альбиноса, и пересаживалось в яйцеклетку обычной буро-зеленой. Все полученные таким способом особи оказались альбиносами.
Ясно, что донор яйцеклетки не может быть матерью. Яйцеклетка здесь – своеобразный инкубатор. И отец, и мать этого фантастического лягушонка – родители той особи, от которой было позаимствовано ядро. Не исключена возможность, что проблема клонирования, подобной репликации всех животных и людей будет решена раньше, чем человечество перейдет рубеж двухтысячного года.
Итак, проблема «лишней» ДНК как будто бы разрешается. Это в основной своей части регуляторная ДНК, диктующая клетке, кем ей быть. Дифференцированные клетки, в свою очередь, взаимодействуя друг с другом, как аминокислоты в полипептидной цепочке, образуют новые структуры. Только связи здесь другие: это главным образом «силы Ван-дер-Ваальса» – те самые, которые участвуют в построении мембран клеток.
Новые структуры – ткани организма. И из них возникают органы и весь организм в целом. Матричный синтез и самосборка – вот что определяет в конечном счете построение фенотипа.
Но этого мало. Фенотип гораздо беднее генотипа, трудами которого он построен. Мы храним в своих генотипах информацию о строении многих фенотипов своих предков – и непосредственных, и гораздо более далеких.
Ребенок часто бывает похож не на отца или мать, а на дедушку или бабушку. Гены, определявшие голубой цвет глаз дедушки, проявились в фенотипе внука, хотя отец и был кареглазым. Такие случаи весьма обычны, но они всегда поражают наше воображение, вселяя какой-то страх перед непонятной силой наследственности (вспомните хотя бы «Собаку Баскервилей», где не только фенотипические черты, но и черты характера пращура Гуго Баскервиля проявляются у Стэплтона, его отдаленного потомка).
Но память генотипа на много порядков выше памяти самых именитых дворянских родословных. Все мы знаем, что киты лишены задних конечностей. Потеряли они ноги, очевидно, еще в начале эоцена. Тем не менее в среднем каждый десятитысячный кашалот рождается с зачаточными задними ногами. А это может означать одно: генный набор, ответственный за появление в фенотипе задних конечностей, сохраняется в заблокированном виде 60 миллионов лет.
Все мы знаем, что волосяной покров млекопитающих развился из роговых чешуек древних рептилий. Аналогичными чешуйками покрыты хвосты мышей и крыс. Древний признак возродился спустя сотню миллионов лет у представителей другого класса!
Итак, в генотипах скрыта информация о структурах весьма древних предков. Но этого мало. Вспомним о способности организмов к регуляции. Например, бактерия кишечная палочка хорошо растет на стандартных средах где источником углерода является глюкоза. Заменим в среде глюкозу другим углеводом – молочным сахаром лактозой. Бактерии немедленно ответят на это воздействие: начнут синтезировать новый фермент, галактозидазу, расщепляющий лактозу на простые сахара (глюкозу и галактозу). Не добавь мы в среду молочный сахар, мы бы и не подозревали, что в геноме кишечной палочки существует ген, управляющий синтезом этого фермента, – ведь в фенотипе обычно он не выражается.
Таких ферментов, адаптивных или индуцируемых, в геномах бактерий закодировано немало, и они помогают им выжить в тех случаях, когда условия среды резко изменяются. Лишите бактерию аниона фосфорной кислоты, без которого не построишь ни ДНК, ни РНК, и она сразу включает ген, кодирующий щелочную фосфатазу. Это фермент, расщепляющий эфиры фосфорной кислоты. Правда, бактерию можно обмануть. Есть вещества, которые индуцируют синтез фосфатазы, но данным ферментом они не расщепляются, ген активируется вхолостую. И наоборот, некоторые соединения могли бы стать источником желанного аниона, но индуцировать синтез этого фермента они не могут.
 Рис. 33. При изменении внешних условий в клетках организмов могут «включаться» гены прежде заблокированные. Результат – изменение внешнего облика организма.
Рис. 33. При изменении внешних условий в клетках организмов могут «включаться» гены прежде заблокированные. Результат – изменение внешнего облика организма.
А – водяной лютик (подводные листья глубоко рассеченные, надводные цельные).
Б – водное растение стрелолист: подводные листья лентовидные, вытянутые, плавающие, почковидные, надводные действительно похожи на наконечники стрел. Индуктор изменения, здесь не вода, а слабая освещенность. Затените стрелолист, растущий на суше, и у него будут возникать лентовидные подводные листья.
В – сосна растущая в окружении себе подобных (1) и отдельно растущая (2) Г – простой опыт, показывающий, что у картофеля из одних и тех же зачатков на свету возникают листья, а в темноте – клубни (на этом основан нехитрый агротехнический прием – окучивание повышающее выход клубней)
Значит, информация о многих признаках фенотипа у бактерий в генотипе запасена впрок, обычно она не проявляется. А как дело обстоит у высших организмов? По-видимому, больше всего признаков фенотипа, запасенных впрок, у растений. Это и понятно: при резкой смене условий внешней среды животное может уйти в места, более благоприятные, а растения должны всю жизнь проводить в тех местах, куда попали их споры и семена.
Сравните, например, стройную сосну в сосновом бору, кряжистую отдельно растущую и чахлую искривленную сосенку на сфагновом верховом болоте. Здесь индуктор, изменяющий облик растения, – целый комплекс факторов внешней среды. Но можно подобрать примеры, где индуктор, казалось бы, легко выявить.
Хорошо известное многим водяное растение стрелолист имеет листья разной формы. Торчащие из воды отличаются листовой пластинкой, действительно похожей на наконечник стрелы, а погруженные в воду – лентовидные. Казалась бы, индуктор признака «лентовидные листья» – вода. На самом деле – уровень освещенности. Вырастите стрелолист без воды, сильно притеняя его, и у него появятся лентовидные листья.
А вот такой опыт может проделать любой. У картофеля клубни развиваются из тех же исходных структур, что и листья. Затените нижние листья на кусте непрозрачным экраном – и они приобретут форму клубней. И здесь индуктор – отсутствие света. Растение, как и бактерию, можно обмануть, заставив признак возникнуть вхолостую.
В мире животных подобные примеры описаны в первую очередь у видов сидячих и малоподвижных. Коралловые полипы, растущие в тихих водах лагуны, имеют совсем другую форму, чем те, которые выносят прибой на наружной отмели атолла. А ведь те и другие могут относиться к одному виду! В одной Франции были описаны многие десятки «видов» всем известных пресноводных ракушек-беззубок пока систематики не сообразили, что беззубка изменяет свой фенотип «с легкостью необычайной», чуть только изменится скорость течения и характер грунта в реке.
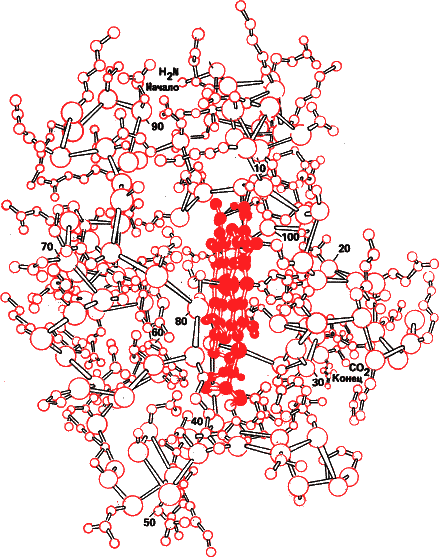 Рис. 30. Модель третичной структуры цитохрома С. Красным в центре молекулы показан активный центр – комплекс железа с органическим соединением порфирином. Ион железа, переходя из двух в трехвалентное состояние и наоборот, может отдавать и получать электрон. Малейшее изменение третичной структуры изменяет активность цитохрома. И вся эта причудливая форма определяется последовательностью аминокислот – первичной структурой кодируемой геном.
Рис. 30. Модель третичной структуры цитохрома С. Красным в центре молекулы показан активный центр – комплекс железа с органическим соединением порфирином. Ион железа, переходя из двух в трехвалентное состояние и наоборот, может отдавать и получать электрон. Малейшее изменение третичной структуры изменяет активность цитохрома. И вся эта причудливая форма определяется последовательностью аминокислот – первичной структурой кодируемой геном.
А активные, подыскивающие оптимальные условия организмы? Принцип регуляции признаков распространяется и на них. И самый простой пример – загар, меланиновый экран, не пропускающий избыточный ультрафиолет к клеткам нашего тела, который возникает у нас после продолжительного облучения солнцем или ртутной лампой.
Думаю, пора подытожить наши рассуждения. Мне всегда нравилось сравнивать фенотип с надводной частью айсберга, которая составляет одну десятую долю его подводной части – той структуры генотипа, которая не получила в фенотипе проявления. Подтаявший айсберг с грохотом переворачивается, обнажая источенную водой, необычной формы подводную часть. Организм, попадая в новые условия, может выявить новые признаки которых у него в фенотипе не было, но информация, кодирующая их структуру, и инструкции по их изготовлению находились в генотипе. Все имеющиеся по сей день в арсенале биологии факты однозначно свидетельствуют: вся информация, описывающая сложность фенотипа, содержится в генотипе (это, разумеется, не относится к таким признакам фенотипа, как синяк под глазом или ампутированная нога, но ведь эти признаки не наследуются).
Вторая аксиома биологии постулирует широкое распространение матричного копирования в жизненных процессах: новая ДНК копируется на матрице старой, мРНК копируется на матрице ДНК и, наконец, на матрице мРНК, с переходом на другой код копируется полипептидная цепь, образующая белок. Вся жизнь – это матричное копирование с последующей самосборкой копий.
Но этого мало. До сих пор мы говорили о статике жизни. Будь процесс матричного копирования на сто процентов идеально совершенным, невозможно было бы появление на свет автора этих записок, ни издателей их, ни читателей. Все мы находились бы на стадии протоклеток, плавающих в теплых водах морей докембрийской эпохи.
Пора нам рассмотреть динамику жизни. Она сводится к проблеме: подчиняясь каким закономерностям, изменяются наследственные матрицы?
Эта проблема и суть аксиомы биологии № 3.
РНК-матюраза – белок-самоубийца, он рубит сук, на котором сидит. Это фермент сплэйсинга, вырезающий интроны из первичного транскрипта, уничтожающий свою матрицу. После того как интроны удалены, начинается синтез цитохрома b. Но век фермента в клетке недолог. Когда концентрация матюразы упадет до определенного уровня, снова накапливается первичный транскрипт, из которого не удаляются интроны, и снова начинается синтез РНК-матюразы. Короче, получается гибкая система авторегуляции по принципу отрицательной обратной связи.
Не все еще ясно в этой схеме. Все ли интроны участвуют в подобной регуляции или же у них есть и другие функции? Насколько широко распространено это явление? Пока не знаем. Но лед уже тронулся: мало кто сейчас будет утверждать, что интроны в синтезе белка не участвуют.
Аксиома третья
И не только вирусы. В результате самосборки возникают все клеточные структуры – рибосомы и клеточные мембраны. А сами клетки? Возьмем для примера простейшее животное; всем известного пресноводного полипа – гидру. Фенотип ее состоит из немногих типов клеток (около десяти). Давно уже ставят эффектные опыты, когда гидр растирают на отдельные клетки и из них в результате процесса, похожего на самосборку, возникает целая гидра. Значит, структура фенотипа гидры однозначно определяется свойствами клеток, его слагающих.
С высшими организмами такой опыт не поставишь: слишком много типов клеток и слишком сложные структуры они образуют.
Если растереть высшее животное, скажем кролика, в кашицу, из клеток он заново не восстановится, Но на ранних стадиях развития подобные эксперименты удавались. Вы знаете из школьного курса, что оплодотворенная яйцеклетка млекопитающего уже в яйцеводах начинает дробиться, образуя зародыш. После трех дроблений зародыш соответственно состоит из восьми клеток (бластомеров).
Зародышей мыши на этой стадии извлекали из яйцеводов и обрабатывали раствором проназы. Это фермент, расщепляющий белки. Дробящаяся яйцеклетка распадалась на отдельные бластомеры. Можно смешать бластомеры разных пород мышей, например различающихся по окраске, отмыть от проназы и увидеть, как они будут слипаться друг с другом, вновь образуя зародыш. Такой зародыш можно пересадить другой мыши и дорастить до рождения и взрослого состояния. Мышей, явившихся на свет в подобных опытах, называют аллофенными.
Они – потомки трех, четырех и более родителей (в зависимости от того, сколько зародышей мы смешали). На рис. 31 показано изумление приемной матери при виде своего мозаичного, полосатого потомства.
Постепенно в умах исследователей сформулировалась идея, что в геноме яйцеклетки закодирована лишь информация о первичной структуре белков и очередности и интенсивности их синтеза. Закодирован, короче говоря, не сам фенотип, а серия инструкций по его созданию – самосборке на уровне частей клетки, самосборке на уровне клеток, тканей и органов.
И тут оказалось, что ДНК в ядре… чересчур много. Всего мы ожидали, но только не этого! Проверим полученный ошеломляющий вывод простым расчетом. Молекулы разных белков различаются по величине и молекулярной массе, среди них есть и крошки, состоящие из немногих десятков аминокислот, и настоящие гиганты. Например, молекулярная масса молекулы синего дыхательного пигмента виноградной улитки гемоцианина около девяти миллионов дальтон. Но в расчетах удобнее оперировать средними величинами. «Средний» белок, характерный для всей живой природы, содержит примерно 300—350 аминокислотных остатков. Значит, размер «среднего» гена, кодирующего белок, без учета размера интронов, вставленных в него, около тысячи нуклеотидов.
Отсюда следует, что геном млекопитающих может содержать информацию о структуре миллиона и более белков, а геном плодовой мушки-дрозофилы – не менее ста тысяч белков. У некоторых земноводных с особо большим геномом – десятки миллионов! Эти фантастические величины совершенно не согласуются с данными, полученными другими методами. У той же дрозофилы гигантские хромосомы слюнных желез поперечно исчерчены, как бы поделены на доли – хромомеры. Генетические эксперименты как будто бы показывают, что в каждом хромомере содержится только один ген, кодирующий белок. А число хромомеров подсчитано – их около пяти тысяч. По-видимому, фенотип дрозофилы определяется синтезом лишь пяти тысяч белков. И это хорошо согласуется с данными, полученными методами биохимии. А ДНК в геноме в 20 раз больше.
Сколько белков могут синтезировать клетки млекопитающих, пока еще точно неизвестно. Но предварительные расчеты показывают: примерно 50 тысяч и уж никак не больше 100 тысяч. А ДНК хватает на 3—6 миллионов, опять получается почти стократный избыток. И если раньше исследователи ломали голову над тем, где еще может храниться генетическая информация, помимо ДНК, то теперь встает вопрос: а что же делает «лишняя» ДНК?
Намек на ответ можно найти в следующей аналогии. Мой коллега Владимир Васильевич Меншуткин, крупнейший наш специалист по моделированию биологических процессов, как-то решил смоделировать на быстродействующей ЭВМ процесс эволюции. В машинную память были вложены описания простейших организмов вроде членистого червя или же ланцетника. Подчиняясь простейшим закономерностям, вложенным в программу, они должны были эволюционировать вплоть до существ, уже похожих на предка человека – питекантропа, на худой конец, австралопитека. Но машина не приемлет человеческого многословия и описание вновь полученных форм выдает в виде краткой строчки условных символов с индексами. Ее приходилось расшифровывать, переводить в слова и полученного машиной зверя рисовать самому.
Меншуткин решил и этот последний этап – реконструкцию – возложить на машину: пусть она, например на экране дисплея, сама изобразит то животное, которое создала. Однако это оказалось неожиданно трудным. Когда мы рисуем машинное животное по выданному описанию, то не пририсовываем, например, хвост к голове, а передние ноги помещаем впереди задних, а не наоборот. К информации, выданной машиной, мы неявно прибавляем свою, из собственной головы. Но машина правильного расположения частей тела, увы, не знает. Ее приходится этому учить – закладывать в машинную память инструкцию по расположению и соотношению частей. Неожиданно получилось, что объем информации, заключенный в этих инструкциях, значительно превышает само описание! А ДНК учить некому, она должна воспроизводить сложный фенотип сама, без помощи более сложных информационных систем. Значит, помимо ДНК, потребной для кодирования белков, должна быть регуляторная, которая еще неизвестными нам путями «включает» и «выключает» нужные и ненужные данной клетке гены.
На рис. 25 дана схема становления сложного многоклеточного организма. Оплодотворенная клетка дробится на два бластомера, потом на четыре. На этих стадиях бластомеры идентичны. Недаром, случайно разъединившись, они развиваются самостоятельно и становятся идентичными копиями одного организма – однояйцевыми близнецами. Кстати, идентичными не совсем: Из одной половинки часто развивается правша, из другой – левша.
Но уже после третьего дробления клетки зародыша становятся неидентичными – по размеру, набору синтезируемых белковых молекул и будущей судьбе в онтогенезе. В результате получается у млекопитающих около сотни различных типов клеток. И фактически все клетки, различные по форме и функциям с генетической точки зрения, отличаются только тем, что в них синтезируются разные белки, то есть транскрипция РНК идет с разных генов.
Ведь геномы у них, как правило, остаются одинаковыми. Из этого правила, конечно, есть исключения. Как уже упоминалось, эритроциты млекопитающих вообще теряют ядра. У организмов со строго фиксированным числом клеток фенотипа часть материала хромосом выбрасывается из ядра: та ДНК, которая не нужна для жизнедеятельности специализированным клеткам. Особенно это наглядно у аскариды: ее хромосомы в процессе развития как бы обтрепываются, теряя значительные куски, но клетки, из которых развиваются половые, сохраняют геном в целости.
Но это исключение. А в подавляющем большинстве случаев, как бы ни отличалась клетка от исходной яйцеклетки, геном ее стабилен. Можно предположить, что, если мы активируем спящие, заблокированные гены ядра, мы заставляем клетку развиваться по несвойственному ей пути.
В этом нас убеждают многочисленные опыты. Вот один из них, быть может, самый красивый. Английский ученый Дж. Гёрдон при помощи микроманипулятора – прибора, позволяющего разрезать бактерию на две равные половинки, вынул из клетки кишечного эпителия шпорцевой лягушки ядро и ввел его в яйцеклетку другой лягушки (ядро яйцеклетки было убито ультрафиолетовым излучением). Чужое ядро прижилось, яйцеклетка стала дробиться, и получился нормальный головастик. Лягушка, получившаяся из него, обладала всеми признаками той особи, из которой бралось ядро. Кстати, любопытный вопрос: кто мать и кто отец полученной в эксперименте лягушки? Для наглядности ядро извлекалось из клетки бело-розовой лягушки, альбиноса, и пересаживалось в яйцеклетку обычной буро-зеленой. Все полученные таким способом особи оказались альбиносами.
Ясно, что донор яйцеклетки не может быть матерью. Яйцеклетка здесь – своеобразный инкубатор. И отец, и мать этого фантастического лягушонка – родители той особи, от которой было позаимствовано ядро. Не исключена возможность, что проблема клонирования, подобной репликации всех животных и людей будет решена раньше, чем человечество перейдет рубеж двухтысячного года.
Итак, проблема «лишней» ДНК как будто бы разрешается. Это в основной своей части регуляторная ДНК, диктующая клетке, кем ей быть. Дифференцированные клетки, в свою очередь, взаимодействуя друг с другом, как аминокислоты в полипептидной цепочке, образуют новые структуры. Только связи здесь другие: это главным образом «силы Ван-дер-Ваальса» – те самые, которые участвуют в построении мембран клеток.
Новые структуры – ткани организма. И из них возникают органы и весь организм в целом. Матричный синтез и самосборка – вот что определяет в конечном счете построение фенотипа.
Но этого мало. Фенотип гораздо беднее генотипа, трудами которого он построен. Мы храним в своих генотипах информацию о строении многих фенотипов своих предков – и непосредственных, и гораздо более далеких.
Ребенок часто бывает похож не на отца или мать, а на дедушку или бабушку. Гены, определявшие голубой цвет глаз дедушки, проявились в фенотипе внука, хотя отец и был кареглазым. Такие случаи весьма обычны, но они всегда поражают наше воображение, вселяя какой-то страх перед непонятной силой наследственности (вспомните хотя бы «Собаку Баскервилей», где не только фенотипические черты, но и черты характера пращура Гуго Баскервиля проявляются у Стэплтона, его отдаленного потомка).
Но память генотипа на много порядков выше памяти самых именитых дворянских родословных. Все мы знаем, что киты лишены задних конечностей. Потеряли они ноги, очевидно, еще в начале эоцена. Тем не менее в среднем каждый десятитысячный кашалот рождается с зачаточными задними ногами. А это может означать одно: генный набор, ответственный за появление в фенотипе задних конечностей, сохраняется в заблокированном виде 60 миллионов лет.
Все мы знаем, что волосяной покров млекопитающих развился из роговых чешуек древних рептилий. Аналогичными чешуйками покрыты хвосты мышей и крыс. Древний признак возродился спустя сотню миллионов лет у представителей другого класса!
Итак, в генотипах скрыта информация о структурах весьма древних предков. Но этого мало. Вспомним о способности организмов к регуляции. Например, бактерия кишечная палочка хорошо растет на стандартных средах где источником углерода является глюкоза. Заменим в среде глюкозу другим углеводом – молочным сахаром лактозой. Бактерии немедленно ответят на это воздействие: начнут синтезировать новый фермент, галактозидазу, расщепляющий лактозу на простые сахара (глюкозу и галактозу). Не добавь мы в среду молочный сахар, мы бы и не подозревали, что в геноме кишечной палочки существует ген, управляющий синтезом этого фермента, – ведь в фенотипе обычно он не выражается.
Таких ферментов, адаптивных или индуцируемых, в геномах бактерий закодировано немало, и они помогают им выжить в тех случаях, когда условия среды резко изменяются. Лишите бактерию аниона фосфорной кислоты, без которого не построишь ни ДНК, ни РНК, и она сразу включает ген, кодирующий щелочную фосфатазу. Это фермент, расщепляющий эфиры фосфорной кислоты. Правда, бактерию можно обмануть. Есть вещества, которые индуцируют синтез фосфатазы, но данным ферментом они не расщепляются, ген активируется вхолостую. И наоборот, некоторые соединения могли бы стать источником желанного аниона, но индуцировать синтез этого фермента они не могут.
А – водяной лютик (подводные листья глубоко рассеченные, надводные цельные).
Б – водное растение стрелолист: подводные листья лентовидные, вытянутые, плавающие, почковидные, надводные действительно похожи на наконечники стрел. Индуктор изменения, здесь не вода, а слабая освещенность. Затените стрелолист, растущий на суше, и у него будут возникать лентовидные подводные листья.
В – сосна растущая в окружении себе подобных (1) и отдельно растущая (2) Г – простой опыт, показывающий, что у картофеля из одних и тех же зачатков на свету возникают листья, а в темноте – клубни (на этом основан нехитрый агротехнический прием – окучивание повышающее выход клубней)
Значит, информация о многих признаках фенотипа у бактерий в генотипе запасена впрок, обычно она не проявляется. А как дело обстоит у высших организмов? По-видимому, больше всего признаков фенотипа, запасенных впрок, у растений. Это и понятно: при резкой смене условий внешней среды животное может уйти в места, более благоприятные, а растения должны всю жизнь проводить в тех местах, куда попали их споры и семена.
Сравните, например, стройную сосну в сосновом бору, кряжистую отдельно растущую и чахлую искривленную сосенку на сфагновом верховом болоте. Здесь индуктор, изменяющий облик растения, – целый комплекс факторов внешней среды. Но можно подобрать примеры, где индуктор, казалось бы, легко выявить.
Хорошо известное многим водяное растение стрелолист имеет листья разной формы. Торчащие из воды отличаются листовой пластинкой, действительно похожей на наконечник стрелы, а погруженные в воду – лентовидные. Казалась бы, индуктор признака «лентовидные листья» – вода. На самом деле – уровень освещенности. Вырастите стрелолист без воды, сильно притеняя его, и у него появятся лентовидные листья.
А вот такой опыт может проделать любой. У картофеля клубни развиваются из тех же исходных структур, что и листья. Затените нижние листья на кусте непрозрачным экраном – и они приобретут форму клубней. И здесь индуктор – отсутствие света. Растение, как и бактерию, можно обмануть, заставив признак возникнуть вхолостую.
В мире животных подобные примеры описаны в первую очередь у видов сидячих и малоподвижных. Коралловые полипы, растущие в тихих водах лагуны, имеют совсем другую форму, чем те, которые выносят прибой на наружной отмели атолла. А ведь те и другие могут относиться к одному виду! В одной Франции были описаны многие десятки «видов» всем известных пресноводных ракушек-беззубок пока систематики не сообразили, что беззубка изменяет свой фенотип «с легкостью необычайной», чуть только изменится скорость течения и характер грунта в реке.
А активные, подыскивающие оптимальные условия организмы? Принцип регуляции признаков распространяется и на них. И самый простой пример – загар, меланиновый экран, не пропускающий избыточный ультрафиолет к клеткам нашего тела, который возникает у нас после продолжительного облучения солнцем или ртутной лампой.
Думаю, пора подытожить наши рассуждения. Мне всегда нравилось сравнивать фенотип с надводной частью айсберга, которая составляет одну десятую долю его подводной части – той структуры генотипа, которая не получила в фенотипе проявления. Подтаявший айсберг с грохотом переворачивается, обнажая источенную водой, необычной формы подводную часть. Организм, попадая в новые условия, может выявить новые признаки которых у него в фенотипе не было, но информация, кодирующая их структуру, и инструкции по их изготовлению находились в генотипе. Все имеющиеся по сей день в арсенале биологии факты однозначно свидетельствуют: вся информация, описывающая сложность фенотипа, содержится в генотипе (это, разумеется, не относится к таким признакам фенотипа, как синяк под глазом или ампутированная нога, но ведь эти признаки не наследуются).
Вторая аксиома биологии постулирует широкое распространение матричного копирования в жизненных процессах: новая ДНК копируется на матрице старой, мРНК копируется на матрице ДНК и, наконец, на матрице мРНК, с переходом на другой код копируется полипептидная цепь, образующая белок. Вся жизнь – это матричное копирование с последующей самосборкой копий.
Но этого мало. До сих пор мы говорили о статике жизни. Будь процесс матричного копирования на сто процентов идеально совершенным, невозможно было бы появление на свет автора этих записок, ни издателей их, ни читателей. Все мы находились бы на стадии протоклеток, плавающих в теплых водах морей докембрийской эпохи.
Пора нам рассмотреть динамику жизни. Она сводится к проблеме: подчиняясь каким закономерностям, изменяются наследственные матрицы?
Эта проблема и суть аксиомы биологии № 3.
* * *
Эта книга была уже сдана в печать, когда появились сообщения о разгадке функции интронов по крайней мере в одном гене – гене цитохрома b митохондрий дрожжей. В этом гене 6 экзонов и 5 интронов. Оказалось, что интроны все-таки кодируют аминокислотную последовательность. Белок «считывается» с первичного транскрипта РНК, из которого еще не вырезаны интроны. Французские ученые, описавшие его, назвали этот белок РНК-матюразой.РНК-матюраза – белок-самоубийца, он рубит сук, на котором сидит. Это фермент сплэйсинга, вырезающий интроны из первичного транскрипта, уничтожающий свою матрицу. После того как интроны удалены, начинается синтез цитохрома b. Но век фермента в клетке недолог. Когда концентрация матюразы упадет до определенного уровня, снова накапливается первичный транскрипт, из которого не удаляются интроны, и снова начинается синтез РНК-матюразы. Короче, получается гибкая система авторегуляции по принципу отрицательной обратной связи.
Не все еще ясно в этой схеме. Все ли интроны участвуют в подобной регуляции или же у них есть и другие функции? Насколько широко распространено это явление? Пока не знаем. Но лед уже тронулся: мало кто сейчас будет утверждать, что интроны в синтезе белка не участвуют.
Аксиома третья
Опечатки генетических программ. Редкая книга обходится без опечаток. В издательских кругах бытует характерный исторический анекдот. В 1888 году известному издателю А. С. Суворину удалось добиться у царской цензуры разрешения на издание радищевского «Путешествия из Петербурга в Москву» тиражом… в сто экземпляров. Издательская культура была у Суворина на большой высоте, а в этом исключительном случае он даже заключил пари, что издаст книгу без единой опечатки. Книга вышла – и на обложке стояло: «Сочинение А. И. Радищева» (напомню, что великого демократа звали Александром Николаевичем).
Для чего я рассказал эту историю? Мы уже убедились, что в основе жизни лежит матричное копирование, в принципе аналогичное тому же книгопечатанию. Ясно, что в каналах передачи информации от ДНК к признакам организма и от ДНК родителей к ДНК потомков должен существовать какой-то шум – те же опечатки, только на молекулярном уровне. Каналов без шума не бывает, иное дело, что шум может быть пренебрежимо малым.
Рассмотрим сначала шумы в канале ДНК – ДНК, приводящие к изменению генетических программ. В первую очередь речь у нас пойдет об упаковке генетического материала.
ДНК или РНК простейших вирусов может представлять лишь цепочку нуклеотидов, ничем не защищенную от внешних воздействий (например, от действия ферментов нуклеаз, расщепляющих нуклеиновые кислоты). Однако у сложных вирусов она заключена в белковый защитный чехол.
ДНК бактерий также единичная последовательность. Концы ее стыкуются, и образуется кольцо, похожее на тысячекратно перекрученную ленту Мёбиуса, хорошо известную любителям математики. Ясно что при репликации кольцо это должно разрываться, иначе дочерняя последовательность будет соединена с материнской, как звенья в цепи. К бактериальной ДНК могут присоединиться молекулы белков, но в общем-то она «голая».
Иное дело у высших организмов с оформленным клеточным ядром. Прежде всего генетическая программа у них – многотомное издание. Если генетическая программа бактерии закодирована в одной молекуле ДНК, одной двойной спирали, то в ядре высших организмов – эукариот – их может быть несколько: от двух у лошадиной аскариды до нескольких тысяч у некоторых одноклеточных организмов – радиолярий и ряда растений. Такие тома называют хромосомами. Считается, что каждая хромосома содержит одну молекулу ДНК, но крайней мере у животных. Однако есть сильные доводы в пользу того, что у многих высших растений в хромосоме может быть несколько десятков, а то и сотня идентичных копий. ДНК в хромосомах чрезвычайно хитроумно уложена в комплексе со специальными ядерными белками гистонами. Иначе нельзя упаковать в микронные объемы молекулы длиной во много десятков сантиметров.
Для чего я рассказал эту историю? Мы уже убедились, что в основе жизни лежит матричное копирование, в принципе аналогичное тому же книгопечатанию. Ясно, что в каналах передачи информации от ДНК к признакам организма и от ДНК родителей к ДНК потомков должен существовать какой-то шум – те же опечатки, только на молекулярном уровне. Каналов без шума не бывает, иное дело, что шум может быть пренебрежимо малым.
Рассмотрим сначала шумы в канале ДНК – ДНК, приводящие к изменению генетических программ. В первую очередь речь у нас пойдет об упаковке генетического материала.
ДНК или РНК простейших вирусов может представлять лишь цепочку нуклеотидов, ничем не защищенную от внешних воздействий (например, от действия ферментов нуклеаз, расщепляющих нуклеиновые кислоты). Однако у сложных вирусов она заключена в белковый защитный чехол.
ДНК бактерий также единичная последовательность. Концы ее стыкуются, и образуется кольцо, похожее на тысячекратно перекрученную ленту Мёбиуса, хорошо известную любителям математики. Ясно что при репликации кольцо это должно разрываться, иначе дочерняя последовательность будет соединена с материнской, как звенья в цепи. К бактериальной ДНК могут присоединиться молекулы белков, но в общем-то она «голая».
Иное дело у высших организмов с оформленным клеточным ядром. Прежде всего генетическая программа у них – многотомное издание. Если генетическая программа бактерии закодирована в одной молекуле ДНК, одной двойной спирали, то в ядре высших организмов – эукариот – их может быть несколько: от двух у лошадиной аскариды до нескольких тысяч у некоторых одноклеточных организмов – радиолярий и ряда растений. Такие тома называют хромосомами. Считается, что каждая хромосома содержит одну молекулу ДНК, но крайней мере у животных. Однако есть сильные доводы в пользу того, что у многих высших растений в хромосоме может быть несколько десятков, а то и сотня идентичных копий. ДНК в хромосомах чрезвычайно хитроумно уложена в комплексе со специальными ядерными белками гистонами. Иначе нельзя упаковать в микронные объемы молекулы длиной во много десятков сантиметров.